Как работает корковая зона: подробности
Не забывая о наших трех схемах (входные сигналы, поступающие вверх по иерархии; разветвляющиеся сигналы, спускающиеся вниз по иерархии; обратная связь с задержкой в таламусе), приступим к рассмотрению того, как зона коры головного мозга выполняет свои функции. Нам необходимо выяснить следующее.
Как зона мозга классифицирует входящие сигналы (вспомните сортировку цветных листов бумаги по ведрам).
Как она усваивает последовательности сигналов (вспомните последовательности интервалов в песне и восприятие лиц посредством комбинаций типа «глаз-нос-глаз»).
Как корковая зона формирует постоянные паттерны, или «имена» последовательностей.
Как она составляет специфические прогнозы (встреча поезда в определенное время или прогнозирование следующего звука мелодии).
Допустим, что колонки одной зоны – это те же ведра, которые мы использовали для воображаемой сортировки получаемых цветных листов бумаги. Каждая колонка помечена точно так же, как и ведро. К клеткам слоя 4 в каждой колонке протянуты волокна ввода из нескольких зон, расположенных ниже по иерархии. Всякий раз, получая правильную комбинацию входных сигналов, они выдают импульсный разряд. Возбуждаясь, клетка слоя 4 «голосует» за то, что полученные сигналы соответствуют своему ярлыку. Точно так же, как при сортировке листов бумаги, входные сигналы могут быть неоднозначными (например несколько колонок могут подходить для единственного входного сигнала). Мы хотим, чтобы зона коры нашего головного мозга сделала выбор: лист бумаги может считаться либо красным, либо оранжевым, но ни в коем случае не тем и другим сразу. Колонка, которой входной сигнал соответствует более всего, должна предотвратить активизацию остальных колонок.
Этим занимаются так называемые тормозные нейроны. Они предотвращают активизацию соседних нервных клеток, эффективно заботясь о том, чтобы был только один «победитель». Эти тормозные нейроны влияют исключительно на зону вокруг «своей» колонки. Другими словами, несмотря на наличие механизма подавления, существует возможность одновременной активизации нескольких колонок. (В мозге ничто не бывает представлено одним единственным нейроном или единственной колонкой.) Для простоты понимания можем представить, что зона выбирает одну и только одну колонку-«победителя», однако на будущее запомним, что на самом деле одновременно активизируются множество колонок. Процесс, используемый корой головного мозга для классификации входных сигналов, сложен и плохо изучен. Я не собираюсь заводить вас в дебри. Вместо этого я предлагаю принять предположение, что зона коры головного мозга отсортировала входной сигнал как активизацию в наборе колонок. Тогда мы сможем сосредоточиться на формировании последовательностей и присвоении им названий.
Каким образом зона коры головного мозга сохраняет последовательность отсортированных сигналов? Я уже излагал свои соображения по этому поводу и сейчас хотел бы углубиться в подробности. Представьте себе, что вы – колонка нервных клеток, а сигнал, поступивший из зоны низшего порядка, заставил активизироваться одну из ваших клеток слоя 4. Вслед за этим возбуждаются клетки в слоях 2 и 3, потом – в слое 5, а затем – в 6-м. Постепенно активизируется вся колонка нейронов. У каждой из клеток слоев 2, 3 и 5 есть тысячи синапсов в слое 1. Если какие-то из этих синапсов активны во время разряда нервных клеток слоев 2, 3 и 5, они усиливаются. Если такое происходит часто, то синапсы слоя 1 становятся достаточно сильными для того, чтобы вызвать вспышки клеток в слоях 2, 3 и 5 даже тогда, когда клетки слоя 4 молчат. Таким образом, некоторые части колонки могут активизироваться, даже если они не получили входного сигнала от более низкой по иерархии зоны коры головного мозга. Именно так клетки слоев 2, 3 и 5 учатся предвосхищать собственную активизацию на основе сигнала, поступающего из слоя 1. «До обучения» колонка может активизироваться исключительно через клетки слоя 4. «После обучения» колонка может частично активизироваться через память. Когда колонка активизируется через синапсы слоя 1, то входящие в нее нейроны предвидят, что произойдет активизация снизу, – это и есть прогноз. Если бы колонка могла разговаривать, она бы сказала: «Когда я активизировалась в прошлом, то как раз этот набор моих синапсов слоя 1 был активен. Как только я увижу такой набор синапсов, сразу же начну генерировать импульс в предвкушении сигнала».
Вспомните, что половина входных сигналов слоя 1 происходит из слоя 5 клеток соседних колонок и других зон коры головного мозга. Это информация о том, что случилось мгновение тому назад. Она говорит нам, какие колонки были активными до того, как активизировалась наша колонка. Она говорит нам, каким был последний интервал мелодии, что я видел последним, что я чувствовал последним, какую фонему устной речи я услышал последней и так далее. Если порядок возникновения указанных сигналов является последовательным во времени, то колонки усваивают последовательность. Они будут возбуждаться одна за другой в определенной последовательности.
Вторая половина сигналов поступает в слой 1 из клеток слоя 6 зон, расположенных выше в иерархии. Эта информация является более стабильной и представляет «имя» последовательности, с которой вы непосредственно сталкиваетесь в данный момент времени. К примеру, если колонки представить как музыкальные фразы, то название мелодии будет «именем» последовательности. Если фонемы будут колонками, то «именем» будет слово, которое вы слышите. Если колонками будут произносимые вами слова, «именем» будет речь, которую вы произносите. Следовательно, в слое 1 содержится и информация об «имени» последовательности, и о последнем сигнале в ней. Благодаря этому одна колонка может использоваться многими последовательностями без возникновения недоразумений. Колонки «учатся» активизироваться в нужном контексте и в правильном порядке.
Я должен подчеркнуть, что синапсы слоя 1 не являются единственными синапсами, участвующими в процессе обучения тому, когда должна активизироваться колонка. Как я уже упоминал ранее, клетки получают и отсылают сигналы ко многим соседним колонкам. Ведь более 90% всех синапсов – соединения с клетками за пределами колонки, и большинство из них расположены не в слое 1. Например, у клеток слоев 2, 3 и 5 есть тысячи синапсов в слое 1, но, кроме того, они имеют и тысячи синапсов в собственных слоях. Общий принцип состоит в том, что нейроны стремятся получать любую информацию, которая поможет им спрогнозировать возникновение импульса снизу. Как правило, активность соседних колонок отличается высоким уровнем корреляции, поэтому существует много связей между соседними колонками. Например, если в поле вашего зрения находится прямая линия, это активизирует колонки одну за другой. Но достаточно часто для прогнозирования активности колонки нужна более обширная информация. Именно тогда вступают в игру синапсы слоя 1. Нервным клеткам или колонкам нейронов не известно, что означает тот или иной синапс, но посредством синапсов они прогнозируют, когда нужно активизироваться.
А сейчас давайте рассмотрим, как зона коры головного мозга формирует имя выученной последовательности. Опять представьте себя зоной коры головного мозга. Активность ваших колонок нейронов меняется при каждом новом входном сигнале. Вы успешно усвоили последовательность активизации ваших колонок, т. е. некоторые клетки ваших колонок активизируются до того, как поступит входной сигнал из более низких зон. Какую информацию вы посылаете в более высокие зоны коры головного мозга? Как мы знаем, клетки слоев 2 и 3 посылают свои аксоны в следующую вверх по иерархии область. Активность этих клеток является входным сигналом для более высоких областей. Вот тут и возникает проблема. Чтобы иерархия работала слаженно, вам нужно транслировать постоянный сигнал на протяжении всей выученной последовательности. Вы должны транслировать «имя» последовательности, а не ее детали. До того как вы «выучите» последовательность, вы можете передавать отдельные ее элементы, но, после того как вы ее усвоили и можете прогнозировать, какие колонки будут активизироваться, вы должны передавать постоянный сигнал. Однако я еще не рассказывал, как это происходит. Итак, пока что вы передаете каждый переменный сигнал независимо то того, можете вы его спрогнозировать или нет. Когда активизируется вся колонка, клетки слоев 2 и 3 посылают новый сигнал вверх по иерархии. Коре головного мозга нужен какой-то способ, позволяющий поддерживать постоянный сигнал на протяжении всей выученной последовательности. Нам нужен способ отключать сигналы, исходящие из слоев 2 и 3, когда колонка прогнозирует свою активность, или же, наоборот, активизировать эти клетки, когда колонка не может спрогнозировать свою активность. Только так можно создать постоянный сигнал имени.
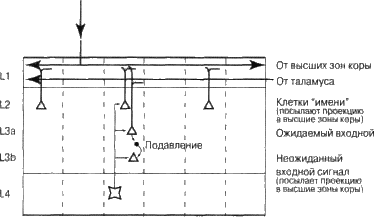
Рис. 6.10. Формирование постоянного «имени» для усвоенной последовательности
Науке очень мало известно о том, как именно кора головного мозга осуществляет подобную операцию. По моему мнению, существует несколько потенциально возможных методов. Я опишу тот, к которому больше всего склоняюсь в последнее время. (Не забывайте, что для нас сейчас наиболее важными являются концептуальные рамки, а не конкретный метод.) Ключевым моментом является создание постоянного сигнала «имени». Я хочу лишь показать, что существуют вполне правдоподобные механизмы для осуществления такого процесса.
Представьте, что вы – колонка (см. рис. 6.10). Постараемся разобраться, как вы учитесь посылать постоянный сигнал в вышестоящую в иерархии область, когда вам удается спрогнозировать свою активность, и переменный сигнал в противном случае. Сначала предположим, что в слои 2 и 3 входят несколько классов клеток. (Впридачу к нескольким типам тормозных нейронов многие анатомы различают несколько типов в слоях За и 3б, т. е. наше предположение вполне оправдано.)
Теперь предположим, что один класс клеток – клетки слоя 2 – учится быть активным на протяжении усвоенных последовательностей. Как группа этих клеток представляет «имя» последовательности? Она посылает постоянный сигнал в высшие зоны коры головного мозга до тех пор, пока их зона может предвидеть, какие колонки будут активизироваться дальше. Если ваша зона усвоила последовательность из трех различных сигналов, тогда клетки слоя 2 всех колонок, представляющих эти три сигнала, будут оставаться активными до тех пор, пока мы остаемся в пределах нашей последовательности. Они и являются «именем» последовательности.
Теперь предположим, что существует другой класс клеток – клетки слоя 3б, которые не активизируются, когда наша колонка успешно прогнозирует поступающие сигналы, но генерируют разряд в случае, если этого не происходит. Клетка слоя 3б олицетворяет' неожиданный сигнал. Она генерирует импульс при неожиданной активизации своей колонки. Она будет поступать так всякий раз, когда колонка, еще не «прошедшая» процесс обучения, активизировалась. По мере того как колонка учится прогнозировать свою активность, клетки слоя 36 «успокаиваются». Вместе взятые, клетки слоя 2 и слоя 3б удовлетворяют нашим требованиям. До того как было пройдено «обучение», и те и другие клетки затухают-вспыхивают в колонке, но по окончании «учебного курса» клетка слоя 2 остается активной постоянно, а клетка слоя 36 пребывает в состоянии покоя.
Как клетки учатся этому? Сначала давайте рассмотрим, как «отключить» клетки слоя 3б, когда колонка успешно прогнозирует свою активность. Допустим, существует клетка За, расположенная над клеткой 36, она имеет дендриты в слое 1. Ее единственная задача состоит в том, чтобы воспрепятствовать активизации клетки слоя 36, если она замечает в слое 1 уже известный паттерн. Когда клетка слоя За видит выученный сигнал в слое 1, она сразу же активизирует тормозной нейрон, который, в свою очередь, препятствует вспышке клетки слоя 36. Этого достаточно, чтобы удержать клетку слоя 3б от вспышки, когда колонка правильно прогнозирует свою активность.
А теперь рассмотрим более сложную задачу: как клетка слоя 2 остается активной на протяжении всей известной последовательности сигналов. Данная задача более сложна, потому что при этом различные клетки слоя 2 из многих колонок должны одновременно оставаться активными, даже если отдельные колонки в текущий момент таковыми не являются. По моему мнению, это может происходить следующим образом. Клетки слоя 2 учатся активизироваться исключительно высшими областями иерархии коры головного мозга. Они формируют синапсы преимущественно с аксонами из клеток слоя 6 областей, расположенных на уровень выше. Таким образом, клетки слоя 2 представляют постоянный сигнал «имени» из высшей области. Когда высшая зона посылает сигнал вниз к слою 1 расположенной ниже области, набор клеток слоя 2 нижней зоны активизируется, представляя все колонки, являющиеся членами последовательности. Клетки слоя 2 тоже имеют проекции в вышестоящей области и представляют собой частично стабильную группу клеток. (Вряд ли они постоянно остаются активными. Скорее всего, они вспыхивают синхронно, наподобие ритма.) Верхняя зона как бы посылает название мелодии в слой 1, расположенный ниже. Это заставляет вспыхивать набор клеток слоя 2, каждую клетку в своей колонке, и они будут активизироваться, когда вы слышите мелодию.
Все эти механизмы вместе взятые позволяют коре головного мозга усваивать последовательности, составлять прогнозы, формировать постоянные репрезентации («имена») для последовательностей входящих сигналов. Это основные операции, необходимые для формирования инвариантных репрезентаций.
Как мы формируем прогнозы о событиях, с которыми ранее не сталкивались? Как мы выбираем одну из десятков возможных интерпретаций входных сигналов? Как зона коры головного мозга формирует частные прогнозы на основе инвариантных схем? Мы уже рассматривали примеры прогнозирования следующего звука мелодии, когда в вашей памяти есть только интервал между отдельными нотами, историю со встречей поезда и воспроизведение Геттисбергской речи. Во всех перечисленных случаях единственным путем решения проблемы было использование последней полученной информации для того, чтобы конвертировать инвариантный прогноз в частное предсказание. Перефразировав ту же мысль в терминах коры головного мозга, можем сказать, что нужно сочетать поток прямой информации (поступающие фактические сигналы) с потоком обратной информации (прогноз в инвариантной форме).
Приведу простой пример того, как это, по моему представлению, происходит. Допустим, кора вашего головного мозга знает, что должна воспринять музыкальный интервал квинту. Колонки вашей зоны реагируют на отдельные интервалы, такие как «до-ми», «до-соль», «ре-ля» и так далее. Вам нужно решить, какие из колонок будут активными. Если зона выше говорит вам, что следует ожидать квинту, это активизирует клетки слоя 2 во всех колонках, представляющих квинты, то есть «до-соль», «ре-ля», «ми-си». Клетки слоя 2 из колонок, которые представляют другие интервалы, активизироваться не будут. Теперь вам нужно из колонок, представляющих квинты, выбрать только одну. Входными сигналами для вашей зоны будут отдельные ноты. Если «ре» была последней нотой, которую вы слышали, тогда все колонки, представляющие интервалы, в которых присутствует нота «ре», т. е. «ре-ми» и «ре-си», получают часть входного сигнала. Теперь в слое 2 активизировались все колонки, представляющие квинты, а в слое 4 мы получили часть сигнала во все колонки, представляющие интервалы, в которые входит нота «ре». Пересечение этих двух множеств и будет ответом на наш вопрос. Это будет колонка, представляющая интервал «ре-ля» (см. рис. 6.11).
Как кора головного мозга определяет пересечение? Мы раньше упоминали о том, что аксоны клеток слоев 2 и 3, выходя из коры головного мозга, формируют синапсы в слое 5. Аксоны, направляющиеся к слою 4 из более низких зон коры головного мозга, создают синапсы в слое 6. Пересечение этих двух синапсов (снизу вверх и сверху вниз) дает нам как раз то, что нужно. Клетки слоя 6, получающие эти два сигнала возбуждаются. Клетка слоя 6 «представляет мнение» зоны коры головного мозга о том, что происходит, т. е. специфический прогноз. Если бы клетка слоя 6 могла разговаривать, она бы сказала: «Я являюсь частью колонки, представляющей нечто. Конкретно в моем случае колонка представляет интервал „ре-ля“. У остальных колонок другие значения. Я представляю свою зону коры головного мозга. Когда я становлюсь активной, это означает, что, по нашему мнению, сейчас звучит интервал „ре-ля“, или же он прозвучит в ближайшем будущем. Возможно, я активизировалась, потому что сигнал снизу вверх, поступивший от ушей, заставил клетку слоя 4 моей колонки возбудить всю колонку. Или же моя активизация может означать, что мы узнали мелодию и прогнозируем следующий интервал. В любом случае моя задача состоит в том, чтобы довести до ведома низших зон коры головного мозга, что, по нашему мнению, происходит. Я представляю наше видение внешнего мира – независимо от того, является он настоящим или выдуманным».
Попробую описать указанный процесс с помощью визуальной картинки. Представьте себе два листа бумаги, усеянные множеством мелких дырочек. Дырочки на одном листе представляют колонки, у которых есть активные клетки слоев 2 и 3 (наш инвариантный прогноз). Дырочки на другом листе бумаги представляют колонки с частичным сигналом снизу. Если мы наложим листы бумаги друг на друга, то некоторые из дырочек совпадут, а некоторые – нет. Дырочки, которые совпали, представляют колонки, которые, как мы полагаем, должны активизироваться.
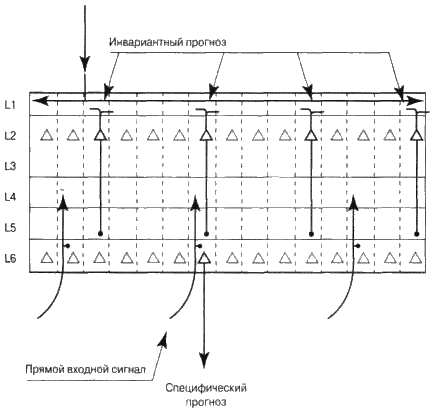
Рис. 6.11. Как зона коры головного мозга формирует специфические прогнозы, используя инвариантные репрезентации
Описанный механизм не только создает определенные прогнозы, он также разрешает неоднозначность входных сенсорных сигналов. Как мы помним из примеров с цветными листами бумаги и искаженным изображением, очень часто входные сигналы, полученные зоной коры головного мозга, являются неоднозначными. Механизм сочетания информации, поступающей снизу вверх и сверху вниз, помогает выбрать один из двух или более вариантов интерпретации. Как только вы приняли решение, каков будет выбор, вы сразу же транслируете этот выбор в область, расположенную ниже по иерархии.
Каждое мгновение нашей жизни каждая зона коры головного мозга сравнивает набор ожидаемых колонок, возбужденных сверху, с набором колонок, которые уже были возбуждены снизу. Там, где эти два набора пересекаются, создается наше восприятие. Если бы поступали совершенные сигналы снизу и у нас были бы совершенные прогнозы, тогда набор возбужденных колонок всегда совпадал бы с набором ожидаемых колонок. Но очень часто этого не происходит. Метод сочетания частичного прогноза и частичного входного сигнала позволяет определиться с неоднозначным входным сигналом, он позволяет заполнить пробелы в информации и остановить выбор на одном из альтернативных вариантов. Именно так мы комбинируем музыкальный интервал с последней услышанной нотой, чтобы спрогнозировать следующую ноту мелодии. Именно так мы определяемся, изображено на картинке два лица или ваза. Именно так мы разветвляем моторный поток на написание или произнесение вслух Геттисбергской речи.
И наконец, вдобавок к отправлению информации в более низкие зоны коры головного мозга, клетки слоя 6 могут посылать выходные сигналы назад, к клеткам слоя 4 той же колонки. Когда они это делают, наши прогнозы становятся входным сигналом. Именно этим мы занимаемся, размышляя или мечтая. Это позволяет нам видеть последовательности наших собственных прогнозов. Мы занимаемся этим, планируя свое будущее, вспоминая выступления или переживая о предстоящих событиях. Стефен Гросберг, длительное время занимавшийся моделированием деятельности коры головного мозга, называет это свернутой обратной связью, я же предпочитаю термин воображение.
И наконец, последний вопрос, который нам нужно затронуть, прежде чем перейти к следующему разделу. Я неоднократно подчеркивал, что чаще всего то, что мы видим, слышим, чувствуем, во многом зависит от наших собственных действий. То, что мы видим, зависит от того, на чем наши глаза остановились после очередной саккады, и от угла поворота головы. То, что мы ощущаем, зависит от того, как мы передвигаем пальцы и конечности. То, что мы слышим, иногда зависит от того, что мы делаем и говорим.
Значит, чтобы спрогнозировать, что мы почувствуем дальше, мы должны знать, что мы сейчас делаем. Моторное поведение и сенсорное восприятие тесно взаимосвязаны. Как нам удается составлять прогнозы, если то, что мы чувствуем, во многом является результатом наших собственных действий? К счастью, у данной проблемы существует неожиданное и простое решение, правда, многие его детали не очень понятны.
Неожиданное открытие: восприятие и поведение – это почти одно и то же. Как я уже упоминал раньше, большинство зон коры головного мозга (если не все) принимают участие в осуществлении движения.
Клеткам слоя 5, имеющим проекции в таламусе, а затем в слое 1, тоже свойственна моторная функция, поскольку у них одновременно есть проекции в моторных зонах «старого» мозга. Таким образом, знание о том, что только что произошло (как в сенсорном, так и в моторном плане), присутствует в слое 1.
Вторая неожиданность (являющаяся следствием первой): моторное поведение должно быть представлено в иерархии инвариантных репрезентаций. Вы генерируете движения, необходимые для осуществления определенного действия, не размышляя о нем во всех деталях. Когда моторная команда опускается вниз по иерархии, она трансформируется в сложные и подробные последовательности, необходимые для осуществления той деятельности, которая была запланирована. Это происходит как в сенсорной, так и в моторной зонах коры головного мозга, что размывает грань между ними. Если зона IT зрительной коры головного мозга воспринимает репрезентацию «нос», то простое действие переключения на репрезентацию «глаза» будет вызывать саккаду, необходимую для оправдания прогноза. Саккада, необходимая для того, чтобы перейти от зрительного восприятия носа к зрительному восприятию глаза, меняется в зависимости от расположения лица. Если лицо расположено поблизости, это требует большей саккады, и наоборот, более отдаленное лицо требует меньшей саккады. Наклоненное лицо требует саккады под углом, который отличается от используемого для лица без наклона. То, какой должна быть саккада, определяется, пока прогноз о том, что следующим мы увидим глаз, спускается к зоне V1. Чем дальше опустился прогноз, тем более точной становится саккада. Заканчивается все тем, что центральные ямки ваших глаз останавливается точно на объекте или же очень близко к нему.
Давайте рассмотрим еще один пример. Для того чтобы я физически переместился из гостиной на кухню, все, что нужно моему мозгу, – это мысленно переключиться с инвариантной репрезентации гостиной на инвариантную репрезентацию кухни. Данное переключение вызывает сложное разворачивание последовательностей. Процесс генерирования последовательности прогнозов того, что я увижу, услышу, почувствую, пока буду переходить из гостиной на кухню, тоже генерирует последовательность моторных команд, заставляющих меня перейти из гостиной на кухню и перемещать мой взгляд, пока я двигаюсь. Прогнозирование и моторное поведение идут рука об руку, в то время как потоки сигналов одновременно двигаются вверх и вниз по иерархии коры головного мозга. Каким бы странным это ни казалось, но, когда в дело вовлечено ваше поведение, ваши прогнозы не только предшествуют ощущению – они также его предопределяют. Прогнозирование перехода к следующему сигналу последовательности вызывает каскадное прогнозирование того, что вы должны пережить дальше. Когда каскадный прогноз разворачивается, он генерирует моторные команды, необходимые для осуществления прогноза. Воображение, прогнозирование и действие являются частями одного и того же разворачивания последовательностей в иерархии коры головного мозга.
Действие в воображении, параллельное разворачивание сенсорного и моторного поведения являются основой поведения, ориентированного на результат. Целенаправленное поведение, этот священный Грааль робототехники, изначально встроено в ткань коры головного мозга.
Конечно, мы можем отключить наше моторное поведение. Я могу представить себе, что вижу что-то, хотя на самом деле этого не вижу. Я могу представить себе, как иду на кухню, хотя на самом деле этого не делаю. Но действие всегда начинается в нашем воображении.
Назад: Строение зон коры головного мозга
Дальше: Поток вниз, поток вверх

