Книга: Об интеллекте
Назад: Последовательности последовательностей
Дальше: Как работает корковая зона: подробности
Строение зон коры головного мозга
А сейчас давайте рассмотрим отдельную зону коры головного мозга, одну из тех, что изображены блоками на рис. 6.5.
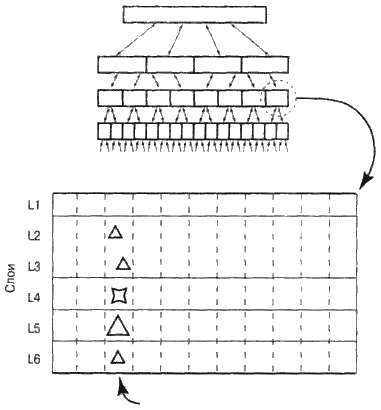
Рис. 6.6. Слои и колонки в зоне коры головного мозга
На рис. 6.6 приведено схематическое строение зоны коры головного мозга. В этом разделе я покажу, как клетки зоны коры головного мозга распознают и вызывают из памяти последовательности сигналов, обеспечивая тем самым формирование инвариантных репрезентаций и составление прогнозов. Начнем с описания структуры, общей для всех зон коры головного мозга. Прежде всего, между ними имеются значительные расхождения в размерах. Наиболее обширными являются первичные сенсорные зоны. К примеру, зона V1 занимает в задней части мозга область, приблизительно соответствующую площади паспорта. Как я уже упоминал выше, она состоит из множества подзон, которые могут быть не больше букв на этой странице. Но давайте пока предположим, что типичная зона коры головного мозга размером с мелкую монету.
Вспомните шесть визиток, с помощью которых в главе 3 мы представили слои ткани неокортекса. Что это за слои? Если мы рассмотрим корковую зону размером с монету под микроскопом, то увидим, что при передвижении снизу вверх плотность и форма нервных клеток различаются. Верхний слой 1 сильнее всего отличается от остальных слоев. Он состоит из густого переплетения аксонов, идущих параллельно поверхности коры, нейронов в этом слое почти нет. Строение слоев 2 и 3 сходно – они густо «населены» большим количеством пирамидных клеток. Слой 4 состоит из нервных клеток звездчатой формы. В слое 5 мы увидим как пирамидные клетки обычных размеров, так и чрезвычайно большие. Несколько типов нейронов есть и в нижнем слое 6.
Визуально мы воспринимаем горизонтальные слои, но большее значение имеют колонки клеток, располагающиеся перпендикулярно. Можете считать колонки вертикальными объединениями клеток, которые работают вместе. (Термин «колонка» вызывает много дебатов в научных кругах. Нейробиологи спорят об их размере, функциях и роли в целом. Для удобства будем мыслить в рамках их архитектуры, поскольку то, что у коры столбчатая архитектура, признается всеми.) Слои в пределах каждой колонки связываются аксонами, которые проходят вверх-вниз и создают синапсы. Отмечу, что наши колонки не имеют четких очертаний – в коре нет ничего простого. То, что колонки существуют, доказывается следующими положениями.
Положение 1. Вертикально расположенные клетки одной колонки активизируются под воздействием одних и тех же стимулов. Так, при подробном рассмотрении колонок зоны V1 мы обнаружим, что некоторые из них соответствуют линейным сегментам с правым уклоном (/), а другие – линейным сегментам с левым уклоном (\). Клетки из одной колонки тесно связаны, именно поэтому вся колонка реагирует на один и тот же стимул. Например, активная клетка слоя 4 активизирует расположенные над ней клетки слоев 3 и 2, что, в свою очередь, активизирует клетки, входящие в слои 5 и 6. Активность распространяется вниз и вверх в пределах колонки.
Положение 2. Наличие колонок нейронов в зонах неокортекса объясняется особенностями формирования коры головного мозга в эмбриогенезе. Эмбриональные клетки-предшественники мигрируют из внутренней мозговой впадины к месту образования коры головного мозга. Каждая из указанных клеток делится, образуя приблизительно сотню нейронов, называемых микроколонками. Эти нейроны связаны друг с другом вертикально так, как я описал. Термин «колонка» часто используется достаточно свободно для описания разных явлений. Под колонкой может подразумеваться общая вертикальная связность в коре или же вполне определенная группа клеток, происходящих от одной клетки-предшественницы. В рамках последнего определения мы можем утверждать, что в коре головного мозга есть несколько сотен миллионов микроколонок.
Чтобы вам легче было представить эту колончатую структуру, вообразите микроколонку в человеческий волос толщиной. Возьмите тысячи волосков и разрежьте их на небольшие сегменты, имеющие высоту строчной буквы Т. Если поставить их бок о бок очень плотно друг к другу и склеить, получится густая щетина. Теперь создайте покров из длинных и очень тонких волосков. Они будут представлять аксоны слоя 1. Приклейте их горизонтально к поверхности нашей щетины. Этот щетиноподобный коврик является упрощенной моделью зоны коры головного мозга размером с монету. Потоки информации преимущественно передвигаются преимущественно вдоль волосков: горизонтально в слое 1 и вертикально в слоях 2-6.
Рассмотрим еще одну важную особенность колонок, а затем перейдем к анализу их назначения. При пристальном рассмотрении мы увидим, что не менее 90% синапсов клеток из одной колонки – это связи с клетками, расположенными вне пределов самой колонки: некоторые соединения – с соседними колонками, некоторые – вообще с другими частями мозга. На основании чего же мы говорим о важности колонок, если большинство связей распределено по всему мозгу, а не локализовано?
Ответ на этот вопрос нам даст модель «Память-предсказание». В 1979 году Вернон Маунткастл не только заявил о существовании единого алгоритма коры головного мозга, но также предположил, что колонка нейронов в коре головного мозга является базовой единицей обработки информации. Правда, он не знал, какова именно функция колонки. Я считаю, что последняя является базовой единицей прогнозирования. Чтобы колонка нейронов могла прогнозировать, когда ей следует активизироваться, нервным клеткам необходимо знать, что происходит в других частях мозга. Именно в этом и заключается смысл обширных синаптических связей.
Ниже мы обсудим этот интересный момент подробнее, а сейчас я хочу дать вам общее представление о назначении подобных соединений нейронов в коре головного мозга. Чтобы предсказать следующий звук мелодии, вы учитываете ее название, ту часть, которая звучит в данный момент времени, время, прошедшее с момента последнего услышанного вами звука, и то, каким он был. Большое количество синапсов, связывающих нервные клетки одной колонки с остальными частями мозга, обеспечивает контекст, в котором и осуществляется прогностическая функция коры головного мозга.
Теперь обратимся к рассмотрению потоков информации, циркулирующих вверх-вниз по иерархии корковых зон. Восходящий поток сигналов проходит относительно прямой путь, который схематически отображен на рис. 6.7. Представьте, что перед нами зона коры головного мозга, состоящая из тысяч колонок. Увеличим одну из них. Входные сигналы из нижних зон попадают в слой 4, который является основным входящим слоем. По дороге они «отмечаются» в слое 6 (скоро вы поймете, почему это так важно). Клетки слоя 4 передают сигналы вверх – к клеткам слоев 2 и 3, находящимся в пределах той же колонки. Далее клетки слоев 2 и 3 передают информацию по аксонам еще выше – к входному слою следующей в иерархии зоны. Таким образом информация передвигается от зоны к зоне вверх по иерархии в пределах одной колонки нейронов.
Путь нисходящего потока информации (рис. 6.8) более сложен. Клетки слоя 6 посылают информацию вниз, в слой 1 области, расположенной ниже в иерархии. Аксоны нейронов слоя 1 распространяются на очень большие расстояния. Следовательно, информация, передвигающаяся вниз, потенциально может активизировать многие колонки нейронов. Хотя «своих» клеток в слое 1 очень мало, сигналы, проходящие через этот слой, могут активизировать клетки слоев 2, 3 и 5, поскольку у последних есть дендриты в слое 1. Аксоны, выходящие из клеток слоев 2 и 3. покидая кортекс, формируют синапсы в слое 5 и, как предполагается, возбуждают клетки слоев 5 и 6. Итак, мы приходим к выводу, что информация, передвигаясь вниз по иерархии, проходит более запутанный путь, чем в случае движения вверх. Благодаря слою 1 она может распространяться в самых разных направлениях. Обратная информация генерируется в клетках слоя б высшей зоны. Затем она распространяется в слое 1 низшей зоны. Клетки слоев 2, 3 и 5 низшей зоны возбуждаются, некоторые из них, в свою очередь, возбуждают клетки слоя 6, который проецируется в слой 1 более низких в иерархии областей и так далее. (При помощи рис. 6.8 понять описанные процессы намного проще.)
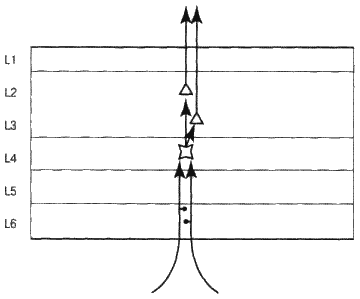
Рис. 6.7. Восходящий информационный поток
Приведу предварительное объяснение того, зачем информация распределяется в слое 1. Чтобы трансформировать инвариантную репрезентацию в какой-то прогноз, ежесекундно надо решать, куда должен идти сигнал, направленный вниз по иерархии. Слой 1 обеспечивает возможность конвертирования инвариантного представления в более специфическую и конкретную репрезентацию. Например, вы можете воспроизвести Геттисбергскую речь как в устной, так и в письменной форме. Общая репрезентация трансформируется двумя разными способами: первый – для устной, а второй – для письменной формы. Горизонтальный поток активности в слое 1 обеспечивает механизм для осуществления такого процесса. Чтобы инвариантные представления высокого уровня могли транслироваться вниз и принимать форму конкретных прогнозов, мы должны располагать механизмом, позволяющим потоку сигналов разветвляться в каждом слое. На эту роль как нельзя лучше подходит слой 1. Даже не зная о его существовании, мы могли бы предвидеть, что такой слой есть.
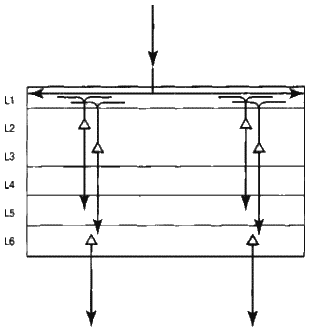
Рис. 6.8. Нисходящий информационный поток
И еще немного сведений из анатомии: аксоны, покидающие слой 6, окутаны оболочкой из белого жирового вещества, которое называется миелином. Это белое вещество выполняет ту же роль, что и изоляция на электрических проводах, – препятствует смешиванию сигналов и повышает скорость их передвижения до 320 километров в час. Аксоны избавляются от белого защитного вещества, попадая в новую колонку нейронов слоя 6 в коре головного мозга.
И наконец, есть еще один непрямой способ связи зон коры головного мозга.
Но прежде я хотел бы вам снова напомнить об автоассоциативном свойстве системы памяти (о котором шла речь в главе 2). Как вы, вероятно, помните, автоассоциативные запоминания служат для сохранения последовательностей сигналов. Когда выходной сигнал группы искусственных нейронов передается назад с целью формирования входного потока для всех нейронов (и при этом обратная связь дополняется задержкой), то таким образом сигналы учатся последовательно двигаться друг за другом. По моему мнению, кора головного мозга использует подобный механизм для сохранения последовательностей, но в нем присутствуют еще некоторые особенности. Вместо того чтобы формировать автоассоциативные последовательности из искусственных нейронов, она формирует их из колонок коры головного мозга. Исходящие данные из всех колонок передаются назад, в слой 1. Выходит, слой 1 располагает свежей информацией о том, какие колонки зоны коры головного мозга только что были активными.
Рассмотрим элементы, изображенные на рис. 6.9. Ученым давно известно, что нейроны особо крупных размеров слоя 5 моторной коры (зона M1) непосредственным образом связаны с мышцами и моторными зонами спинного мозга – они обеспечивают управление вашими мышцами и передвижение. Каждый раз, когда вы говорите, печатаете, выполняете любые самые сложные действия, эти клетки посылают высококоординированные импульсы, заставляя сокращаться ваши мышцы.
Совсем недавно ученые открыли, что крупные нейроны слоя 5, возможно, выполняют какие-то функции и в других частях коры головного мозга, а не только в моторных зонах. Например, крупные нейроны слоя 5 зрительной зоны имеют проекции в зоне, отвечающей за движения глаз. Сенсорные зрительные зоны V2 и V4 не только занимаются обработкой зрительных входных сигналов, но также помогают предопределять движение глаз, а следовательно, управляют тем, что вы видите. Крупные нейроны слоя 5 присутствуют в каждой зоне коры головного мозга, что наводит на мысль об их немаловажной роли во всех типах движений.
Аксоны этих нейронов не только участвуют в формировании поведения, они разветвляются надвое. Одно ответвление идет к таламусу, изображенному на рис. 6.9 в виде овальной фигуры. Таламус человека находится в центре головного мозга, под корой, сверху «старого мозга», и окружен белым веществом. Размером и формой он похож на два маленьких птичьих яйца. Все зоны коры головного мозга направляют многочисленные аксоны в таламус, от него тоже исходят аксоны в обратном направлении. Науке известно достаточно об этих связях, но сам таламус очень сложен по своей структуре, и его роль до сих пор мало изучена. Таламус необходим для того, чтобы вести нормальный образ жизни: его повреждение приводит к устойчивому «растительному» состоянию.
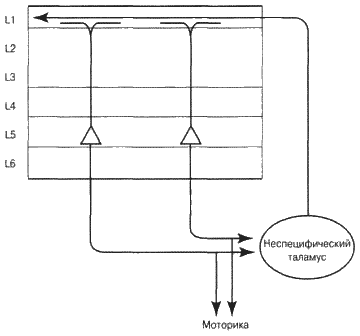
Рис. 6.9. Как посредством таламуса сообщаются текущее состояние и текущее моторное поведение
Существуют несколько путей от таламуса к коре головного мозга, но сейчас лишь один из них представляет для нас интерес. Он начинается с крупных нейронов слоя 5, проецирующихся на группу клеток таламуса, считающихся функционально неспецифическими. Аксоны этих неспецифических клеток идут назад, в слой 1 множества различных зон коры головного мозга. Например, нервные клетки слоя 5 зон V2 и V4 посылают аксоны к таламусу, а таламус, в свою очередь, отправляет информацию назад к слою 1 зон V2 и V4. Аналогичный процесс происходит и в других зонах коры головного мозга. Клетки слоя 5 множества корковых зон посылают сигналы в таламус, который, в свою очередь, отсылает назад информацию в слой 1 этих же самых и связанных с ними зон. Я считаю, что такой круговорот полностью дублирует обратную связь с задержкой, которая позволяет автоассоциативной памяти усваивать последовательности сигналов.
Только что я упомянул о двух источниках входной информации, поступающей в слой 1. Высшие зоны коры головного мозга возбуждают активность в слое 1 более низких зон коры. Активные колонки в пределах одной зоны возбуждают активность в пределах слоя 1 той же зоны через таламус. Эти входящие в слой 1 сигналы можно представить как название песни – считать словами песни (входящие сверху сигналы) и знание, в каком месте песни мы сейчас находимся (задержка в деятельности активных колонок в пределах одной зоны). Таким образом, слой 1 содержит большое количество информации, необходимой нам для прогнозирования времени активизации колонки: название последовательности и наше местонахождение в ней. Используя эти два сигнала слоя 1, зона коры головного мозга в состоянии обучаться и запоминать огромное количество последовательностей сигналов.

