К.ПРИБРАМ
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Перевод с английского Я. Н. Даниловой и Е. Д. Хомской
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
Перевод с английского Я. Н. Даниловой и Е. Д. Хомской
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
KARL H.PRIBRAM Stanford university LANGUAGES OF THE BRAIN Experimental paradoxes and principles in neuropsychology Prentice-hall, inc. Englewood cliffs, new jersey.
ПРЕДИСЛОВИЕ РЕДАКТОРА РУССКОГО ИЗДАНИЯ
Предлагаемая советскому читателю книга принадлежит перу одного из наиболее творческих представителей американской нейропсихологии – профессору Стэнфордского университета К. Прибраму.
Автор этой книги начал свою деятельность нейрохирургом и затем перешел к изучению функций мозга животных. Вместе со своими многочисленными сотрудниками он провел большое число исследований, в которых он пытался выяснить, какую роль в поведении животного играют отдельные структуры головного мозга и как изменяется поведение животного после их разрушения. Работы автора, посвященные функции лобных долей и выработке и удержании программ, направляющих поведение животного, и роли задних (височно-затылочных) отделов мозга в переработке поступающей информации, хорошо известны психологам и физиологам; именно эти работы и выдвинули К. Прибрама на место одного из ведущих авторитетов нейропсихологической науки.
К. Прибрам известен советскому читателю и по ряду его выступлений, получивших широкий отклик. Написанная им совместно с Дж. Миллером и Ю. Галантером книга «Планы и структура поведения» была издана в Советском Союзе (изд-во «Прогресс», М., 1965). В ней был выдвинут новый подход к анализу поведения животных, исходящий из учета той роли, которую играют в поведении «Образы», создаваемые при переработке получаемой информации, и «Планы», лежащие в основе активной деятельности животных и человека. Концепции, предложенные авторами этой книги, близкие к современной кибернетике и учению о саморегулирующихся функциональных системах, развитому в нашей стране П. К. Анохиным, получили большое распространение и были положительно оценены советской научной общественностью. Признание работ К. Прибрама привело к тому, что ему было поручено наряду с виднейшими учеными-психологами выступить с обобщением итогов всех психофизиологических докладов, которые были представлены на XVIII Международном психологическом конгрессе в Москве.
Книга К. Прибрама «Языки мозга» носит совсем особый характер.
С одной стороны, она отличается очень большой широтой и новизной подходов к проблемам мозговой организации психических процессов. Автор привлекает к разрабатываемой им концепции мозговой деятельности новейшие данные физики (голографии), молекулярной биологии, морфофизиологии нейронных структур, учение о саморегулирующихся системах и кибернетику. В этом отношении книга лишена всякой традиционности и не столько резюмирует уже накопленные в науке данные, сколько пытается проложить новые пути, которые, по всей вероятности, будут с достаточной полнотой разработаны лишь будущими поколениями исследователей; однако уже сейчас тот интересный новый материал, который предлагает автор, и уникальные по своей выразительности иллюстрации, которые читатель найдет с первых страниц книги, придают ей большую свежесть и дают важнейшую информацию о последних достижениях в этой области науки.
Пусть многие из высказываемых автором гипотез отличаются чрезмерной смелостью и иногда являются спорными; книга толкает на новые поиски, заставляет пересмотреть многие уже устоявшиеся концепции, и в этом – будящем мысль воздействии – несомненно, одно из основных достоинств книги.
С другой стороны, эта книга, как и другие публикации автора, представляет бесспорный интерес в том отношении, что она отражает попытки выйти из того глубокого кризиса, в котором оказалась американская научная мысль, в течение десятилетий испытывавшая тормозящее влияние упрощенных и механистических схем бихевиоризма.
Как и в прошлой книге, написанной совместно с Дж. Миллером и Ю. Галантером, автор решительно отходит от бихевиористского понимания поведения как цепи стимулов и реакций; он столь же решительно отбрасывает самую мысль о возможности игнорировать проблемы сознания, с одной стороны, и анализ физиологических механизмов, лежащих в основе сознательной деятельности, – с другой.
Пусть самая возможность понять сознательную деятельность как продукт сложнейшего общественно-исторического развития еще чужда автору (который в большой мере остается биологом и пытается биологически и нейрофизиологически трактовать механизмы, лежащие в основе психической деятельности). Пусть он иногда определяет свою позицию как позицию «субъективного бихевиоризма» (иначе говоря, изучение поведения, не исключающее, а предполагающее анализ субъективного мира и ищущее его нейрофизиологических объяснений); пусть он высказывает ряд положений, которые, очевидно, вызовут острые дискуссии. Однако его решительный отход от упрощенных, механистических схем бихевиоризма, попытки проложить новые пути в нейропсихологии представляют несомненный интерес, и именно эти попытки дают читателю возможность увидеть контуры будущей психофизиологии.
Остается пожалеть, что автор недостаточно знаком с успехами советской физиологии (в частности – с работами П. К. Анохина, ряд положений которого почти полностью совпадает с его взглядами) и с теорией деятельности, развиваемой в советской психологической науке. Более близкое знакомство с данными обеих дисциплин существенно помогло бы ему уточнить ряд высказываемых им идей.
Можно не сомневаться в том, что книга К. Прибрама, в которой автор пытается разобраться в том, на каких «языках» говорит мозг и в каких параметрах выражается его деятельность, будет с пользой прочитана советскими исследователями и послужит для них толчком к новым продуктивным размышлениям.
А. Р. Лурия
© «Прогресс», 1974
ПРЕДИСЛОВИЕ АВТОРА К АНГЛИЙСКОМУ ИЗДАНИЮ
Появление книги «Языки мозга» вызвано несколькими обстоятельствами. Во-первых, существует профессиональная потребность сформулировать для самого себя и своих коллег научные взгляды, которые сложились во время моей работы и сейчас направляют ее. Таким образом, являясь прежде всего теоретическим исследованием, книга содержит анализ тех парадоксов и загадок, которые неожиданно возникали в ходе экспериментов. В то время, когда проводилось исследование, ряд фактов не поддавался объяснению в рамках теории, разделяемой большинством исследователей. Поскольку попытки найти объяснение таким парадоксам придавали своеобразие научным достижениям лаборатории, изучающей отношение между мозгом и поведением, я постарался сохранить его в этой книге.
Но не одни лишь профессиональные интересы побудили меня написать эту книгу в ее настоящем виде. Обилие новых данных и важность разработки проблем взаимоотношения мозга, поведения и сознания вызвали в последние годы интерес к этим проблемам со стороны широкого круга людей. К ним относятся не только специалисты по психологии или неврологии, но и молодые, начинающие исследователи, студенты, инженеры и физики, биологи и биохимики, социологи и психиатры, философы и др. Такая аудитория требует чего-то большего, чем «самые последние данные о базолатеральном ядре амигдалы» или «различии между задачами на активное и пассивное избегание». Интуитивно она догадывается, что открытия в области изучения мозга м поведения важны не только «для науки», но и для самих читателей.Эта книга и является попыткой разрешить многие «опросы, возникающие перед теми, кто, как и я, старается дать объяснение тем парадоксальным результатам, которые были получены при изучении мозга и поведения.
Поэтому я попытался найти в этой книге такую манеру изложения, которая сочетала бы в себе описание деталей» интересующих профессионалов, с тем, что представляет всеобщий интерес. Основное направление моих лабораторных исследований создает идеальные условия для выполнения этой задачи: цель моих экспериментов состоит в сущности в определении посредством поведенческого анализа функций различных систем нейронных структур, составляющих мозг. Эта «системная» нейропсихология является связующим звеном между нейрофизиологией – электрическим и химическим изучением функций нервных клеток (и их частей)-и экспериментальной психологией – поведенческим анализом функций организма как целого. За последние годы появилась и третья линия исследований, связанная с использованием компьютеров при изучении многих интересующих нас проблем. Компьютеры помогают контролировать эксперименты, анализировать полученные данные и указывают новый путь для будущих исследований. Можно существенно сэкономить усилия исследователей, если дать какому-либо научному подходу «зеленую улицу», чтобы проверить пригодность тех или иных методов и подробно описать последствия их применения, которые далеко не очевидны, хотя и подразумеваются. Закон, выведенный с помощью компьютера, имеет для биолога, изучающего поведение, такое же значение, как эксперимент in vitro (в стеклянной пробирке) для биохимика. В обоих случаях такое моделирование способствует разработке системы научных понятий, с помощью которых можно адекватно анализировать процессы, протекающие в живых организмах.
Поэтому такой «системный» подход к проблемам нейропсихологии волей-неволей предполагает знакомство исследователя с достижениями других дисциплин, имеющих свой собственный круг проблем и разработавших соответствующие понятия и свой подход к решению этих проблем. Короче говоря, нейропсихолог, стоящий на такой позиции, должен пользоваться разными «языками».
Результаты моих собственных исследований подтвердили необходимость прибегать к множеству различных «языков». Очевидно, мозг организует перцептивные, моторные и мнестические процессы путем многократного перекодирования получаемой информации и многократной перестройки своей собственной активности. Сенсорные возбуждения трансформируются в динамические структуры нервной активности, которые позволяют сохранять информацию без ее чрезмерной потери. Дальнейшее преобразование в другие нервные структуры, в другие нервные «коды» происходит при последующей «переработке информации» и формировании сложных форм поведения. Поэтому многое в моей работе связано с анализом набора тех мозговых кодов, тех «языков» мозга, которые используются на той или иной фазе психологической переработки. Какие мозговые коды делают возможным опознание зрительного образа? Какие мозговые коды координируют постройку гнезда или искусное исполнение фортепьянной сонаты? Какие мозговые коды создают ощущение голода, сонливость, апатию или заинтересованность? Каковы те операции мозгового кодирования, которые позволяют мозгу одного человека общаться с мозгом другого? Что же представляют собой «языки мозга»?
В этой книге предпринята попытка определить общие принципы мозговых кодирующих механизмов и преобразований, используемых при перекодировании. В ходе работы мы столкнулись с целым рядом парадоксов: экспериментальные результаты порой настолько отличались от того, что мы ожидали, что мы неизбежно должны были усомниться в правильности наших прежних представлений о работе мозга. Естественно, что это заставляло нас разрабатывать новые теории и предлагать новые гипотезы.
Книга «Языки мозга» использует биологический язык химии, физиологии, неврологии, язык психологов, изучающих поведение, и язык инженеров, имеющих дело с компьютерами. Но поскольку эти языки нужно было сделать понятными для широкой аудитории, которая включает в себя как представителей различных дисциплин, так и людей, не являющихся специалистами, мы сохраняем в этой книге лишь основные понятия и слова всех этих языков.
Книга состоит из четырех частей, каждая из которых относительно независима от других и может служить введением к последующей, – учет специфических интересов отдельных групп читателей мог бы продиктовать другую структуру книги.
Итак, в книге «Языки мозга» излагаются принципы мозгового кодирования. В каждой из ее четырех частей исследуются различные проблемы. Первая часть посвящена основной функции мозга и «логике» построения нейронных структур, которые позволяют мозгу формировать применяемые им коды. Во второй части показана роль мозга в организации психических процессов. В ней анализируется процесс кодирования, участвующий в восприятии, мотивации и эмоции. В третьей части рассматриваются проблемы нейронного контроля и пластичности поведения. И, наконец, последняя часть посвящена структуре коммуникативных процессов в терминах знаков, символов и внутренней речи, которые регулируют действия человека.
Таким образом, в книге показана необходимость мозга формировать различные коды, что приводит к появлению разных языков и превращает интеллектуальное сообщество в довременную Вавилонскую башню; вместе с тем она предоставляет и средства, позволяющие преодолеть это затруднение. В дискуссиях о тех качествах, которые делают человека человеком, исследователи часто пользуются дихотомиями: сознание – тело, сознание – машина, сознание – мозг, сознание – поведение. На страницах этой книги я попытался подойти к анализу таких дихотомий, исходя из того, что они являются выражением различных «языков мозга»_
Я благодарен прежде всего тем авторам, которые за последнее время написали ряд работ по физиологической психологии. Их успех помог мне найти свою собственную манеру изложения «Языков мозга». Это сделало книгу дополнением к общепринятому пониманию отношений между мозгом и поведением.
В создании этой книги принимали участие многие исследователи. В ней цитируются работы тех, кто в течение ряда лег вместе со мной создавал лабораторию. Многие другие помогли» мне уточнить мои концепции в дискуссиях, подробные ссылки на их работы будут даны в следующей книге, за которую я теперь принимаюсь; там же будут освещены более специальные вопросы, разрабатываемые в моей лаборатории.
«Известно, что когда Клерк Максвелл был: ребенком, у него была мания требовать, чтобы ему все объясняли; и, когда люди отделывались от него с помощью туманных словесных объяснений, он нетерпеливо прерывал их, говоря: «Да, но я хочу, чтобы вы мне сказали, что же из этого следует!» Поскольку его интересовала истина, то только прагматик мог бы сказать ему, что именно следует из сказанного… Истина возникает из фактов, но она и опережает факты, и кое-что к ним прибавляет, а эти факты вновь создают или открывают истину… и так до бесконечности. Между тем сами по себе «факты» не составляют истины. Они просто существуют. Истина состоит в той уверенности, которая начинается с фактов и кончается ими».
Уильям Джеме
ЧАСТЬ ПЕРВАЯ ФУНКЦИИ МОЗГА: МЕХАНИЗМ ДВУХ ПРОЦЕССОВ
«Язык, с помощью которого передается информация [в мозге] … не соответствует и не должен соответствовать тому языку, которым люди пользуются в общении друг с другом».
Питтс и Мак-Наллок
Глава I
СОСТОЯНИЯ И ОПЕРАТОРЫ
ВВЕДЕНИЕ
«Я вас люблю». В Париже была весна, и эти слова были произнесены с прелестным скандинавским акцентом. Это событие произошло на заседании ЮНЕСКО, посвященном проблемам изучения мозга и человеческого поведения. Слова эти были произнесены не белокурой красавицей, а маленьким блестящим металлическим устройством, которое держал в руках известный психолингвист.
Устройство поразило всех нас простотой своей конструкции. Громкоговоритель регулировался с помощью только двух ручек. Одна изменяла состояние электронной цепи, которое моделировало напряжение голосовых связок; другая регулировала импульсы, генерируемые цепью и имитировавшие взрывные звуки воздушной струи, ударявшейся по связкам.
Может ли это простое устройство иметь отношение к изучению человеком самого себя? Нельзя ли вызывать и контролировать все поведение с помощью столь же простого нейронного механизма? Не является ли нервнйя система «двухкнопочным» механизмом двойного процесса, в котором один процесс выражается в терминах нейроэлектрических состояний, а другой – в терминах особых пульсирующих операторов, воздействующих на эти состояния? То, что нервная система фактически действует посредством импульсов, хорошо известно. Существование нейроэлектрических состояний мозга также было установлено, но доказательства их существования были получены не сразу, и понимание их значения для изучения поведения приходило лишь постепенно даже в нейрофизиологии. Поэтому в первой главе рассматриваются данные, свидетельствующие о правдоподобии двухпроцессной модели функций мозга. .
Чтобы понять, в чем заключаются функции мозга, мы должны вначале понять, из каких единиц состоит нервная система. Классический анализ определяет эти единицы как нейроны – клетки, которые структурно и функционально изолированы друг от друга мембранными оболочками. Однако такое упрощенное представление об изолированном нейроне как единственном организаторе мозговых процессов вызывает затруднение при попытке охарактеризовать нейроэлектрические состояния как часть такого двойного механизма. В двух первых разделах дается беглый обзор основ нейронной теории и тех причин, по которым в настоящее время в нее должны быть внесены некоторые поправки.
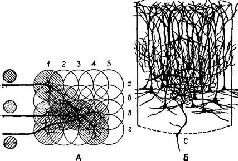
В конце XIX в. в нейробиологии возникла полемика о том, состоит ли мозговая ткань из единиц – клеток, подобно всем другим тканям тела. Результаты этой полемики были настолько убедительны, что нейрон и его способность действовать как единица, разряжающаяся электрическим потенциалом по закону «все или ничего», перестали быть предметом теоретического рассмотрения. Еще никто не «видел» нейрон мозговой ткани; никто не проследил полностью его протяженность и не показал, что он действительно изолирован от других нейронов (рис. 1-1).
Рис. 1-1. Сканирующая электронная микрофотография, показывающая расположение нервных волокон в сетчатке Necturus'a. Волокна (дендриты) берут начало от внутреннего членика колбочки и распространяются в сторону наружного. Следует отметить, что точки контакта необязательно находятся на нервных окончаниях (Lewis, 1970).

Каким же образом нейронная теория получила в дальнейшем всеобщее признание и почему мы вновь вернулись к этой проблеме сегодня? Спор вокруг нейронной теории достиг своей кульминации в конце XIX в. Непрерывность нервной ткани признавали такие известные нейроанатомы, как Билыповский и Гольджи; Валытейер и Рамон-и-Кахал рассматривали нейрон как независимый, но соприкасающийся с другими нейронами. Однако убедительный анализ был дан не этими нейроанатомами, а сэром Чарлзом Шеррингтоном в его лекциях в Йельском университете, опубликованных как классическая работа «Интегративная деятельность нервной системы». Ранее Шеррингтон совместно с Фостером предложил термин «синапсы» для обозначения предполагаемых связей между отдельными нервными клетками. Он внес большой теоретический вклад в нейронную теорию, противопоставив нейрофизиологические данные, основанные главным образом на изучении электрических явлений в нервных стволах, нейропсихологическим, базирующимся на изучении рефлекторной деятельности у «спинального животного» (нашедшей отражение в концепции рефлекторной дуги, см. гл. V). Шеррингтон высказал мысль, что парадоксальные расхождения результатов нейрофизиологических и поведенческих исследований могут получить объяснение, если обратиться к нейроанатомическому описанию синапсов. Основные расхождения состояли в том, что (1) в изолированном стволе сигнал, вызванный стимуляцией какой-нибудь его точки, передается в двух направлениях, тогда как проведение сигнала при осуществлении спинального рефлекса происходит в одном направлении, и (2) скорость проведения импульса по нервному стволу значительно выше, чем скорость ответа, получаемого в результате рефлекса. Помимо этих, было отмечено девять других расхождений. Большинство из них касалось степени соответствия между адресованным организму стимулом и полученным ответом: соответствие всегда было большим для нервного, чем для поведенческого ответа. Чтобы объяснить эти расхождения, синапсам приписывались такие предполагаемые функции, как пространственная конвергенция и временная суммация, утомляемость, облегчение и торможение. Следующее утверждение Шеррингтона дает представление о его концепции:
«Наиболее характерными особенностями проведения в рефлекторных дугах по сравнению с нервными стволами являются: 1) меньшая скорость, если судить по измерениям латентного периода между нанесением раздражения и появлением конечного эффекта, причем это различие больше для слабых раздражителей, чем для сильных; 2) менее близкое соответствие между моментом прекращения раздражения и моментом прекращения конечного эффекта (т. е. имеет место отчетливое «последействие»); 3) менее близкое соответствие между ритмом раздражения и ритмом конечного эффекта; 4) менее близкое соответствие между градуальным увеличением интенсивности раздражения и градуальным увеличением интенсивности конечного эффекта; 5) значительное сопротивление к прохождению нервного импульса, которое, однако, без труда преодолевается последовательностью импульсов (временная суммация); 6) необратимость направления проведения по сравнению с обратимостью в нервных стволах; 7) утомляемость в противоположность сравнительной неутомляемости нервных стволов; 8) более значительная изменчивость пороговых величин раздражения по сравнению с нервными стволами; 9) рефрактерный период,» проторение», торможение и способность впадать в состояние шока в степени, неизвестной для нервных стволов; 10) большая зависимость от кровообращения и снабжения кислородом (Ферворн, Винтерштейн, Бейер и др.); 11) более выраженная восприимчивость к действию различных лекарств-анестетиков» (Шеррингтон, 1969, стр. 40-41).
Короче говоря, результаты электрофизиологического исследования нервных стволов и поведенческого изучения рефлекса не совпадали. Нервы казались простыми проводящими путями для импульсов; рефлекс же обнаружил такую сложность организации, которая не могла быть объяснена простой «проволочной» моделью. Поэтому Шеррингтон принял «нейронную теорию», высказав предположение, что «провода», образующие нервную систему, составляют не непрерывную сеть, а слегка отделены друг от друга и включают структуру, которую он назвалсинапсом. Затем он наделил синапс всей той сложностью, которую необходимо принимать во внимание при объяснении рефлекторного поведения. Таким образом, свойства изучаемого поведения были отнесены за счет свойств соединений между нейронами, а не их проводниковой части (рис. 1-2 и 1-3).
Рис. 1-2. Скопление нейронов в сером веществе спинного мозга обезьяны, иллюстрирующее обычное расположение тел нервных клеток и их аксонных и дендритных отростков. Большая часть пространства между телами клеток занята нервным сплетением, состоящим из этих отростков и множества входящих и разветвляющихся аксонных окончаний. Сплетение аксонов и дендритов, посредством которого главным образом осуществляется связь одного нейрона с другим, известно под названием «нейропиль». р – перикарион; а – аксон; d – дендрит; пр – нейропиль; л – ядро нейрона большого размера, вероятно мотонейрона. Парафиновый срез, 1,5м, окраска по Бодиану (Х150). (Видимое пространство вокруг тел нервных клеток – артефакт в результате сморщивания ткани). (Bodian, 1967.)

Рис. 1-3. Схема микроструктуры синаптических связей в коре. Частично перекрывающие друг друга окружности изображают область соединительных контактов между разветвлениями входных аксонов и кортикальных дендритов (Scheibel and Scheibel, см. Chow and Leiman, 1970).
Теоретическая гипотеза Шеррингтона в значительной степени игнорировалась исследователями в области нейрофизиологии и нейропсихологии. Внимание было сосредоточено на существовании и свойствах синапсов. Эти свойства детально изучались с помощью электронной микроскопии и записи электрической и химической активности соединений. Однако интерпретация этих результатов почти не выходила за рамки вопроса: «Как осуществляется прохождение нервных импульсов через барьер синапса?» Ссылка на разряды, генерируемые клеткой, и на проводящие свойства синаптических путей слишком часто рассматривалась как самоочевидная и достаточная.
Немногие ученые придерживались тезиса Шеррингтона о том, что сложность поведения (и психологических процессов) следует объяснять сложностью организации соединительных (синаптических) механизмов в центральной нервной системе.
Цель данной книги состоит в том, чтобы восполнить этот пробел. В ней сделана попытка, исходя из идей Шеррингтона, решить проблему взаимоотношений между мозгом, сознанием и поведением, подобно тому как Шеррингтон решал проблему отношений между спинным мозгом и рефлексом. Проблемы, возникающие при изучении головного мозга, тоньше и сложнее, чем проблемы, связанные со спинным мозгом. Тем не менее можно уже теперь достигнуть значительных успехов в понимании этих проблем при осторожном использовании классического нейробиологического подхода Шеррингтона. Однако неврологи, физиологи и психологи часто его игнорируют, переходя от отчаяния к неопределенным надеждам, а порой даже вообще отрицают существование такой проблемы. Изучение отношений между мозгом, сознанием и поведение» может быть продуктивным только в том случае, если ставятся ограниченные проблемы. Каковы же эти проблемы?
НЕКОТОРЫЕ НЕЙРОПСИХОЛОГИЧЕСКИЕ ФАКТЫ
Хотя нейрофизиологический эксперимент основывался на нейронной теории, некоторые известные психологи неоднократно отмечали, что понимание нервной системы как функционирующей исключительно в виде набора дискретно проводящих единиц не согласуется с полученными ими экспериментальными данными. Эти исследователи объясняли свои наблюдения, прибегая скорее к той или иной модели поля, чем к квантовой, дискретной, вероятностной нейронной теории. Разногласия, коротко говоря, сводились к следующему:
«Физиолог слепо верит в то, что, поскольку мозг состоит из нейронов, он способен только к такому возбуждению, которое является суммой возбуждений многих нейронов, и что эти центральные нейроны подчиняются тем же законам и возбуждаются при тех же условиях, что и периферические нейроны, которые были изучены экспериментально. Этому утверждению физиолога психолог иногда противопоставляет другое мнение, а именно, что организация центрального возбуждения протекает иначе, что оно определяется рамками получаемого индивидом опыта» (Boring, 1932, р. 32).
Проблема возникает особенно тогда, когда проводятся нейропсихологические эксперименты, предполагающие разрушение мозга. Результаты этих экспериментов послужили основанием для защиты двух противоположных точек зрения. По мнению одних авторов, каждая кортикальная точка, каждая клетка или группа клеток специализируется на выполнении какой-нибудь одной функции. Интеграция, необходимая для объяснения поведенческих и психологических процессов, согласно этой точке зрения, достигается за счет врожденных или образующихся при жизни прочных ассоциативных связей между нейронами.
Другая точка зрения, которой придерживается и автор этой книги, состоит в том, что в мозговой ткани возникают определенные важные для организации поведения и психологического процесса взаимодействия и эти взаимодействия нельзя рассматривать только как постоянные ассоциативные связи между нейронами. К. Лешли, первый сторонник этой точки зрения, построил свою аргументацию по трем линиям: 1) эквивалентность рецепторной, функции; 2) спонтанная реорганизация моторных реакций и 3) сохранение поведения после разрушения любой части мозга, в то время как лишь разрушение всей мозговой системы устраняет раз возникшее поведение. Остановимся на этих трех линиях доказательства.
Что касается эквивалентности рецепторной функции, то эксперименты показывают, что «абсолютные характеристики стимула имеют сравнительно небольшое значение для поведения и что реакция определяется отношениями возбуждений, которые при возникновении в любой группе рецепторных клеток этой системы имеют одинаковый эффект». Например, «животное, тренированное в выборе большего из трех кругов, может сразу же положительно реагировать на поле с наиболее широкими линиями при сравнении трех полей с полосами различной ширины» (Lashley, 1960, 238-239). Подробные данные такого рода содержатся во второй части книги; сейчас же достаточно отметить, что такая проблема существует.
Что касается спонтанной моторной реорганизации, то «результаты показывают, что, когда привычно используемые двигательные органы перестают функционировать вследствие их удаления или паралича, наблюдается непосредственное спонтанное использование других моторных систем, которые ранее не были связаны с этой деятельностью или не использовались при ее осуществлении» (Lashley, 1960, р. 239). Например, основной рисунок мышечного напряжения во время .акта письма, когда человек держит карандаш в пальцах, сохраняется и тогда, когда он переходит к письму, зажав карандаш зубами. Каким образом мозг обеспечивает это, составляет содержание первых глав третьей части этой книги.
Доказательство взаимозаменяемости частей функциональных систем организма в организации поведения и психологических процессов распространяется также и на центральные отделы мозга. Это доказательство содержится во второй части. Здесь же лам необходимо только отметить, что даже значительное повреждение мозга может оказать лишь небольшое влияние на то крайне сложное поведение, которое, казалось бы, должно быть особенно чувствительно к разрушению, если считать, что интеграция целиком зависит от наличия постоянных ассоциативных связей.
Лешли (см. Beach и др., 1960) следующим образом резюмирует проблему:
«Эти три линии доказательства показывают, что некоторые хорошо координированные виды деятельности, казалось бы зависящие от определенных областей коры, могут в известных пределах осуществляться любой частью коры. Это может происходить в результате наличия многих дублирующих рефлекторных путей, проходящих через кору, и такое объяснение, по-видимому, можно дать всем известным случаям сохранения функций после частичного разрушения определенных областей, имеющих отношение к этим функциям. Но оно не годится для объяснения фактов сенсорной и моторной эквивалентности. Эти факты говорят о том, что, если была выработана условная реакция (например, положительная реакция на определенную структуру зрительного сигнала), эта реакция будет вызываться возбуждением сенсорных клеток, которые раньше никогда не стимулировались таким способом. Точно так же однажды приобретенные двигательные акты (например, открывание задвижки ящика) могут быть сразу же выполнены двигательными органами, которые не включались ранее в выполнение этих актов» (р. 237-240). В утверждении Лешли смешаны по крайней мере два вопроса: взаимозаменяемость частей организма и перенос навыка, который? включает в себя проблему сходства и новизны. Тем не менее приведенные здесь высказывания Лешли говорят о необходимости согласования данных психологии с данными нейрофизиологии. Такое согласование предполагает более широкое использование фактов, касающихся работы мозга, чем то, которое возможна сейчас вследствие узкого толкования нейронной теории. Между тем окончательная точка зрения на функции мозга должна быть основана на классических нейрофизиологических данных. Поэтому остановимся на тех последних успехах нейрофизиологической техники, которые показывают, что такое согласование действительно возможно (Bullock, 1959). Этот путь позволяет провести различие между первичными структурами интранейронных нервных импульсов и первичными интернейронными структурами активности; такой путь значительно расширяет диапазон тех подходов, которые применяются для изучения функций мозга.
МИКРОСТРУКТУРА СОЕДИНЕНИЙ
Значение нейроэлектрических процессов, генерируемых в синапсе и позади него, стало очевидным для нейрофизиологов после того, как были получены определенные данные. В течение многих лет электрическая активность, отводимая от кожи черепа, рассматривалась как отражение совокупности нервных импульсов, генерируемых нейронами мозга, лежащими где-то ниже участка записи. Когда стала возможной одновременная регистрация электрической активности локальных участков мозговой ткани и их совокупности, это предположение стало вызывать сомнение (Purpura, 1958). Запись электрической активности нейрона при внутриклеточном отведении показывает, что даже при отсутствии широко распространяющихся нервных импульсов возникают ритмические изменения медленных потенциалов, и, когда их сравнивают с ритмами, одновременно записанными от группы нейронов той же самой точки, они совпадают (Creutzfeldt et al., 1966; Fugita and Sato, 1964; Elul and Adey, 1966; Morrell, 1967). Когда мембрана нервной клетки подвергается поляризации постоянным током, амплитуда внутриклеточных ритмов меняется и это изменение отражается в суммарной записи (Eccles, 1964, см . рис. 1-4).
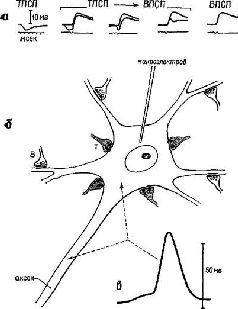
Рис. 1-4. Схема нейрона (б) с возбудительными (Б) и тормозными (Т) синапсами и внутриклеточные записи (а) медленных постсинаптических потенциалов (ВПСП и ТПСП) – вверху. Следует отметить, что импульс, генерируемый на начальной части аксона, имеет амплитуду выше 50 мВ, тогда как амплитуда внутриклеточных записей имеет диапазон около 5 мВ. Только нервному импульсу аксона свойственно распространение (см. Eccles, 1967).
Таким образом, было получено серьезное доказательство того, что-главный вклад в электрическую активность, записываемую суммарно в виде электроэнцефалограммы (ЭЭГ), вносится медленно возникающими флуктуациями потенциалов нейронных мембран (рис. 1-5).

Рис. 1-5. Записи медленных дендритных потенциалов (справа) с указанием примерного уровня их регистрации в коре. Калибровка 0,2 мВ; 10 мсек (Purpura, 1967).
Рис 1-6. Диаграмма связи между внутриклеточными записями медленных потенциалов и ЭЭГ, записанной с поверхности коры той же самой области мозга, для 500-секундного периода времени (Adey, 1967).
Только тогда, когда активность большого числа единиц синхронизирована – когда их разряды вызываются вспышкой света или звуковым щелчком, – суммарная запись и импульсы, генерируемые отдельными нейронами, оказываются одинаковыми (рис. 1-7 и 1-8).

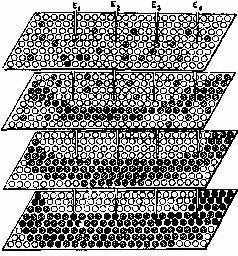
Рис 1-7 и 1-8. Отношения между синхронизацией волн ЭЗГ, нейронными разрядами и циркуляцией нейронной активности при одновременной записи четырьмя микроэлектродами, удаленными друг от друга на 100- 150 м- Внизу: двухмерная схема нейронных полей, показывающая нейронную активность каждой последовательной стадии (Verzeano et al., 1970).
В одном таком исследовании (Fox and O'Brien,. 1965) животному предъявлялось много световых вспышек (от 3000 до 5000). Когда разряды нейрона были усреднены для всего числа предъявлений, вероятность разряда одиночной клетки зрительной коры в период, следовавший за вспышкой (в течение 1 сек), приблизилась к волновой форме симультанно записанного суммарного потенциала. Таким образом, суммарный потенциал указывает на вероятность, с которой будет разряжаться нейрон после предъявления кратковременного стимула, способного синхронизировать группы нейронов. Однако при отсутствии столь отчетливо выраженной синхронизации суммарная запись отражает электрические изменения потенциала, возникающие в сплетении нервных волокон, которые представляют собой нервную ткань, но фактически не слособны вызывать спаиковые разряды нейронов (Li, Cullen, and Jasper, 1956; Creutzfeldt, 1961; Verzeano and Negishi, 1960; см. рис. 1-5 и 1-6).
Эти экспериментальные данные заставляют фактически различать два типа нейроэлектрической активности: импульсные разряды нейрона, с одной стороны, и градуальные изменения медленного потенциала – с другой. Распространяются только нервные импульсы; градуальные изменения усиливаются и ослабевают в мозговой ткани локально, они чувствительны к различным влияниям, таким, как локальная химическая среда, которая по своей природе не является строго нейронной.
Различие между градуальными медленными потенциалами и нервными импульсами зависит частично от того, что амплитуда и скорость проведения нервного импульса пропорциональны диаметру нервного ствола, в котором возникает импульс. Таким образом, в нервных волокнах большого диаметра импульсы имеют, как правило, значительную амплитуду и быстро распространяются. В нервных волокнах с малым диаметром амплитуда электрического потенциала нервного импульса меньше (по крайней мере при экстраклеточном отведении) и скорость его проведения ниже. По мере удаления нервных волокон от тела клетки они утончаются и разветвляются; в результате этого импульсы становятся все более слабыми и замедленными, особенно на окончаниях волокон (рис. 1-9).
Рис. 1-9. Основные типы нейронов в центральной нервной системе млекопитающих в соответствии с выполняемой ими функцией, иерархическим уровнем и вероятной степенью разветвленности их отростков (Bodian, 1967).
Поэтому необходимы химические медиаторы, которые усиливали бы изменение потенциала, возникающее в нервных окончаниях, чтобы электрический сигнал мог распространить свое действие через структурные соединения между нейронами. Несмотря на химический усилитель, потенциалы, генерируемые позади синапса (названные постсипаптическими потенциалами), первоначально возникают как рудиментарные медленные градуальные изменения, подверженные влиянию химического посредника, создаваемого текущим по сосудам током крови и не нейронными (глиальными) клетками, окружающими волокна. Кроме того, более тонкие нервные волокна теряют толстое изолирующее покрытие (миелиновую оболочку), которая у крупных волокон препятствует взаимодействию импульсов. Медленные потенциалы, возникающие в таких тонких волокнах, как разветвления дендритов, следовательно, не встречают препятствий для реального локального взаимодействия.
Упрощенная до некоторой степени картина (рис. 1-10) взаимоотношений между изменениями медленных потенциалов и нервными импульсами выглядит следующим образом: внутри нейрона электрохимические процессы ведут к деполяризации мембраны – возрастанию градуальных потенциалов, пока они не становятся нервными импульсами. Там, где соединяются нейроны, или где тонкие, немиелинизированные волокна формируют сплетение и образуют контакты относительно бесструктурным способом, названным эфаптическим (рис. 1-11), или где окончания волокон действительно образуют структурный контакт с другими нейронами посредством синапса (рис. 1-12), нервные импульсы постепенно затухают, то есть превращаются в небольшие медленные потенциалы, не отличимые от спонтанно генерированной локальной градуальной активности (см. гл. III).
Рис. 1-10. Схема, показывающая отношения между градуальными медленными потенциалами, нервными импульсами и химической секрецией, которая действует через структурированные соединения между нейронами (синапсы) как химический возбудитель (Grundfest, 1967).
Короче говоря, эффективный интранейронный механизм характеризуется тем, что возрастание градуальной активности приводит к появлению распространяющихся нервных импульсов.
Эффективный механизм соединений характеризуется противоположными чертами: нервные импульсы затухают, трансформируясь в локальные медленные потенциалы. Соединительный механизм в свою очередь влияет на интранейронную мембранную деполяризацию, но только после того, как реализуется возможность пространственных и временных взаимодействий между конфигурациями медленных потенциалов. Именно в этих взаимодействиях между конфигурациями и состоит специфическое значение медленных потенциалов.
Такая точка зрения на активность соединений несколько отличается от широко распространенного понимания синаптической функции. В большинстве нейрофизиологических исследований синапса ставился вопрос: «Каким образом нервные импульсы или передаваемая ими информация преодолевают синаптическую щель?» Настоящий подход обращает внимание на медленные потенциалы, как таковые; в основе его лежит предположение, что медленные потенциалы образуют динамические структуры, которые выполняют какую-то функцию, помимо той роли, какую они играют в передаче импульсов. Согласно принятой здесь точке зрения, структуры медленных потенциалов «вычисляют» взаимодействие между соседними нейронами в пространстве и в какой-то мере их взаимодействие во времени скорее с помощью непрерывного (аналогового), чем дискретного (цифрового) механизма, работающего по принципу «все или ничего».
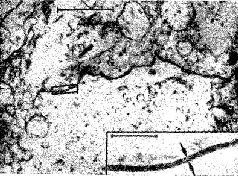
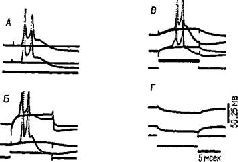
Рис. 1-11. Электрофизиологическое доказательство (представленное внутриклеточными записями двух нейронов) эфаптической передачи через неструктурированные контакты между нейронами, выявленные с помощью электронной микроскопии. Залпы электрических импульсов (В, В) и переменный ток (Г) воздействовали на одну клетку, а запись активности производилась от другой. Слабая стимуляция одной клетки (А) вдали от другой вызывала импульсы почта одновременно в обеих клетках (Grundiest, 1967).

Рис. 1-12. Синаптические контакты на пересечении двух нервных волокон в нейропиле брюшного ганглия Aplesya Californica. Увеличение примерно в 35 000 (Lewis, 1970).
Совокупность медленных потенциалов, в любой момент присутствующая на обширном участке, может быть описана как состояние, имеющее некоторую микроструктуру. Приход импульсов на эфаптические или синаптические соединения никогда не бывает одиночным. Аксонные окончания обычно многочисленны, ибо аксоны ветвятся на своих концах. Соединение между двумя нейронами может осуществляться с помощью большого количества синапсов, доходящего до 1000. Дендриты, уже по определению напоминающие деревья, обладают многочисленными тонкими волокнистыми ветвями, которые, пересекаясь, образуют множественные контакты между нейронами. Эти контакты у некоторых клеток (например, амакриновых клеток сетчатки) включают структурные синапсы (рис. 1-13).
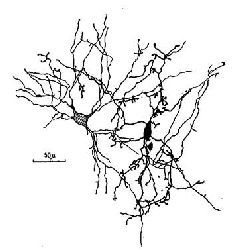
Рис. 1-13. Зарисовка, показывающая многочисленные точки контакта (синаптического и эфаптического) между двумя нейронами мозга (Ralston, 1968).
Таким образом, эфаптические и синаптические процессы, возникающие в местах соединений между нейронами, формируют определенную динамическую структуру, заключение о природе которой может быть сделано на основе знания тонкой структуры мозга и его электрической активности. Можно, в частности, предположить, что эти динамические структуры создают волновые фронты. Это предположение становится особенно полезным при рассмотрении проблем эквивалентности (см. вторую часть книги). Это не значит, что микроструктуру медленного потенциала следует рассматривать в терминах механической волны, но иногда это сходство полезно иметь в виду. Ведь именно интерпретация динамических структур медленных потенциалов как образующих волновые фронты помогает представить себе то, что происходит.
«Нейрон – это реле, действующее по закону «все или ничего». Импульс, достигающий синапса, вызывает очень небольшой и временный электрический эффект равный 0,001 вольта и длящийся от 0,01 до 0,02 сек. Необходимо примерно в 10 раз большее возбуждение, чтобы вызвать разряд Нейрона.
Так как требуется конвергенция многих импульсов на какой-нибудь «дин нейрон, чтобы вызвать его разряд, то цепи, состоящие из одиночных нейронов, не могут обеспечить распространение волны активности в коре.
Распространение активности представляет собой, скорее, передний фронт движения во многих направлениях, когда параллельно активизированы многие клетки в каждом синаптическом звене цепи… (рис. 1-14).

Рис. 1-14. На этой диаграмме синаптические области коры изображены в виде кружков на плоскости. Движение процесса, которое может осуществляться во многих направлениях и характеризуется специфической нейронной структурой возбуждения, показано черными кружками для одного направления и темно-серыми для другого. Области с контурными кружками не активизируются ни одной из динамических структур. Следует отметить, что на пересечении этих двух направлений движения одни и те же области принимали бы участие в обоих процессах и поэтому каждая из них была бы изображена в виде очень большого серого кружка, а не как темно-серый или черный кружок, как показано на рисунке (Eccles, 1970).
Мы сразу же получаем объяснение одного замечательного свойства нервной сети: каким образом два совершенно различных входных воздействия (одно на клетки А, и А2, другое – на клетки А3 и А4) могут быть переданы посредством одной и той же динамической структуры клеточных связей, противодействуя друг другу и достигая совершенно различных выходов (Г3 – Г4) . . . (рис. 1-15).
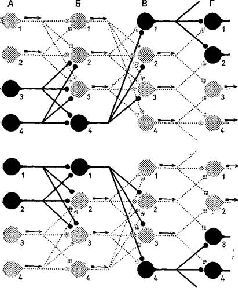
Рис. 1-15. Модель очень схематизированной нейронной сети, иллюстрирующая простейший случай проведения в системе с множеством направлений. На верхней и нижней диаграммах представлены нервные чзети с одной и той же анатомией. В колонках А, Б и В изображены синаптические контакты 12 клеток, клетки генерирующие импульс (со стрелками) – светло-серого цвета, молчащие клетки – черные. Предполагается, что клетка разряжается импульсом, если возбуждается двумя или больше синапсами (также светло-серого цвета). Таким образом, входное воздействие А]А 2 вызывает разряд Г 3 Г 4 (верхняя диаграмма), в то время как вход от А 3 А 4 достигает выхода на Г1Г2 (нижняя диаграмма). Нейроны Бг, Б 3 , Вг, В 3 активизируются в зоне пересечения этих двух входных воздействий. Серьезным недостатком этой диаграммы является то, что она игнорирует тормозные элементы (Eccles, 1968).
Конечно, передача волнового фронта в коре происходит гораздо сложнее. При вовлечении 100 нейронов на каждой релейной станции передняя волна может захватывать более 100 000 нейронов в одну секунду. Такая волна обладает… богатыми потенциальными возможностями…» (Eccles, 4958, р. 4-7).
Другой вывод следует из модели не распространяющейся стоячей волны, основанной на идеализированном представлении об анатомическом распределении связей между нейронами (Scholl, 1956). Все предположения, лежащие в основе этой модели, вполне разумны: 1) нейроны распределены в случайном порядке; 2) богатство связей между клетками уменьшается с расстоянием;. 3) пространственная структура распределения процессов в каждой клетке отличается тем, каким образом она передает возбуждение другим клеткам; 4) наблюдается затухание возбуждения во времени, то есть изменение потенциала в местах соединений имеет конечную длительность, и, наконец, 5) возбуждение само себя поддерживает.
Последнее предположение при отсутствии информации о тормозных взаимодействиях говорит о том, что любое состояние равновесия будет неустойчивым и что «вскоре обнаружится, что-активность в одних местах уже прекратилась, а в других возникла. По этой причине всегда предпочтительнее активность,, имеющая определенную пространственную и временную организацию…» (Beurle, 1956).
Детальные исследования электрической, а также анатомической структуры некоторых наиболее организованных мозговых образований в целом подтверждают обоснованность этих заключений.
«Лучше всего вто можно понять, если представить себе, что кора мозжечка… непрерывно подвергается воздействиям вследствие возникновения микроволн; каждая волна – это небольшой гребень активации длиной 3 мм , она возникает от клетки Пуркинье, которая имеет на другом конце тормозной сток. Эти волны не распространяются, но, конкурируя и интерферируя друг с другом, они, конечно, сильно модифицируют структуру волновых форм; более того, такая интерференция ведет даже к укорочению-волны, длительность которой становится менее 100 мсек. Эта операция конкурирующего взаимодействия волновых структур должна быть ключом к разгадке действия нейронного механизма…» (Eccles, Ito and SzentagoLhai,. 1967, p. 342).
Действительно, запись электрической активности, полученная от множества электродов, которая одновременно может быть представлена в пространстве (на топоскопе), говорит о возникновении какой-то волновой активности (Walter and Shiptom, 1951;. Lilly, 1949; Ливанов и Ананьев, 1955; Rernond, 1961). В каком отношении эта волновая форма соответствует нераспространяющейся стоячей волне, о которой говорил Берл (Beurle), остается еще исследовать. Важно в данном случае то, что описание нейронной активности медленных потенциалов как механической волны может помочь представить себе природу микроструктуры медленного потенциала или – более формально – послужить моделью, которая поможет дать более точную интерпретацию данных. Таким образом, подобный подход, состоящий в уподоблении медленного потенциала механической волне, хотя и не является необходимым для описания микроструктуры медленного потенциала, может в данном случае оказаться полезным (см. гл. VIII, рис. 1-16).

Рис. 1-16. Диаграмма, показывающая зависимость ЭЭГ космонавта Ф. В. от высоты подъема в камере, имитировавшей полет «Джеминай». Соотношение было исследовано на протяжении 70-минутного периода времени, в течение которого было сделано 40 записей. Каждая из них длительностью 20 мин. Обратите внимание на характеристику волнового фронта диаграммы и сравните ее с диаграммой на рис. 1-6 (Adey, 1967).
В заключение скажем, что наиболее распространенные представления о функции мозга основаны на том, что нервные импульсы возникают в нейронах и по ним передаются. Хотя точные нейрофизиологические исследования иногда предостерегают против чрезмерного упрощения, эти привычные представления, как ни странно, игнорируют активность соединений за исключением тех случаев, когда она имеет отношение к передаче нервных импульсов. Согласно этим взглядам, первичная задача синапса (или дендрита) состоит в передаче (или генерации) импульсов.
В противоположность этому Дж. Бишоп (1956) в заключительном обзоре, посвященном «действительной истории нервного импульса», утверждает, что «главные и наиболее характерные функции нейронов и других тканей, способных к возбуждению, проявляются посредством градуальных ответов». Он считает, что медленные градуальные потенциалы являются «более общим и более примитивным ответом, чем ответ «все или ничего», и что последний, вероятно, возник, когда древний многоклеточный организм стал слишком большим…». Бишоп анализирует довод в пользу утверждения, что кора головного мозга «все еще в большой степени функционирует посредством связей, характерных для примитивного неиропиля, что является самым подходящим; механизмом для поддержания непрерывного или устойчивого состояния, в отличие от передачи информации о таких состояниях». Вероятно, дендриты скорее, чем «проводящий импульс аксон»„ являются важнейшими элементами ткани с градуальным ответом. Подход, принятый в данной работе, согласуется с точкой зрения Бишопа. Активность соединений обладает эффективным способом действия, в котором доминируют механизмы непрерывного-нарастания и убывания медленных градуальных потенциалов. Благодаря этому возникает богатая и часто игнорируемая возможность понимания состояния как части двухпроцессного механизма, характеризующего мозговые функции. Это состояние вовсе не носит общего, глобального характера. Скорее, наоборот, оно само имеет микроструктуру, состоящую из медленных потенциалов соединений. Следовательно, нет нужды рассматривать нейрофизиологию всех психологических процессов исключительно в терминах операций проведения нервных импульсов. Признание Межпроцессного механизма деятельности мозга открывает перед исследователями более широкие возможности, эта концепция может быть очень плодотворной.
РЕЗЮМЕ
Классической единицей анализа функции мозга был нейрон. Признание двухпроцессного механизма вводит дополнительную-единицу: нейронное соединение, активность которого может становиться частью организации (микроструктуры медленных потенциалов), иногда не связанной с рецептивным полем одиночного нейрона. Таким образом, нейронных соединений- значительно» больше, чем станций на пути передачи нервных импульсов.
Глава II
ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ И МЕХАНИЗМЫ ПАМЯТИ
ПОИСКИ ЭНГРАММЫ
Одной из важных особенностей состояний мозга является их пластичность. В модификации мозговых состояний выражается самое основное свойство нервной системы, ее способность образовывать временные связи. Во второй главе будет более детально рассмотрена ткань мозга, в которой формируется микроструктура медленных потенциалов, с точки зрения ее пластичности.
До недавнего прошлого, несмотря на значительные усилия исследователей, еще не было прямых доказательств нейронной модификации, возникающей в результате индивидуального опыта организма. В 1950 г . Лешли закончил свою известную работу, посвященную исследованию энграммы, следующими словами: «Анализируя данные, касающиеся локализации следов памяти, я испытываю иногда необходимость сделать вывод, что научение вообще невозможно. Тем не менее, несмотря на такой довод против него, научение иногда происходит» (Lashley, 1950, р. 501).
Сейчас положение изменилось. Интенсивное изучение того, каким образом изменяется мозг под влиянием индивидуального опыта, привело к открытию нескольких направлений в исследовании проблемы. Мы рассмотрим данную проблему в три этапа. Настоящая глава будет посвящена развитию долговременных изменений в нервной системе в процессе научения. В главе III будет описан механизм средней длительности, который дает возможность настраивать нейронные цепи таким образом, что они приобретают возможность отвечать на последующие воздействия не так, как они отвечали раньше. Глава IV посвящена проблеме временной фиксации следов, которая делает возможным существование организующих операций.
Почему проблема биологии памяти, поиски доказательств того, что опыт вызывает постоянные изменения в нервной ткани, оказались столь трудными? Когда в каких-либо органах тела возни-(Кают со временем нормальные структурные изменения, мы говорим о росте и развитии ткани. Но в мозгу число нейронов не увеличивается после начального периода, непосредственно следующего за рождением. Таким образом, практически все развитие поведения и научение происходит при отсутствии каких-либо указаний на рост и развитие в нервной системе (рис. П-1).
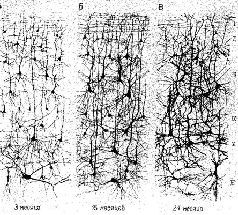
Рис. II-1. Схемы срезов коры головного мозга (импрегнированные по Гольджи) детей в возрасте 3, 15 и 24 месяцев. Обратите внимание на увеличивающееся ветвление и утолщение дендритов. До недавнего времени попытки установить корреляцию такого роста с жизненным опытом терпели неудачу (Conel (A), 1947; (Б), 1955; (В), 1959).
Этот парадокс может быть разрешен тремя направлениями исследований. Первое отрицает важность того факта, что нейроны не способны к воспроизведению, и делает попытку установить, что существует какая-то другая форма роста нервной ткани в зависимости от индивидуального опыта. Второе обращается к изучению нейроглии, ненейронных элементов нервной ткани, число которых может увеличиваться на протяжении жизни индивида. Наконец, мощным стимулом для объяснения этого парадокса послужили данные о том, что важные изменения связаны с химическими процессами хранения информации.
РОСТ НЕРВНОЙ ТКАНИ
Возможность роста нервной ткани не отрицается полностью тем фактом, что нейроны не способны к воспроизведению, как другие клетки тела. Нейроны отличаются друг от друга своими длинными волокнами, протяженностью разветвлений на теле клетки и теми амебовидными структурами на концах, названными конусами роста, которые можно наблюдать в культуре ткани и даже in situ (рис. П-2).
Рис II-2 Конус роста в действии Фрагменты фильма с замедленной съемкой. Обратите внимание на заметные изменения формы от фрагмента к фрагменту (Pomerat, 1964)
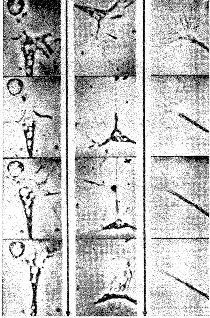
Конус роста толкает и давит на находящуюся перед ним ткань и может открывать путь, по которому он затем сможет медленно течь, удлиняя конец нервного волокна. Таким образом, рост может происходить при условии наличия свободного пространства, когда же его нет, амебовидный конец сокращается только для того, чтобы снова и снова продвигаться вперед. В центральной нервной системе рост обычно исключен потому, что элементы ткани расположены очень близко друг к другу. Правда, возникли большие сомнения в этом, так как результаты электронной микроскопии показали, что внеклеточное пространство, которое было найдено во всех частях организма, существует и вокруг нейронов.
Поэтому исследователи мозга стали наблюдать за ростом нервного волокна, создавая в мозгу свободное пространство. До недавнего времени такие попытки не приводили к значительным результатам: обычная реакция мозговой ткани на инсульт состоит в разрушении поврежденных частей и разжижении их, в образовании полости или кисты, оболочки которых препятствуют любой попытке проникновения в них. Однако современная техника дала исследователям инструмент, позволяющий избежать этой обычной реакции. Циклотрон излучает радиацию, которая резко замедляет процессы, происходящие в мягкой ткани. Таким образом, описанные разрушения можно произвести локально и только на том участке, на который направляется разрушающее воздействие. С помощью этого метода можно разрушать определенные слои коры взрослого кролика без видимого повреждения соседних. При этом методе повреждение ткани достаточно локально, так что обычно наблюдаемая реакция на большие разрушения нервной ткани не имеет места. Таким образом, создается пространство для активного роста нервных волокон (рис. И-3, П-4, П-5).

Рис П-3 Разрушение (I) в пятом слое постцентральной области коры кролика на 132-й день после воздействия пиковой дозы радиации в 48 000 рад Более тщательное исследование позволяет обнаружить многочисленные дендриты, достигающие области разрушения. Окрашено тионином (ХЗО) (Rose et al, 1961)
Рис. II-4. Область, заключенная в рамки на рис. И-3, показана при увеличении в 200 раз. Обратите внимание на апикальные дендриты входящие в область разрушения (Rose et a)., 1961).
Рис. II-5. Разрастание аксонов в области разрушения в стриарной коре крысы на 19-й день после облучения альфа частицами в 9000 рад. Окрашено по Водиану (Х390) (Kruger, 1965).
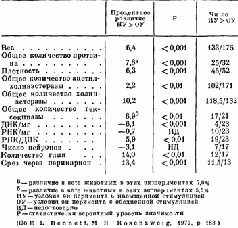
Таблица II-l
Влияние различного жизненного опыта на затылочную кору крыс о возрасте от 25 до 105 дней, содержащихся в насыщенных или обедненных стимулами условиях
Срезы, сделанные спустя несколько недель или месяцев после такого повреждения, показывают, что сохранившиеся волокна становятся толще (Kruger, 1965). Это утолщение, по-видимому, является совершенно нормальным увеличением размера волокна. Так как диаметр волокна часто является индикатором его длины, то утолщение означает в этих случаях, что мог иметь место и рост нервного волокна. Более прямым доказательством роста является появление большого числа нормальных, четко ориентированных волокон, которые не были видны ни до, пи сразу же после воздействия циклотрона (Rose, Malis, Baker, 1961). Следовательно, рост волокон возможен и в зрелом мозгу, если для этого есть благоприятные условия.
Эти эксперименты, конечно, не могут показать, является ли рост действительно результатом жизненного опыта; для этого должен быть проведен другой тип исследований. Чтобы проверить это, условия содержания крыс менялись таким образом, что одни из животных получали много стимулирующих воздействий (игра, решение задач), тогда как другие подвергались сравнительно ограниченной стимуляции. Иногда это ограничение касалось либо одной, либо другой сенсорной модальности (например, крысы содержались в полной темноте), чтобы проверить возможность различного воздействия на некоторые области мозга. Сравнение мозга животных, содержащихся в различных условиях, показало, что стимуляция вызывает заметное утолщение соответствующих отделов коры у животных, жизненный опыт которых был богаче (Krech, Rosenzweig, Bennett and Diamond, 1964).
Тщательный гистологический анализ утолщенной коры показал – как и следовало ожидать, так как число нейронов не увеличивается после рождения, – что количество нервных клеток на единицу объема, по существу, несколько уменьшилось. Однако было отмечено увеличение числа разветвлений базальных дендритов, увеличение числа и расширение дендритных шипиков – маленьких волосковых выступов, которые, как предполагают, являются местом активного соединения нейронов, распространяющих свои волокна горизонтально по коре. Большая же часть утолщения происходит за счет увеличения не нервных клеток – глии (рис. II-6, II-7).
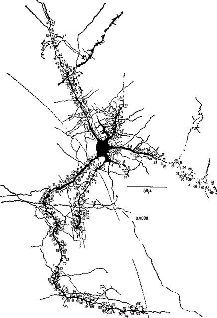
Рис. II-6 Сильно развитая дендритная система нейрона, имеющая 121 шипик, каждый из которых может действовать как точка соединения с другими дендритами (Calvin, см. Valverde, 1967).
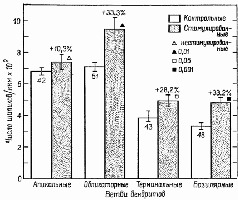
Рис II-7. Влияние стимуляции окружающей среды на развитие шипиков у 4 различных типов дендритных разветвлений пирамидных клеток коры Животные подвергались воздействию среды, насыщенной стимулами («стимулированные»), или среды, содержащей малое число раздражителей (контрольные). Числа внутри столбиков обозначают число нейронов, использованных при изучении каждого типа дендритных шипиков, которые определялись на различных участках ветвления дендритов Животные в возрасте 8 дней. В целом график показывает среднее число шипиков на микрометр для всех 4 типов дендритных ветвлений Все величины, характеризующие дендритные ветвления, включают стандартные ошибки и получены для 5 контрольных и 5 «стимулированных» животных (Schapiro and Vukovich, 1970).
РОЛЬ НЕЙРОГЛИИ
Вторым центром внимания при изучении проблем сохранения следов памяти стали имеющиеся в нервной ткани не нервные клетки, названные нейроглией (или просто глией). Глиальные клетки происходят из той же эмбриональной ткани, что и нервные. Долгое время считалось, что назначение глии состоит исключительно в том, чтобы служить источником питания для нейронов. Эта функция глии была достаточно хорошо доказана (например, Hyden, 1965). Глия идеально расположена для выполнения этой роли – некоторые из ее клеток (астроциты) окружают кровеносные сосуды, и таким образом посредством кровообращения может происходить метаболический обмен. Другие (олигодендрог-лия), имеющие небольшое количество ветвей, плотно оплетают нервные волокна, особенно на концах, заполняя таким образом межклеточное пространство вокруг нейронов на этих участках. Кроме метаболической функции, олигодендроглия способствует образованию изолирующего материала (миелина), .который покрывает крупные нервные волокна (рис, II-8).
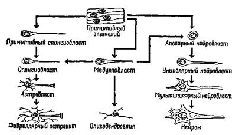
Рис. II-8. Развитие нейронов и нейроглии (Bailey, 1933).
Другое важное назначение олигодендроглии состоит, по-видимому, в том, что она направляет конусы роста нейронов. В периферической нервной системе известны клетки, тесно Связанные с глией и названные шванновскими, которые направляют рост нервных волокон, регенерирующих после повреждения. При этом имеет место такая последовательность событий: отсеченное или поврежденное нервное волокно погибает, дегенерирует по направлению к телу клетки, от которой оно происходит. Тело клетки может быть локализовано в спинном мозгу на расстоянии нескольких футов ( 1 фут = 304,8 мм ) от места повреждения. Немедленно начинает размножаться особый тип репаративных клеток, имеющихся в оболочке нерва, которые и заполняют пространство, освободившееся после гибели волокна. На фазе дегенерации они создают колонку ткани. Когда на теле клетки начинается процесс регенерации, путь, по которому будет следовать растущий конец нервного волокна, готов. Когда же такая колонка отсутствует, конус роста нервов распространяется во все направления и образует угол, который, конечно, не имеет функционального значения и часто болезненно чувствителен.
Эти специфические репаративные шванновские клетки в эмбриогенезе происходят из того же источника, что и нервная и гли-альная ткани. Более того, эти шванновские клетки обвиваются вокруг нервных волокон (рис. II-9) и способствуют образованию миелиновой оболочки подобно тому, как это делает олигодендрог-лия в центральной нервной системе.
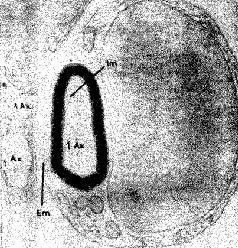
Рис. II-9. Фотография поперечного среза аксона периферического нерва (Ах), полученная с помощью электронного микроскопа и показывающая оболочку аксона, образованную шванновской (Em., Im.) клеткой (Truex, Carpenter, 1969).
Эти данные и привели большинство исследователей к объединению олигодендроглии и шванновских клеток в один класс. Вполне возможно, следовательно, что олигодендроглия направляет рост нейронов в центральной нервной системе, подобно тому как это делают шванновские клетки в периферической нервной системе.
Тот факт, что глия обвивается вокруг окончаний нервных волокон, ставит перед нами интересную проблему. За исключением тех случаев, когда соединение между нервами осуществляется посредством особых структур, называемых синапсами, изменения электрических потенциалов, возникающие в нейронах, вызывают потенциалы и в глиальных клетках.
Насколько же сильно влияние этих клеток на конфигурации, образуемые потенциалами соединений?
Хотя на такие вопросы еще не получено ответа, они заставляют предположить, что происходящие в глии изменения могут постоянно менять активность групп нервных клеток, с которыми она так тесно связана.
ХИМИЧЕСКАЯ ПЛАСТИЧНОСТЬ
Ряд экспериментов прямо указывает на участие глии в механизме памяти. Одним из веществ, обеспечивающим взаимодействие между глией и нейронами, оказалась рибонуклеиновая кислота (РНК – химическое вещество, определяющее конфигурацию-протеинов), которая сама является производной дезоксирибонуклеиновой кислоты (ДНК), молекулы генетической памяти. Фактически эти исследования показали, что при стимуляции нейронов вырабатывается больше РНК, чем в любой другой ткани тела (Hyden, 1961). После прекращения стимуляции и уменьшения выработки РНК в нейроне большие концентрации молеку-лярно сходной РНК начинают появляться в находящейся по соседству глии (Hyden, 1969).
Эти эксперименты, проведенные на мозге кроликов и крыс, были усовершенствованы введением выполняемой под микроскопом процедуры отделения глии от нейронов в вестибулярном ядре – группе нейронов, контролирующей поддержание равновесия животного. Стимуляция этого механизма «равновесия» первоначально осуществлялась следующим образом: животное помещалось в центрифугу и вращалось в ней. В более поздних экспериментах от крыс требовалось, чтобы они карабкались по наклонно натянутой проволоке. При успехе они достигали платформы с едой, при неудаче падали на расположенную внизу клетки решетку, по которой пропускался слабый электрический ток. На различных стадиях эксперимента микрохимическими методами определялась концентрация РНК в вестибулярных нейронах и глии животных. Во время пассивного вращения в центрифуге общее количество РНК в нейронах увеличивалось; увеличение содержания РНК было обнаружено и в течение нескольких часов после вращения. Активное карабканье по проволоке не только увеличивало общее количество РНК, но и изменяло соотношение фракций РНК, которые могли быть идентифицированы по боковым цепочкам, характерных для этих молекул (рис. II-10).

Рис. II-10. Установка эксперимента с крысой, взбирающейся по проволоке, чтобы достать пищу (Нуden, 1965).
По-видимому, никто не сомневается в том, что при физиологической стимуляции нервов или во время выполнения какого-то задания начинается выработка РНК. Однако дело обстоит не так уж просто. Спустя 24-48 часов после прекращения стимуляции изменение концентрации РНК и ее структуры более не отмечается ни в нейронах, ни в глии. Было высказано предположение, что РНК играет роль посредника между ДНК и протеинами, которые создают основу для более постоянной записи возбуждений. Тем не менее определение РНК полезно как указание на активный характер процесса памяти.
Этот взгляд на функцию РНК предполагает, что сохранение следа происходит в других макромолекулах, таких, как полипептиды, протеины, липопротеины и мукоиды (см. Bogoch, 1968; Glassman, 1967). Все эти молекулы имеют сложное строение и синтезируются в результате многих метаболических реакций. Следовательно, можно провести эксперименты, в которых тормозящие обмен вещества действовали бы на то или другое звено цепи синтеза. Эти вещества можно вводить как до и во время, так и после тренировки крыс. С помощью этой техники было обнаружено, например, что вещество, которое препятствует синтезу протеинов, может нарушать процесс припоминания, если лнъекция сделана спустя 5 или 6 часов после того, как крыса приобрела опыт в выполнении задания. Это означает, что эти вещества вызвали блокаду синтеза протеинов. Однако позднее было показано, что, если сохранение следов проверяется через несколько недель после инъекции, припоминание задания ухудшается лишь незначительно. Инъекция этих веществ, по-видимому, нарушает не конструкцию молекулы памяти, а, скорее, препятствует процессу обращения к ней (Agranoff, Davis and Brink, 1965). Однако роль протеинов и других макромолекул (особенно ли-пидов мозга) в механизме памяти, возможно, не столь ограниченна, как это предполагается гипотезой, проверяемой экспериментами с введением веществ", тормозящих обмен. В силу своей сложности макромолекулы имеют уникальные конфигурации, -структуры, которые могут меняться на короткое время и вызывать другое состояние (рис. II-11).
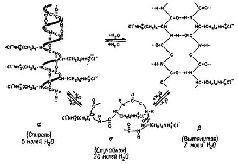
Рис. II-11. Схема возможных конфигураций молекулярных структур, которые может принимать молекула нуклеиновой кислоты (полимер poly-L-lysine hydrochlorid) (Blout, 1967).
С помощью структурных изменений можно хорошо объяснить временную память, связанную с такими образами, которые характерны для лиц с «фотографической» (или «эйдетической») памятью. Один такой эйдетически одаренный студент недавно был обнаружен в Гарварде. В результате экспериментальной проверки было показано, что он способен сохранять в памяти каждую деталь своих зрительных восприятий в течение 8 дней. Вторая часть книги будет в значительной мере посвящена рассмотрению этой временной спирализации молекул. Изучение структурных изменений мозговой ткани сейчас едва ли возможно, однако техника развивается такими темпами, что эта область исследования многое обещает уже в ближайшем будущем (Sjostrand, 1969).
Некоторые исследователи продолжают придерживаться гипотезы, что РНК является непосредственным хранилищем памяти, несмотря на серьезные доводы против этой гипотезы. Они основывают свою точку зрения яа весьма спорных экспериментах с «переносом», которые были проведены на планариях, крысах и обезьянах. В этих экспериментах экстракт РНК от контрольных животных и животных, обучавшихся выполнению задания, вводили необученным животным. Те из них, которым вводили РНК от животных, имевших опыт обучения, иногда, но не всегда решали задачи быстрее по сравнению с теми, которым вводили РНК от контрольных животных. Согласно некоторым данным, эффект различного действия РНК «обученного» и «контрольного» животного исчезал в результате обработки экстракта РНК веществом, избирательно разрушающим РНК (см. обзор McGonnell, 1970).
Сторонники химической гипотезы сохранения следов, оказавшись перед необходимостью выбора между двумя возможными решениями, в конце концов остановились на нейронных медиаторах. Уменьшение скорости и амплитуды нервных импульсов, когда они достигают окончаний аксонов, возникает из-за заметного уменьшения диаметра окончаний волокон. Следовательно, пресинаптических потенциалов самих по себе еще далеко не достаточно, чтобы вызвать постсинаптический потенциал. Однако пресинаптического электрического заряда достаточно, чтобы дать-толчок высвобождению медиатора, который хранится на окончании аксона в маленьких пузырьках (рис. II-12).
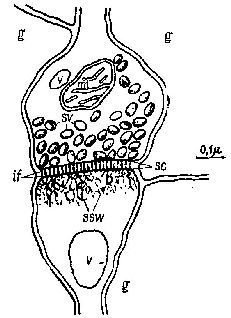
Рис. II-12. Типичная клетка коры. Обычная форма синапса-в мозгу млекопитающих. Ак-сонная (пресинаптическая) сторона вверху; дендритная (постсинаптическая) сторона – внизу, g – рлия; if – внутрисинаптические волоконца; mi – митохондрия, sc – синаптическая щь; ssw – субсинаптическая паутина; sv – синаптические пузырьки; v – везикулярное тело (Calvin. 1967, см . Roberts, 1S62).
Память могла бы зависеть от легкости, с которой высвобождается медиатор, или от его количества. Таким образом, можно провести такие эксперименты – и они действительно проводятся, – когда медиатор нейтрализуется в результате действия фармакологических веществ или его разрушение блокируется во время тренировки животного (Deutsch, Hamburg and Dahl, 1966). В результате таких действий у крыс нарушается процесс научения, но, как и в исследованиях с применением веществ, тормозящих обмен, возникает вопрос, не вызывает ли введение фармакологических веществ побочных эффектов, меняющих деятельность мозга и, следовательно, влияющих на процесс запоминания, – таких побочных эффектов, как появление локальных электрических разрядов в частях мозга, не связанных непосредственно с сохранением следов памяти, как таковой, но препятствующих обращению к ней во время проверки результатов научения.
ИНДУКЦИЯ КАК МОДЕЛЬ ХРАНЕНИЯ СЛЕДОВ ПАМЯТИ
Моя реакция на изобилие экспериментальных данных, полученных при исследовании энграммы в 1960-е гг., двойственна. Во-первых, можно прийти к выводу, что существует не один механизм памяти, не единая молекула памяти. Пластичность нейрона многообразна, и память не является единым процессом. Воображение, узнавание, припоминание предполагают использование некоторых или всех основных форм пластичности нервной ткани и различных способов сохранения и воспроизведения следа. Поэтому первый вывод, который следует сделать, состоит в том, что память, как она обычно представляется, не единый механизм, лежащий в основе процессов, которые позволяют организму устанавливать связи между отдельными звеньями опыта.
Во-вторых, простая модель следов, предполагающая постоянную модификацию мозговой ткани на нейронном уровне, может быть сформулирована уже сейчас. Такая модель должна обеспечить не только возможность сохранения следов, но и доступность считывания зафиксированного изменения. Наконец, такая модель должна быть основана на уже имеющихся данных и отвечать здравому биологическому смыслу. В настоящее время наиболее вероятным источником такой модели может служить аналогия с процессом, происходящим во время эмбрионального развития организма (рис. II-13).

Рис. II-13. Связь в виде треугольника, сформированная между тремя спинальными ганглиями эмбриона in vitro. Рисунок иллюстрирует тенденцию нервной ткани формировать узоры нервных волокон (Weiss, 1967).
Такие структуры, как глаз, формируются только тогда, когда ткань, из которой они развиваются, соответствующим образом стимулируется. Большая часть эмбриональной ткани эквипотенциальна, то есть ДНК во всех клетках тела, по существу, одна и та же. Это означает, что потенциал первоначально подавлен, находится в состоянии «репрессии». «Дерепрес-сия», или освобождение потенциальных возможностей, происходит тогда, когда появляются соответствующие условия. Определение эмбриологами того, каковы эти условия, заняло почти целое столетие. В настоящее время хорошо известно, что РНК и некоторые эндокринные секреты являются индукторами такой «дерепрессии». Маловероятно, что эти химические соединения несут в себе всю информацию, необходимую для индукции; большая доля специфики процесса должна быть отнесена за счет субстрата. Например, РНК, выделенная из печени теленка, может быть использована как индуктор для хрусталика глаза крысы.Отношения между индуктором и субстратом, по-видимому, следующие.
1. Индукторы извлекают и реализуют потенциальную генетическую информацию организма
2. Индукторы в известной мере специфичны в отношении характера извлекаемой генетической информации, но они неспецифичны относительно вида особи или ткани
3. Индукторы определяют общую схему индуцированного свойства, специализация деталей возникает в результате деятельности субстрата
4. Индукторы непосредственно не являются толчком для развития, они составляют особый класс стимулов
5. Чтобы быть эффективными, индукторы должны взаимодействовать с субстратами Однако, чтобы вызвать эффект, недостаточно простого контакта – ткань должна быть готова правильно реагировать
6 Индукция обычно развивается в результате действия в двух направлениях благодаря химическому взаимодействию между индуктором и субстратом.
Данные о роли РНК в сохранении следов памяти в настоящее лремя могут быть объяснены при помощи модели, основанной на этом эмбриональном процессе индукции. Модель предполагает, что возбуждение нервных структур сопровождается образованием РНК. Эта нейронная РНК вызывает изменения в окружающей «лигодендроглии, в результате чего начинается химический процесс между нейроном и глией, характеризующийся реципрокными отношениями изменений нейронной и глиальной концентрации РНК (и множества метаболитов). Происходит изменение в функциональном взаимодействии между глией и нейроном. На первом этапе может возникнуть соответствующее изменение концентрации РНК в глиальной клетке, которое затем в течение длительного времени будет вызывать изменение в структурах липидов, протеинов и липопротеинов во всех больших молекулах, составляющих мембраны, через которые осуществляется взаимодействие нейрона с глией. Такие макромолекулярные изменения могут влиять на легкость освобождения и разрушения нейронных медиаторов. Эти структурные изменения обратимы и могут стираться или быть вытеснены другими изменениями. Однако, если при неоднократном повторении одного и того же типа воздействия' изменения в молекулярной структуре будут продолжаться достаточно долго, они вызовут эффективные изменения мембранной проницаемости, которые в свою очередь способствуют появлению в возбужденном нейроне большого количества РНК, метаболитов и медиаторов, оказывающих влияние на окружающую его глию в том месте, где происходит деление глиальной клетки. Конус роста нейрона, освободившийся от инкапсулированной глии, свободно воздействует на недавно сформировавшиеся дочерние клетки в образует новые связи с нейронами. Таким образом, клеточное деление олигодендроглии направляет конусы роста нейронов центральной нервной системы во многом так же, как на периферии их рост направляют соответствующие шванновские клетки (рис. II-14).
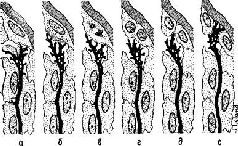
Рис. II-14 Шесть фаз гипотетического процесса индукции
КОНСОЛИДАЦИЯ СЛЕДОВ
Любое изменение глиально-нейронной организации следует рассматривать как изменение в микроструктуре активности медленных потенциалов соединений. Как мы уже говорили, такие изменения могут быть временными либо вести к возникновению более устойчивых изменений в нервной системе, связанных с ростом нейронов. Имеется много данных, свидетельствующих о том,, что для фиксации в мозгу следов памяти – энграмм – требуется время. Эти данные вполне согласуются с индукционной моделью хранения следов памяти, потому что они указывают на возможность выделения по крайней мере двух – если не больше – процессов в зависимости от времени их появления.
После достаточно сильной травмы головы человек не способен воспроизвести в памяти события, происходившие в период, непосредственно предшествовавший травме. Длительность этого периода зависит от тяжести травмы. Этот феномен, названный ретроградной амнезией, может быть использован для изучения фиксации следов, то есть для изучения процессов консолидации.
Самым распространенным методом, чтобы вызвать ретроградную амнезию в лабораторных условиях, является применение электроконвульсивного шока на крысах, хотя используются также и другие методы, такие, как удар по голове, быстрая анестезия с помощью эфира или введение определенных веществ, вызывающих амнезию. Полученные данные говорят о том, что, чем скорее после того, как возник навык, вызываются конвульсии, тем сильнее их тормозное, амнестическое действие на последние из навыков, приобретенные в этом эксперименте. Так, в одном исследовании (Chorover and Schiller, 1965) максимальный эффект отмечался в тех случаях, когда конвульсии следовали не позднее, чем через 15 сек после условной реакции избегания. Эффект практически исчезал, когда проверку выработки условного рефлекса и конвульсии разделял интервал длительностью 1 час. Эффект в значительной мере зависит от сложности задания, которое нужно было выполнить.
Однако это еще не все. При соответствующих условиях возникает более сложный, двухэтапный процесс консолидации. Допустим, испытуемый должен нажимать на рычаг, чтобы получить в качестве вознаграждения пищу, предъявляемую со слегка меняющимися интервалами. В какой-то момент выполнения задачи включается сигнал, за которым через определенный промежуток времени постоянно следует удар электрического тока в ногу. Как правило, при сигнале двигательный ответ испытуемого значительно уменьшается или совсем исчезает. Было высказано предположение, что этот эффект вызывается возникновением определенного эмоционального состояния. После действия тока и выключения сигнала восстанавливается нормальная скорость ответа. Когда подобный электрошок давался через 48 часов после последнего опыта и испытание возобновилось через четыре дня после завершения серий с электрораздражением, все испытуемые, подвергавшиеся его действию, были совершенно не способны «эмоционально» реагировать на сигнал. С другой стороны, если повторный опыт отодвигался на 30, 60 или 90 дней после завершения серий с электрораздражением, снова был получен условный эмоциональный ответ. В этих экспериментах (Brady, 1951) превышался критический интервал, в течение которого может быть максимально нарушен процесс консолидации, однако их результаты можно интерпретировать и как указание на то, что в течение одного-двух дней после эксперимента оказывается ослабленным какой-то механизм, необходимый для воспроизведения следа, на основе которого возникает эмоциональная реакция.
Результаты этих экспериментов по консолидации следов памяти можно интерпретировать, в соответствии с индукционной моделью, следующим образом: сразу же после испытания появление шоковых конвульсий, анестезия и т. д. препятствуют метаболическому обмену, происходящему между нейроном и олигодендрог-лией, тормозя индукцию, вызываемую путем секреции РНК в нерве и сопровождающуюся ее активацией в глии. Следовательно, образование энграммы прекращается и припоминание становится невозможным. Напротив, когда конвульсии возникают позднее, связанная с РНК фаза процесса индукции оказывается уже завершенной. В этом случае временно приостанавливается деление глиальной клетки, амебовидные движения окончаний нейронов временно прекращаются и конус роста втягивается назад. Однако при восстановлении этих механизмов процесс запечатления энг-раммы продолжается с того момента, когда он был прерван, и при завершении этого процесса становится возможным припоминание.
Чтобы твердо установить, что консолидация следа памяти действительно происходит, необходимо найти такой метод, который улучшал бы научение. Это, в сущности, нетрудно. Когда крысам вводили сульфат стрихнина или аналогичные возбуждающие вещества не ранее чем за 10 мин до начала или 30 сек спустя после пробы на прохождение лабиринта или на выработку дифференцировки, то навык у них вырабатывался быстрее, нем у контрольных; консолидация, таким образом, происходила скорее (McGaugh and Petrinovich, 1959).
Эти эксперименты служат красноречивым свидетельством того, что консолидация является необходимым условием возникновения следа памяти. Консолидация возникает в мозгу, но что же именно происходит во время этого процесса, который делает возможным запоминание? Ответ на этот вопрос могут дать следующие наблюдения.
В моей лаборатории на различные области коры мозга обезьян наносилась алюминиевая паста для создания локальных раздражений, которые в отдельных случаях вызывали эпилепсию. Эти раздражения проявлялись в измененной электрической активности – в возникновении патологических медленных волн и спайковых разрядов. Хотя такие нарушения не мешали обезьянам помнить решение задач, с которыми они неоднократно имели дело до раздражения, эти нарушения замедляли первоначальный процесс выработки данного навыка почти в пять раз (Pribram, 1966). Более того, способность вырабатывать такие навыки в целом не подвергалась нарушению; оно касалось только тех задач, выполнение которых было связано с определенной частью мозга, и ограничивалось лишь начальным периодом процесса выработки навыков, предшествующим появлению первых признаков научения. Таким образом, вызываемые в этом опыте раздражения не всегда блокируют консолидацию следов.
Может быть, одиночная энграмма, ограниченная лишь одним нейронным фокусом, является недостаточно эффективной, чтобы проявить себя? Большинство результатов экспериментов по научению у человека показывает, что, для того чтобы запомнить задание, необходимо его многократное повторение (Trabasso and Bower, 1968).
Во время повторения происходит такое распределение повторяемого материала, что он включается в целую систему связей предварительно зафиксированного опыта. Таким образом, результаты экспериментов с нарушениями, вызванными раздражением, можно объяснить тем, что процесс повторного возникновения и распределения энграммы был задержан. Проверку этой интерпретации можно осуществить путем сравнения результатов научения обезьян с повреждениями мозга в условиях распределенного концентрированного научения.
Гистологический анализ мозговой ткани, подвергшейся воздействию алюминиевой пасты, показывает наличие спутанных клубочков нервных волокон, во многом напоминающих соответствующие образования в периферических нервах, когда их рост не направляется соответствующей колонкой шванновских клеток. Возможно ли, чтобы это воздействие избирательно уничтожало олигодендроглию, вызывая ненаправленный рост нервных волокон? Химический анализ ткани, в которую была введена алюминиевая паста, конечно, не осуществим. Однако, чтобы получить ответ на этот вопрос, был проведен остроумный эксперимент (Morrell, 1961, рис. II-15).

Рис. II-15. Спонтанные разряды во вторичном эпилептогенном очаге активности. Вторичный очаг активности (два верхних канала) продолжает разряжаться, несмотря на прекращение разрядов в области первичного разрушения (нижние два канала). Калибровка: 100 мкв и 1 сек (Morrell, 1967).
Разрушение, вызванное раздражением в одном полушарии головного мозга, через несколько месяцев вызывает в контралатеральной коре появление «зеркального очага» измененной электрической активности, который возникает при участии мозолистого тела. Этот «зеркальный очаг» непосредственно не испытывал разрушающего химического воздействия, но обладал всеми признаками, характерными для первичного очага разрушения, вызванного раздражением. Существенно ли меняется РНК в «зеркальном очаге» по сравнению с нормальной тканью мозга? Может ли измененная РНК быть непосредственной причиной этого раздражения и последующих трудностей в восстановлении следа памяти?
Индукция в памяти, так же как и индукция в эмбриогенезе, является, видимо, многоэтапным процессом, который требует времени для своего завершения. Можно ожидать, что каждая стадия в таком процессе имеет свое уязвимое место, которое можно выявить с помощью соответствующего метода, примененного в критический момент. Уже многое известно о «критических периодах» в развитии эмбрионов и поведения в первые годы жизни. Определение природы критических, то есть сензитивных, периодов в процессе образования следов памяти представляет собой богатую область для исследования и экспериментов.
Можно еще многое сказать – и это будет сделано – о консолидации и модели индукции, но сначала нам нужно составить более полное представление о принципах работы мозга и о структуре психологических процессов. Поэтому рассмотрим в следующей главе некоторые из способов, посредством которых жизненный опыт организма может вносить свои изменения в мозг.
РЕЗЮМЕ
Длительно сохраняющиеся изменения происходят в соединительных аппаратах мозговой ткани. Хотя зрелые нервные клетки не делятся, механизм постоянной модификации мозговой ткани обладает многими свойствами механизма дифференциации эмбриональной ткани. В обоих случаях можно наблюдать экспериментально вызванный направленный рост новых нервныхволокон, который меняет пространственную структуру связей между нейронами. Следовательно, долговременная память является скорее функцией соединительных структур, чем функцией процессов в самой нервной клетке, генерирующей нервные импульсы.
Глава III
ЗАТУХАНИЕ АКТИВНОСТИ В НЕРВНОЙ СИСТЕМЕ И ТОРМОЗНЫЕ ВЗАИМОДЕЙСТВИЯ
ОРИЕНТИРОВОЧНАЯ РЕАКЦИЯ И ПРИВЫКАНИЕ
В первой главе были проанализированы два основных класса переменных, характеризующих деятельность мозга. Взятый в качестве модели процесс воспроизведения речи показал возможности, открывающиеся благодаря модификации этих двух переменных. Во второй главе были детально рассмотрены данные, свидетельствующие о пластичности мозга и его способности к постоянным изменениям; однако мы еще не говорили о свойствах самого механизма, который делает возможным осуществление таких изменений. При игре на скрипке, флейте или органе участвуют те же самые основные переменные, что и в речевом общении, хотя сами инструменты различаются тем, как они воспроизводят звуки.
Поэтому настоящая глава начинается с рассмотрения организации нервной системы, позволяющей ей быть аппаратом, создающим все богатство нашего опыта и поведения. Эта организация во многом зависит от относительно постоянных структур, образующихся в тех системах, посредством которых мозг приходит в соприкосновение со своим окружением. Благодаря своей доступности эти системы могут служить некоторыми частными моделями и, следовательно, подвести к исследованию более центральных процессов, анализ которых тебует иногда больших усилий. Поскольку довольно сложная организация имеет место уже при первоначальном контакте организма с его окружением, уместно начать наше исследование именно с этого процесса.
Экспериментальные данные говорят о том, что в любой момент времени происходит отбор сенсорного возбуждения с помощью хранящейся в памяти информации о прошлом опыте. Этот процесс сравнения, в ходе которого выявляется, соответствует ли текущее возбуждение хранящимся в мозгу записям, направляет как внимание человека, так и его действия.
Е. Н. Соколов (1960) провел следующий простой опыт, в котором обнаружилось одно из тех противоречий, которые порождают полезные парадоксы и указывают направление экспериментальным исследованиям в различных разделах нейропсихологии. Человеку предъявляется звук гудка. Сначала он вызывает ориентировочный рефлекс. Электрическая активность мозга характеризуется следующими чертами – активацией в латеральных отделах больших полушарий головного мозга (в виде увеличения доли низкоамплитудных и высокочастотных компонентов ЭЭГ) и гиперсинхронизацией в медиальных и базальных структурах мозга, на что указывает возрастание ритмичности и рост амплитуды волн среднечастотного диапазона. Кроме того, возникают другие локазатели ориентировочного рефлекса: увеличивается приток крови к голове за счет оттока от кончиков пальцев; меняется электрическое сопротивление кожи, частота сердцебиения и дыхания. Однако при многократном повторении звука гудка все эти реакции заметно уменьшаются. В таком случае говорят, что испытуемый обнаружил привыкание к стимулу.
Однако исчезновение реакции на повторяющиеся звуки оказывается обманчивым. В действительности многие из компонентов этой реакции еще сохраняются, так как, если стимул слегка меняется (например, звук становится тише), все первоначально исчезнувшие реакции восстанавливаются. Конечно, это парадокс. До открытия этого явления Соколовым высказывалось предположение, что привыкание просто повышает порог нервной системы на входе. Результаты, полученные Соколовым, означают, что лицо, которое обнаружило привыкание, должно было сопоставить действующий звук с сохранившимся представлением о ранее воспринятых звуках – иначе почему бы снижение интенсивности звука снова вызвало полноценную ориентировочную реакцию? Соколов проверил свою интерпретацию, многократно предъявляя испытуемому тон определенной длительности. Затем он неожиданно предъявлял более короткий звуковой раздражитель. Сразу же после прекращения звука возникали ориентировочные реакции; таким образом, изменение ответов происходило при наступлении тишины. Эти реакции продолжались в течение всей «ожидаемой» длительности звука, а затем медленно исчезали. Мы все испытывали эту удивительную реакцию на неожиданное наступление тишины (рис. III-1). Я назвал это «феноменом Бовери». В течение многих лет на Третьей авеню в Нью-Йорке существовала надземная железная дорога, которая производила ужасающий шум; когда она была снесена, люди, жившие в домах вдоль железной дороги, периодически просыпались от отсутствия звуков во время сна и звонили в полицию, чтобы сообщить, что происходит нечто странное, хотя они не могли точно определить, что именно. Эти звонки раздавались в те моменты, когда раньше с грохотом проносились поезда. Появление чего-то странного было, конечно, связано с наступлением «внезапной тишины», которая заменила ожидаемый шум.
Рис. III-1 А – угашение ориентировочного рефлекса (ЭЭГ и КГР) на раздражитель интенсивностью, условно обозначенной как 160; Б – растормаживание ориентировочного рефлекса при снижении интенсивности раздражителя до 120; В – то же самое, что и А; Г – возникновение ориентировочного рефлекса на уменьшение длительности раздражителя; Д – то же, что в А; Е – возникновение ориентировочных реакций на неожиданное увеличение длительности раздражителя. Следует отметить, что КГР также возникает с задержкой, что обусловлено замедленностью процессов в автономной нервной системе (Е. Н. Соколов, 1960).
Множество данных свидетельствует о том, что ориентировочные реакции и привыкание длятся сравнительно долго и являются в высшей степени специфическими. Каждая часть мозга характеризуется своими электрическими сигналами и своей последовательностью развития электрических явлений при осуществлении ориентировочной реакции. Если возникает привыкание, то нарушение его (ориентировочная реакция) при всяком изменении ситуации продолжает возникать на протяжении шести месяцев; изменения в последовательности событий, в обстановке, в которой они происходят, или в интенсивности, в длительности или конфигурации самих событий будут вызывать восстановление ориентировочных реакций.
Нейрофизиологии привыкания посвящено много исследований, и она вызвала много споров из-за различных экспериментальных подходов к решению этой проблемы. Для того чтобы стало возможным объяснение этого явления, необходимо уточнить некоторые факторы. Большинство нейрофизиологов называют привыканием любое постепенное ослабление ответа при повторном предъявлении того же самого стимула. Психологи же считают привыканием только те случаи, когда затухание ответа имеет место после нескольких проб и когда легко вызвать восстановление ориентировочной реакции. Недавно некоторые исследователи мозга, в том числе и я, приняли еще более строгий критерий Соколова: неожиданное уменьшение интенсивности, длительности стимула или его отсутствие должны вызывать восстановление ориентировочного рефлекса, если прежде этот рефлекс угасал в результате привыкания.
Важно также место электродов при электрофизиологической записи. Так, в моей лаборатории мы обнаружили, что привыкание действительно характеризуется затуханием ответа на любом уровне зрительной системы обезьяны от сетчатки до коры головного мозга. Однако при разном расположении электродов в коре одни из них дают признаки ослабления, другие – увеличения ответа, а третьи не обнаруживают никаких изменений. Следовательно, при повторении стимула в коре возникает изменение структур возбуждения; растормаживание разрушает эту структуру. Если бы мы смешали записи от всех кортикальных отведений или сделали бы записи от черепа, а не через электроды, вживленные непосредственно в кору, мы не смогли бы обнаружить этот эффект, о чем действительно сообщают некоторые исследователи.
Много работ было выполнено и на клеточном уровне. Была произведена запись от отдельных клеток спинного мозга и других отделов мозга. Во многих местах были обнаружены нервные клетки с затуханием ответа при повторении стимуляции. Такая реакция возникает даже у беспозвоночных, например у Aplesya Cali-fornica (Pinsker et al., 1970; Kupfermann et al., 1970; Castellucci et al., 1970). Однако можно ли у всех нейронов, у которых обнаружено привыкание, вызвать явления растормаживания? Недавно полученные Р. Томпсоном данные (Grove, Thompson, 1970) говорят о том, что можно различать по крайней мере три группы нейронов: нейроны, у которых реакция просто затухает; нейроны, реакция которых первоначально усиливается; и нейроны, характеризующиеся сначала усилением, а затем затуханием реакции. -Лишь у последней группы возможно растормаживание. Такие эксперименты до сих пор были проведены только на спинном мозге. Другие отчеты об экспериментах на структурах ствола мозга (верхнее двухолмие) свидетельствуют о том, что возможно, хотя и не обязательно, что одиночные нейроны с затухающими ответами в этой структуре могут обнаруживать растормаживание (Gerbrandt, Bures, Buresova, 1970).
Эти данные, а также другая работа Томпсона (Thompson and Spenser, 1966) показывают, что привыкание связано с взаимодействием нескольких нейронов. Томпсон использовал препарат, принесший известность Шеррингтону: животное, у которого спинной мозг отделен от головного. Повторная стимуляция электрическими залпами входных волокон спинного мозга приводит к постепенному уменьшению рефлекса сгибания конечности (вызванного подобно коленному рефлексу у больного, подвергающегося врачебному осмотру). Этот препарат создавал идеальные условия для изучения места и физиологической природы тех изменений в спинном мозге, которые связаны с привыканием (рис. III-2). Ни входные, ни выходные нейроны не участвуют в изменении рефлекса и, следовательно, не ответственны за привыкание: решающее значение имеют интернейроны, связывающие нейроны входа и выхода. Во-вторых, вопреки ожиданию введение фармакологических веществ, обычно используемых нейрофизиологами при изучении пре- и постсингштических .эффектов, не оказывало никакого влияния; эти вещества не приводили к затуханию ответа, свидетельствующему о привыкании. Это значит, что привыкание не связано с изменениями медленных потенциалов соединительных контактов. Сходные результаты получил Соколов (Соколов, Пакула и Аракелов, 1970). Он обнаружил, что медленные потенциалы мембраны нейрона и генераторные потенциалы, записанные от его аксона, которые дают начало нервным импульсам, могут изменяться независимо друг от друга. Кроме того, полученные им результаты свидетельствуют о том, что механизм генерации нервных импульсов может обнаружить привыкание.
Рис. III-2, Затухающий ответ эфферентной единицы на повторяющееся электрическое раздражение кожи (Thompson, 1967).
Таким образом, в некоторых случаях процессы затухания, лежащие в основе привыкания, по-видимому, связаны не с пре- и постсинаптическими изменениями, а с какими-то другими механизмами. Однако в других случаях были получены противоположные результаты. Иногда\по не всегда) этот процесс связан с уменьшением или увеличением притока медиаторных веществ,
С. Шарплесс (1967, 1969) показал, что при стимуляции нейронов у позвоночных не возникает ни сенситизации, ни десенси-тизации их сияаптических мембран. Напротив, Э. Кендел (Castel-lucci et al., 1970; Kupfermann et al., 1970; Pinsker et al., 1970) нашел, что привыкание и его нарушение у беспозвоночных локализуются в пресинаптических окончаниях и предполагают участие эффективного механизма секреции возбуждающих веществ в синапсах.
Стало быть, полученные данные не согласуются между собой. Необходимо найти ответ на целый ряд вопросов. Каким образом вообще механизм возникновения возбуждений может испытывать влияния, если не через посредство медленных потенциалов? Запускается ли внутри нейрона некоторый биохимический процесс и если да, то как (особенно если мы имеем дело с системой, которая обычно обнаруживает спонтанную активность и, следовательно, каким-то образом генерирует потенциалы)? Какова природа биохимических изменений и как они развиваются во времени? Соколов предполагает, что в этом механизме участвует РНК. Если он прав, то не связано ли привыкание с первыми этапами индукционного процесса или механизм привыкания совершенно независим от процесса постоянного хранения следов памяти? Возможно, что механизм постоянного хранения действует только при условии длительного и повторного привыкания. Если это так, то как это происходит? Наконец, каким образом изменение в структуре нервного импульса, вызываемого привыканием, действует на микроструктуру медленного потенциала на следующем синаптическом уровне? Чтобы ответить на некоторые из зтих вопросов, рассмотрим нейронную организацию сетчатки, этого маленького «кусочка мозга», который является гораздо более доступным для изучения, чем центральная нервная система.

