Глава 4
ЦЕНТРАЛЬНАЯ И ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Головной мозг
Самое высшее и наиболее совершенное достижение природы — головной мозг человека изучается сотни лет учеными всего мира, но до сих пор мы не можем сказать, что строение, химический состав и функции головного мозга полностью известны.
«Можно с правом сказать, — говорит И. П. Павлов, — что неудержимый со времен Галилея ход естествознания впервые заметно приостанавливается перед высшим отделом мозга, или, вообще говоря, перед органом сложнейших отношений животных к внешнему миру. И, казалось, что это недаром, что здесь действительно критический момент естествознания, так как мозг, который в высшей его формации — человеческом мозгу — создавал и создает естествознание, сам становится объектом этого естествознания».
Исследования русских физиологов — В. А. Бехтерева, И. П. Павлова и многих других позволили проникнуть в таинственную лабораторию высшей нервной деятельности.
Благодаря современным методам физического и химического исследования, усилителям электрических токов, микроскопам и электронным микроскопам, спектрографам и другим сложным приборам, изобретенным гением человека, удалось разрешить некоторые спорные вопросы физиологии центральной нервной системы. Этот таинственный орган, перед которым останавливалась мысль крупнейших исследователей на всем протяжении истории человечества, постепенно стал открывать перед нами свои загадочные свойства. Шаг за шагом он становится доступным познанию во всех своих деталях и своеобразных особенностях.
Головной мозг составляет передний и наиболее важный отдел центральной нервной системы. Слой серого вещества покрывает полушария головного мозга, образуя его наиболее совершенную и сложную часть — кору. В толще головного мозга можно обнаружить скопления нервных клеток, образующих так называемые подкорковые центры — высшие, промежуточные и подчиненные, — деятельность которых связана с отдельными функциями нашего организма. Густая сеть нервных волокон, объединяющих и связывающих различные центры, пучки нервных путей, выходящих из клеток коры и входящих в нее, составляют ткань мозга, сложную и непревзойденную по своему строению, химическому составу и физиологическому назначению.
Головной мозг разделяется на пять отделов: передний мозг с полушариями большого мозга, промежуточный мозг, средний мозг, мозжечок, продолговатый мозг с Варолиевым мостом (рис. 13).
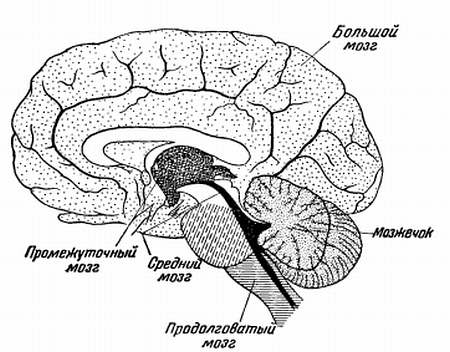
Рис. 13 . Отделы головного мозга
Большой мозг состоит из двух полушарий, на поверхности которых кора образует глубокие борозды и причудливые извилины. Каждое полушарие разделяется на отделы, называемые «долями» — лобной, теменной, затылочной и височной.
Кора больших полушарий связана нервными путями со всеми нижележащими отделами центральной нервной системы, а через них — со всеми органами тела. С одной стороны, импульсы, поступающие с периферии, доходят до той или иной точки коры, с другой, — кора посылает «распоряжения» в нижележащие отделы мозга, а оттуда к различным органам. Кора головного мозга осуществляет тончайшее равновесие между организмом и внешней средой, регулирует и направляет физиологические процессы, протекающие внутри организма, обеспечивает его сложнейшее функциональное единство.
Каждый анализатор (зрения, обоняния, слуха и т. д.) имеет, по представлению И. П. Павлова, в коре головного мозга центральную часть (ядро), где осуществляется высший анализ и синтез, и широкую периферическую зону, в которой аналитические и синтетические процессы совершаются в элементарном виде. Между ядрами отдельных анализаторов разбросаны и перемешаны нервные элементы, принадлежащие самым различным анализаторам. Если ядро анализатора в силу каких-либо причин разрушено или выбыло из строя, его функцию перенимают периферические элементы того же анализатора. Современная физиология отвергает и узкий («абсолютный») локализационизм и принцип однородности (равноценности) всех участков коры мозга.
На последнем Всесоюзном съезде невропатологов и психиатров в 1963 г. обсуждалась проблема локализации функций в коре головного мозга, и все же осталось много нерешенных и неясных вопросов, требующих уточнения и проверки. Нервные образования, которые мы привыкли называть «центрами», не ограничиваются корой головного мозга. Они включают и подкорковые структуры, значение которых недооценивается некоторыми исследователями. Работы советских физиологов, проведенные на обширном экспериментальном материале, подтвердили представление И. П. Павлова о «подвижной» («динамической») локализации функций в коре головного мозга. Кора головного мозга отличается необычайной пластичностью.
Одни отделы ее легко перенимают функции других, компенсируя расстройство их деятельности, вызванное различными причинами.
Наиболее важная задача современной науки выявить анатомическую основу физиологических процессов и одновременно установить связи и взаимосвязи между всеми явлениями, наблюдаемыми в головном мозгу. Исследования, проведенные русскими и иностранными учеными, показали, что в центральной извилине мозга, расположенной кпереди от центральной борозды, находится специальная двигательная область. Раздражение ее электрическим током вызывает сокращение определенных мышц противоположной стороны тела. Удаление этой области хирургическим путем ведет к расстройству координированных движений, шаткости походки, ослаблению мышц.
У человека ранение двигательной области сопровождается обычно параличами и другими тяжелыми нарушениями деятельности организма.
С помощью метода условных рефлексов удалось показать, что так называемые двигательные центры содержат чувствительные клетки, к которым приходят периферические раздражения от двигательного аппарата (костей, суставов, мышц). Эта область является мозговым концом двигательного анализатора в такой же степени, как затылочная область мозговым концом зрительного анализатора, височная — слухового анализатора и т. д. В двигательной области имеются чувствительные клетки, расположенные в верхних слоях коры, и двигательные, сосредоточенные в ее нижних слоях. Импульсы от рецепторов двигательного аппарата поступают в чувствительные клетки передней мозговой извилины, а отсюда уже передаются двигательным клеткам головного и спинного мозга. Таким образом, каждый двигательный акт, каждое так называемое произвольное, волевое движение детерминировано, т. е. обусловлено раздражениями, поступающими в кору головного мозга из внешней или внутренней среды.
Позади центральной борозды расположена чувствительная область коры. В ней заканчивается длинный путь, начавшийся в рецепторах кожи и внутренних органов. Здесь расположен его конечный этап. Каждое полушарие мозга связано в основном с противоположной половиной тела.
Однако существуют связи полушария и с одноименной половиной тела.
Еще не так давно считалось общепризнанным, что рецепторы внутренних органов не имеют своего представительства в коре головного мозга. Однако работы последних лет показали, что и интерорецепторы связаны с корой, хотя раздражение их не вызывает определенных ощущений и не доходит до сознания.
В настоящее время можно считать установленным, что процессы, протекающие в коре головного мозга, регулируют и направляют деятельность внутренних органов.
Каждый из личного опыта знает, что волнения, переживания, неожиданные известия, напряженная умственная деятельность сопровождаются характерными явлениями со стороны сердца, желудочно-кишечного тракта, мочевого пузыря и т. д. У лиц, страдающих грудной жабой или язвенной болезнью желудка, приступы болей возникают обычно при психических травмах, эмоциях, огорчениях и т. д.
Кора головного мозга является высшим распределителем деятельности всех внутренних органов. Раздражая электрическим током некоторые области коры, выключая их хирургическими или фармакологическими методами, исследователи установили, что можно по желанию изменять — ослаблять, усиливать, перестраивать — работу сердца, желудка, кишок, почек и других внутренних органов. Бесспорно, действие коры головного мозга на мочевой пузырь и прямую кишку, доказано ее влияние на слюно- и потоотделение, на состояние мышц, на просвет сосудов и т. д.
Условные рефлексы образуются не только при действии какого-либо внешнего агента, но и при всяком раздражении интерорецепторов. Условным раздражением может служить импульс со слизистой оболочки желудка, кишечника, мочевого пузыря, с рецепторов сосудов, селезенки и т. д. Любой сигнал из внутренних органов, если его сочетать с безусловным раздражением, может стать условным стимулом оборонительной, пищевой или какой-либо другой безусловной деятельности животного. Это говорит о том, что импульсы из внутренних органов обязательно проникают в кору головного мозга, так как только в этом случае возможна выработка на них условных рефлексов.
В верхней части задней центральной извилины мозга располагаются центры, воспринимающие ощущения из нижних конечностей, в нижней части — центры рецепторов кожной поверхности головы, лица и шеи. В коре затылочных долей обоих полушарий находятся зрительные центры, в височной доле — слуховые.
Если вырезать у собаки затылочные и височные доли больших полушарий, у нее начинают появляться некоторые отклонения от нормальной деятельности со стороны зрения и слуха. После удаления затылочных долей собака не теряет способности видеть. Она обходит встречающиеся на пути предметы, различает свет и темноту и в то же время не узнает хозяина. У нее разрушен мозговой конец зрительного анализатора, и это лишает ее возможности производить тонкий анализ зрительных раздражений. То же самое происходит и при разрушении мозгового конца звукового анализатора. Животное отличает только тишину от звука, но совершенно не в состоянии дифференцировать звуки, различать шумы и тона и т. д.
Иногда у людей наблюдаются заболевания, называемые «психической слепотой» или «психической глухотой». Такие люди хорошо видят предметы, но не в состоянии их узнать. Они хорошо слышат, но не понимают слов, не воспринимают речи и музыки. Эти заболевания возникают в тех случаях, когда целость коры головного мозга в затылочной или височной области нарушена. Нервные окончания у них в порядке, не пострадали и проводящие пути. Импульсы исправно передаются в головной мозг, но разрушено корковое ядро зрительного и слухового анализатора и, следовательно, расстроена аналитическая и синтетическая деятельность коры головного мозга.
Задняя центральная извилина воспринимает, по-видимому, также и чувство боли. Правда, вопрос этот нельзя считать окончательно решенным. Мнения исследователей разошлись, и если одним удавалось вызвать чувство боли при раздражении электрическим током задней центральной извилины, другие повторить этого опыта не сумели.
В 1883 г. В. М. Бехтерев, работая в клинике профессора И. П. Мержеевского, удалял у собак участки коры мозга в верхней части височной области и наблюдал значительное ослабление болевой чувствительности на противоположной половине тела. С тех пор накопилась большая, но весьма разноречивая литература по вопросу о локализации болевых центров в коре головного мозга. Хорошо известны опыты немецкого невролога Ферстера, который, раздражая кору головного мозга в области задней центральной извилины, не мог вызвать ощущения боли.
Однако, как было установлено более поздними исследованиями, раздражение верхней теменной дольки, особенно внутренней ее поверхности, вызывает обычно сильную боль. Некоторые исследователи наблюдали болевую реакцию при раздражении электрическим током отдельных участков коры головного мозга и мозжечка.
Интересные эксперименты были поставлены в Тбилиси профессором Д. М. Гедеванишвили. Он обнаружил участок коры в затылочной доле головного мозга, раздражение которого слабым электрическим током вызывало у кошек сильнейшую «реакцию гнева». Кошки становились необычайно агрессивными, начинали мяукать и кричать.
У них появлялись характерные для боли движения хвоста, приходили в движение брови и усы. Животные кричали, рвались из рук экспериментатора, выпускали когти и т. д.
Наблюдаемая картина, по мнению автора, воспроизводит болевую реакцию у кошки, вызванную раздражением периферического чувствительного нерва. Исходя из этого Д. М. Гедеванишвили пришел к выводу, что «реакция гнева» зависит от возбуждения специального центра болевого ощущения. Дальнейшие его опыты показали, что раздражение «центра боли» приводит к повышению кровяного давления, сердцебиению, одышке и т. п.
Многие экспериментаторы пытались вызвать болевое ощущение, раздражая электрическим током отдельные участки коры головного мозга (преимущественно области, расположенные кзади и кпереди от центральной извилины). Одни сообщали об удачных опытах, другие о неудачных.
По-видимому, при таком воздействии усиливаются только уже имеющиеся боли, особенно во внутренних органах, вызванные тем или другим болезненным процессом. Таково мнение видного американского ученого Суита.
Все эти опыты требуют проверки и уточнения. Вряд ли в коре головного мозга имеется строго локализованный «центр боли». Надо полагать, что наблюдаемая реакция вызывается не болью, а возбуждением корковых вегетативных центров. Большинство исследователей склоняется к мысли, что у человека болевая чувствительность связана с теменной долей головного мозга и задней центральной извилиной. Однако эмоциональную окраску чувство боли приобретает под влиянием лобных долей головного мозга. Одно время при лечении некоторых психических заболеваний производилась перерезка нервных путей, связывающих лобные доли с другими частями мозга. В этих случаях чувство боли не исчезало, но боль становилась безразличной, как бы далекой от реального мира.
В настоящее время широко применяется метод отведения электрических потенциалов от коры головного мозга. При помощи специальных усилителей удается записать на бумаге или фотопленке электрические токи, возникающие в различных отделах головного мозга. Нормальная активность мозга характеризуется определенным ритмом электрических колебаний и закономерной их величиной. При сильных болевых раздражениях электрическая активность коры мозга ослабевает, величина ритмичных колебаний (так называемых альфа-волн) уменьшается. Анализ полученных данных показывает, что при боли усиливаются процессы возбуждения по всей коре мозга и в то же время в ней появляются отдельные очаги застойного возбуждения. Эти изменения наиболее выражены по нисходящему ряду в теменных, затылочных, височных и лобных долях мозга. Однако они не специфичны для боли. Аналогичная картина была отмечена при сильных температурных воздействиях и даже при поглаживании кожи, прикосновении к ней и т. д.
Зрительные бугры
Помимо коры головного мозга, огромное значение для восприятия болевого ощущения имеют зрительные бугры.
«В физиологии нервной системы, — писал В. М. Бехтерев в 1883 г., — нет более темной области, как отправление зрительных бугров». Несмотря на то, что за истекшие годы появилось огромное количество работ, посвященных анатомии и физиологии зрительных бугров, роль этой области мозга далеко еще нельзя считать окончательно выясненной. Лишь в последние годы некоторые стороны деятельности зрительных бугров стали значительно яснее и стала более понятной их роль в физиологии больших полушарий мозга.
Зрительные бугры представляют собой большие скопления серого вещества в межуточном мозгу. С двух сторон, с правой и левой, они образуют стенки третьего желудочка мозга с огромным количеством самых разнообразных нервных путей, которые связаны почти со всеми отделами головного, продолговатого и спинного мозга.
На разрезе легко обнаружить, что в зрительных буграх имеется пять основных серых ядер, разделенных прослойками белого вещества. К передней части внутреннего (медиального) ядра подходят волокна обонятельного тракта, к задней его части — волокна протопатической (см. стр. 132) болевой чувствительности. Здесь по современным представлениям формируются чувства «удовольствия» и «неудовольствия». Задняя часть наружного (латерального) ядра является основным центром проприоцептивной чувствительности, т. е. получает импульсы из мышц и суставов, а также тактильные, болевые и температурные сигналы.
Остальные ядра не имеют непосредственного отношения к восприятию боли. Роль их многообразна и зависит в значительной «степени от связей с другими отделами мозга.
Бесконечное число нервных волокон соединяет ядра зрительных бугров с ядрами подбугорья, где заложены центры высших вегетативных центров (обмена веществ, терморегуляции, сосудистого тонуса и т. д.) с другими подкорковыми образованиями и, в первую очередь, с корой головного мозга. Чтобы убедиться в этом, достаточно взглянуть на рисунок 14.
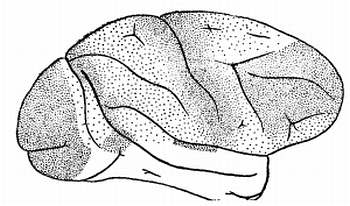
Рис. 14. Окончания бугрово-корковых путей в мозгу обезьяны макаки
Каждая точка, изображенная на поверхности коры головного мозга обезьяны макаки, показывает окончание пучка волокон, соединяющих зрительные бугры с корой. Эти волокна пронизывают всю толщу мозга. Они обеспечивают бесперебойную двустороннюю связь от зрительных бугров с корой и передают импульсы в обе стороны: от клеток коры к зрительным буграм и обратно.
В течение многих лет дебатировался вопрос о расположении высших центров болевого ощущения. До недавнего времени считалось наиболее вероятным, что болевое ощущение формируется не в коре головного мозга, а в зрительных буграх. Это предположение поддерживали многие виднейшие физиологи и невропатологи мира. Впервые его высказали английские ученые Гэд и Холмс, а в дальнейшем французы Лавастин и Лериш, русские исследователи Аствацатуров и Орбели, а за ними и многие другие. В то же время большинство исследователей склоняется к мысли, что эта область мозга является лишь важнейшим, узловым, но в то же время промежуточным центром болевого восприятия.
Зрительные бугры представляют древнейшую часть головного мозга. Они образовались в нервной системе живых существ, населявших землю за много миллионов лет до появления человека. Лишь в процессе эволюции, на более высоких ступенях зоологической лестницы появилась та часть мозга, которую мы называем полушариями.
Спор между сторонниками «корковой» и «бугровой» локализации болевого восприятия потерял в настоящее время свою остроту. Вскрылись новые обстоятельства, позволившие по-новому проследить путь болевых ощущений, совершенно иначе расценить давно известные факты и установившиеся, как казалось многим, «ученические» истины.
Уже давно известно, что при раздражении какого-нибудь чувствующего нерва в коре головного мозга можно обнаружить два вида электрической активности, различающихся временем их возникновения.
В 1942 г. американские ученые Морисон и Демпси показали, что и при электрическом раздражении ядер зрительного бугра в коре можно получить два типа электрических реакций. Первый с коротким скрытым периодом, обнаруживающимся преимущественно в чувствительных зонах коры, в корковых ядрах тех или иных анализаторов. Второй — с длинным скрытым периодом, распространяющимся по всей коре и не поддающимся точной локализации. Эта распространенная (генерализованная) реакция коры головного мозга наблюдается при раздражении всех видов чувствующих волокон — зрительных, слуховых, обонятельных, тактильных, температурных и болевых. Можно предположить, что периферические раздражения поступают в кору по двум различным проводящим системам. Один путь — специфический, знакомый нам во всех деталях. Другой — до недавнего времени гипотетический, мало изученный.
Ретикулярная формация
Используя специальные так называемые стереотаксические дриборы, физиологи научились вводить тончайшие, невидимые невооруженным глазом электроды не только в скопления нервных клеток, расположенные в глубинных отделах мозга, но даже в отдельные нейроны.
Электрический ток, пропущенный через такие электроды, способен раздражать нервные ядра мозга, а в случае необходимости разрушать строго ограниченные участки мозговой ткани. При этом экспериментатор имеет возможность наблюдать, каким становится поведение животного, как изменяются в его организме физиологические и биохимические процессы.
Хирурги во время операции на центральной нервной системе давно заметили одно интересное обстоятельство.
Оказалось, что можно во время операции производить любые разрезы через кору больших полушарий мозга, можно удалять значительные участки мозговой ткани, иногда даже целое полушарие, тем не менее пациент продолжает бодрствовать и не теряет сознания. Однако стоит лишь слегка задеть инструментом некоторые точечные участки в глубине мозга, как больной немедленно впадает в бессознательное состояние, похожее на сон. В конце 40-х годов нашего столетия два видных физиолога — американец Г. Мэгоун и итальянец Д. Моруцци — провели на животных серию экспериментов, которые не только объяснили это явление, но и заложили основу изучения функций особого образования мозга, известного под названием ретикулярной формации.
Ретикулярная формация представляет скопление нервных клеток в центральных отделах той части головного мозга, которую называют мозговым стволом. Нервные клетки ретикулярной формации, благодаря большому числу ветвящихся и переплетающихся отростков, образуют густую нервную сеть, откуда и возникло название «ретикулярная» или в переводе на русский язык сетевидная формация. Анатомы уже давно описали этот своеобразно построенный отдел мозга. Но значение его стало понятным лишь сравнительно недавно.
Если у животного разрушить определенные участки ретикулярной формации мозгового ствола, сохранив неповрежденными другие мозговые образования, наступает состояние длительной спячки. Вывести животное из спячки не удается, даже применяя самые сильные внешние раздражители. Сигналы об этих раздражениях беспрепятственно достигают коры больших полушарий головного мозга, но животное на них не реагирует. При этом электрические потенциалы, возникающие в коре мозга, ничем не отличаются от потенциалов, наблюдаемых при глубоком естественном сне.
Попробуем через специально вживленные электроды раздражать слабым электрическим током сохранившиеся после опыта участки ретикулярной формации. Животное сразу просыпается, а на электроэнцефалограмме появляются типичные сдвиги, названные физиологами «реакцией пробуждения».
Результаты этих опытов убедительно показывают, что ретикулярная формация непосредственно участвует в процессах регулирования сменяющих друг друга состояний — сна и бодрствования. В чем же смысл этого регулирования?
Как известно, мозг человека содержит полтора десятка миллиардов весьма сложно устроенных нервных клеток.
Довольно подробно изучены многочисленные, иногда в высшей степени причудливые, связи этих клеток друг с другом. Казалось бы активное состояние — возбуждение, возникнув в одной клетке, может беспрепятственно распространяться по всему мозгу. Если бы это на самом деле имело место, у нас не было бы никаких оснований восторгаться деятельностью мозга. Любое воздействие на организм приводило бы к одной и той же однотипной реакции, напоминающей судорожный припадок, сходный с эпилептическим. А между тем мы прекрасно узнаем, как точно приурочены ответы мозга к вызвавшему их воздействию. Достигается это благодаря очень точной и четкой системе регулирования физиологических процессов в мозгу. Одни его отделы способны воспринимать и перерабатывать приходящие к ним раздражения, а другие нет.
При возбуждении одних нервных клеток в других, иногда лежащих по соседству, иногда отдаленных, возникает торможение (подавление, угнетение).
Исследования последних лет показали, что в мозгу имеются специальные образования, предназначенные главным образом для регулирования деятельности его отдельных рабочих частей. Одним из наиболее изученных образований подобного рода и является ретикулярная формация.
Было доказано, что для нормальной деятельности головного мозга одного только поступления в кору чувствительных сигналов недостаточно. Необходимо еще определенное рабочее состояние, рабочая настройка клеток коры. Лишь в этом случае могут реализоваться необходимые процессы восприятия и перерабатываться поступившая в кору информация. Такое рабочее состояние элементов коры создает ретикулярная система. Внешне это состояние проявляется в виде бодрствования. Вот почему ретикулярная формация получила название активирующей системы мозга (рис. 15).
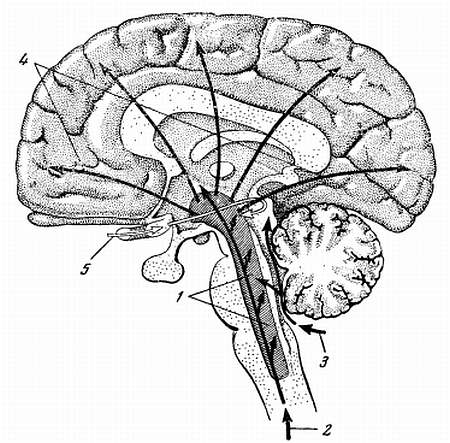
Рис. 15. Активирующие механизмы мозга
1 — ретикулярная формация, 2 — восходящие чувствительные пути спинного мозга, 3 — путь слуховых импульсов, 4 — путь активирующих импульсов к коре головного мозга, 5 — путь зрительных импульсов
Возникает вопрос, каким же образом ретикулярная формация способна длительно активировать кору больших полушарий, создавать в ней состояние бодрствования? Оказалось, что от всех чувствительных путей, направляющихся в головной мозг и передающих сигналы, вызванные светом, звуком, прикосновением, давлением, теплом, холодом, болью, отходят ответвления, заканчивающиеся у поверхности клеток ретикулярной формации. Любое внешнее раздражение — вспыхнувшая лампа, стук в дверь, рукопожатие, боль — возбуждает ретикулярную формацию. Она как бы сама «заряжается» энергией и заряжает кору головного мозга. Сигналы, воспринятые ею, теряют свою специфичность. Они становятся лишь инструментом «зарядки», «тонизирования» коры головного мозга.
Вначале все казалось очень простым и легко понятным. Ретикулярная формация, благодаря притоку соответствующих импульсов от органов чувств, создает рабочую «атмосферу» в клетках коры и тем самым поддерживает состояние бодрствования. Но на самом деле это только схема, далеко не исчерпывающая сложных взаимоотношений в центральной нервной системе.
Мы все хорошо знаем как трудно заснуть после дня, богатого событиями и переживаниями. Если же к этому еще примешиваются тревоги, волнения, заботы, может наступить бессонница, т. е. ненормальное, длительное, как бы неотключаемое состояние бодрствования. Человек не может заснуть, хотя приток раздражений извне даже уменьшен. Хорошо известно, что страдающие бессонницей пытаются выключить даже самые слабые и привычные раздражители. Они останавливают на ночь тикающие часы, занавешивают окна, укутывают голову одеялом.
Понять и объяснить эти явления удалось сравнительно недавно, когда было обнаружено, что, помимо нервных влияний, передающихся из ретикулярной формации «вверх», в коре больших полушарий существуют влияния, поступающие в обратном направлении — от коры к ретикулярной формации. Возбуждение, возникшее в определенных участках коры, оказывает в одних случаях стимулирующее, в других подавляющее влияние на ретикулярную формацию. Следовательно, сама кора головного мозга может и поддерживать, и подавлять бодрствование. Система кольцевых связей: кора — > ретикулярная формация — > кора дает возможность высшим отделам центральной нервной системы регулировать через ретикулярную формацию свое рабочее состояние. Этим, вероятно, объясняется то обстоятельство, что только некоторые сигналы «заряжают» ретикулярную формацию.
Не все поступающие из окружающего мира раздражения способны прервать наш сон. Спящая мать реагирует на едва заметное движение ребенка, но не просыпается от грохота проносящегося поезда. Многие люди пробуждаются по заданию в требуемое время, многие умеют засыпать в любых обстоятельствах, несмотря на свет, шум, разговоры. Кора головного мозга, этот высший распорядитель, распределитель, регулятор и организатор деятельности организма посылает, в зависимости от прошлого опыта человека пли животного, в ретикулярную формацию только выборочные, определенные сигналы.
Можно считать доказанным, что бодрствование поддерживается активирующим («заряжающим», «облегчающим») влиянием ретикулярной формации на клетки коры головного мозга. Угнетение, торможение клеток, ретикулярной формации, блокада ее активирующего влияния на кору лежит в основе различного торможения центральной нервной системы, т. е. сна. Вот почему перерыв или прекращение активирующих влияний, постоянно поступающих в кору бодрствующего мозга, вызывает сонливость, сон, потерю сознания. В то же время короткие электрические раздражения ретикулярной формации пробуждают спящее животное.
Бодрствование выключается и при некоторых болезненных состояниях организма, например при травме черепа. Это также связано с подавлением активирующих влияний ретикулярной формации на кору больших полушарий.
В лаборатории П. К. Анохина был поставлен такой опыт. Кролику наносили болевое раздражение, опуская заднюю лапу в горячую воду, нагретую до 60°. Было отмечено, что при этом в коре головного мозга возникают определенные изменения электрической активности. Если же кролику ввести предварительно аминазин, вещество блокирующее некоторые элементы ретикулярной формации, болевое ощущение не вызывает сдвигов в электроэнцефалограмме. Врачи знают, что аминазин облегчает чувство боли. Но самое интересное заключается в том, что аминазин блокирует не всю ретикулярную формацию, а только те ее элементы, которые возбуждаются под влиянием боли, страха и других отрицательных эмоций. При положительных эмоциональных реакциях, например при кормлении ретикулярная формация продолжает активировать клетки коры.
Из этого был сделан вывод, что ретикулярная формация, которую все исследователи привыкли считать «неспецифической», содержит определенные нервные элементы, влияющие не на всю кору, а только на отдельные комплексы клеток и, следовательно, обладает специфическим действием на те или другие функции.
В дальнейшем был выявлен еще один интересный факт.
Если у крысы разрушить определенную область ретикулярной формации, она начинает поглощать огромные количества пищи, никогда не насыщаясь и превращаясь постепенно в жировой ком.
Раздражение током других участков ретикулярной формации вызывает у кошек приступы ярости, а разрушение их превращает послушное, прирученное животное в дикое и агрессивное.
Возникает вопрос, какой же смысл в столь широко распространенном влиянии ретикулярной формации на самые различные проявления деятельности нервной системы, на функции практически всех частей организма?
Вернемся снова в физиологическую лабораторию и проделаем несложный опыт. Опять-таки через вживленные электроды пропустим слабый электрический ток в ретикулярную формацию бодрствующего животного. Действие тока скажется почти моментально. Животное «замирает», настораживается, чего-то ждет. Мышцы его напряжены, а на электроэнцефалограмме отмечаются изменения, характерные для высокой активности высших отделов головного мозга.
Такая же картина наблюдается у животного при новом и неожиданном внешнем раздражении, например свистке, вспышке света, окрике. Подобного рода реакцию И. П. Павлов назвал ориентировочным рефлексом, рефлексом «что? что такое?», рефлексом на новизну. Разрушение ретикулярной формации делает невозможным осуществление ориентировочного рефлекса.
Ретикулярную формацию можно рассматривать как устройство мозга, обеспечивающее состояние «общей мобилизации» организма по сигналу тревоги. Извне поступил новый и неожиданный сигнал. Что это: пища? опасность? друг? враг? Организм должен быть подготовлен к любой деятельности, которая потребует: схватить добычу, убежать, спрятаться, не обращать внимания, если тревога окажется ложной. Эта подготовка к любому виду деятельности выражается в переходе высших отделов мозга в более активное состояние, в изменении настройки двигательного аппарата, в обостренном восприятии действительности органами чувств (не поступят ли еще какие-нибудь сигналы, проясняющие обстановку?), в изменении дыхания, работы сердца и других внутренних органов.
Резервы подтянуты, организм пришел в состояние боевой готовности.
Но вот сигналы повторяются. Теперь уже необходимо произвести анализ обстановки, выработать план действий. Теперь уже мобилизация всех нервных приборов не нужна. Требуется лишь настройка тех механизмов нервной системы, которые принимают непосредственное участие в соответствующих неотложных мероприятиях. И высшие отделы головного мозга подавляют деятельность ретикулярной формации. Приказ об общей мобилизации отменен.
Ориентировочный рефлекс угасает…
Советские и зарубежные исследователи установили, что клетки ретикулярной формации возбуждаются не только физическими сигналами. Они чутко реагируют на меняющееся содержание в крови углекислоты, сахара, некоторых химических веществ, гормонов и, в первую очередь, гормона, вырабатываемого надпочечниками — адреналина.
Так, например, при голодании, когда «голодная» кровь омывает ретикулярную формацию, она мобилизует нервные механизмы поведения, направленные на поиски и добывание пищи. Как остроумно отметил один ученый, она «способствует переводу внутренних потребностей организма в поведение». Затрудненное дыхание у спящего человека приводит к накоплению углекислоты в крови, что вызывает возбуждение ретикулярной формации и пробуждение. Это спасает человека от удушения во сне.
Еще в начале нашего столетия И. П. Павлов, изучая образование условных рефлексов у животных, говорил об особом значении для психической деятельности подкорковых образований мозга, «заряжающих» энергией кору больших полушарий. Теперь уже ни у кого не возникает сомнений, что подкорка является своеобразным аккумулятором энергии и в ней находится особое регулирующее «зарядное» устройство — ретикулярная формация (рис. 16).
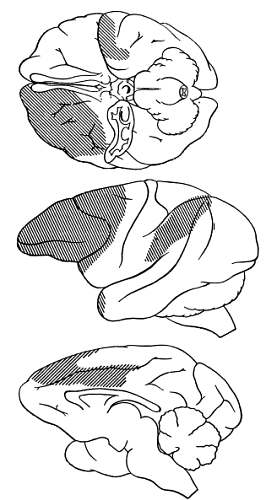
Рис. 16. Проекция активирующей системы мозгового ствола в коре больших полушарий головного мозга обезьяны макаки
Разумеется, она не представляет единственный регулирующий механизм мозга. Природа чрезвычайно изобретательна и предусмотрительна: в мозгу имеются и другие регулирующие аппараты. Любые попытки некоторых зарубежных ученых поставить ретикулярную формацию во главе всей деятельности мозга должны быть отвергнуты, как не соответствующие научной истине.
Изучение функций ретикулярной формации головного мозга явилось важным шагом вперед на пути познания конкретных материальных механизмов головного мозга.
Путь этот долгий, трудный и некоторую, правда небольшую, его часть мы прошли.
Какое же отношение имеет ретикулярная формация к восприятию боли?
Можно считать установленным, что при любом раздражении поверхности кожи поступающие с периферии импульсы заряжают ретикулярную формацию мозгового ствола.
Болевое раздражение, как и всякое другое, воспринимается клетками чувствительной зоны коры головного мозга. Но вслед за этим через какой-то короткий промежуток времени, исчисляемый тысячными долями секунд, наступает общее возбуждение всей коры, обусловленное активирующим влиянием ретикулярной формации.
По мнению австрийского исследователя Гоффа, высказанному еще в 1953 г., существуют два механизма возникновения чувства боли в головном мозгу, связанные у здорового человека воедино. При раздражении одного механизма (специфического) возникает простое ощущение боли, при раздражении второго механизма (неспецифического) — сложное, комплексное ощущение, обозначаемое как страдание, недуг, болезнь. Это ощущение осуществляется при участии целого ряда нервных образований: зрительных бугров, подбугровой области, ретикулярной формации и коры головного мозга. Благодаря первичному процессу возбуждения, развивающемуся в зрительных буграх, мобилизуются центры подбугровой области (обмена веществ, сосудодвигательные, потоотделения, секреции, эндокринной регуляции и т. д.), возникают разнообразные вегетативные реакции, столь характерные для боли. В то же время ретикулярная формация мобилизует всю кору для реакции на болевое раздражение, так что последняя оказывается в состоянии привести в действие многообразную систему мероприятий, необходимых для устранения источника боли или для компенсации вызываемых ею нарушений жизнедеятельности организма. Во время болевого раздражения ретикулярная формация посылает в кору головного мозга огромное число нервных сигналов. Такие же сигналы идут из зрительных бугров и подкорковой области. Это приводит к резкому изменению электрической активности коры.
Разрушение бугровой области мозга у человека, вызванное закупоркой питающей ее артерии или опухолью, ведет к своеобразным изменениям чувствительности, известным в клинике под названием таламического (бугрового) синдрома. Лица, страдающие этим синдромом, жалуются на болезненные ощущения в одних участках кожной поверхности и на отсутствие чувствительности в других. Нередко легкие раздражения кожи, прикосновение, толчок вызывают приступы мучительных болей. Больные почти постоянно испытывают чувство «ползанья мурашек», давления, покалывания и т. д. Возникнув, ощущение сохраняется значительно дольше, чем у нормального человека.
При достаточно сильном раздражении электрическим током ядер зрительных бугров у лиц, страдающих тяжелыми болями в различных частях тела, удастся вызвать характерные изменения электрической активности коры в области задней центральной извилины. При этом испытуемые ощущают сильнейшие боли именно в той области тела, где находится болевой очаг. После хирургического удаления соответствующего центра в зрительном бугре, электрическое раздражение не вызывает боли.
Зрительные бугры собирают множество отдельных раздражений, возникших в самых различных рецепторах нашего тела. Однако огромное количество разнообразных импульсов так и не доходит до нашего сознания. По-видимому, не все сверхсильные раздражения, способные вызвать чувство боли, воспринимаются сознанием. Возникновение боли говорит о том, что болевые импульсы достигли высшего отдела центральной нервной системы — коры больших полушарий головного мозга.
Если зрительные бугры являются в основном центром древней, грубой, ничем не смягченной (протопатической) чувствительности, то кора головного мозга способна дифференцировать сигналы тонкой чувствительности, призванной отрегулировать и локализовать чувство боли. Конечные центры эпикритической чувствительности находятся в коре (стр. 132).
Раздражение специфических нервных окончаний кожи, мышц, внутренних органов воспринимается как болевое ощущение именно корковыми клетками головного мозга. Кора больших полушарий играет основную роль в восприятии и «осознании» боли. Субъективное ощущение боли формируется именно в коре. Здесь объединяются и связываются в единое целое все раздражения, поступающие с периферии по разнообразным центростремительным путям. Частично эти пути заканчиваются в зрительных буграх, но благодаря наличию богатой сети корково-бугровых волокон, благодаря ретикулярной формации, так или иначе связываются с клетками высших отделов мозга.
Осознание и дифференцировка болевого ощущения происходят в высшем распорядительном отделе центральной нервной системы — в коре больших полушарий головного мозга — при участии «активирующих систем» мозгового ствола.
Вегетативная нервная система
Еще не так давно в медицине и биологии считалось установленным, что нервный аппарат человека и животных состоит из двух самостоятельных, почти независимых друг от друга систем — анимальной и вегетативной. Начало этому учению положил 150 лет назад французский врач и физиолог Мари Франсуа Биша. Все функции организма он разделил на анимальные, свойственные только животным, и вегетативные — общие как животным, так и растениям.
Ощущение, движение, речь составляют, по Биша, группу анимальных функций; питание, рост, размножение — группу вегетативных. Отсюда и нервная система, регулирующая функции животной жизни, получила название анимальной, а нервная система, регулирующая функции растительной жизни — вегетативной (рис. 17).
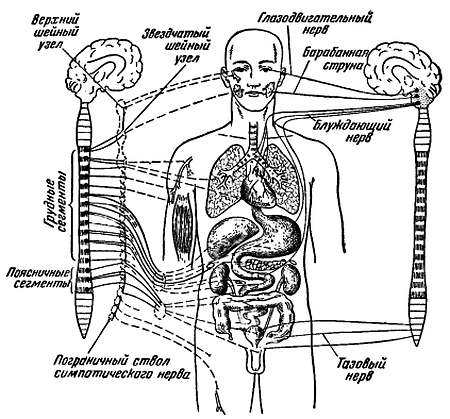
Рис. 17. Схема строения вегетативной нервной системы (по К. М. Быкову)
Вегетативная нервная система подразделяется на два больших отдела — симпатический и парасимпатический.
Большинство органов и тканей снабжено как симпатическими, так и парасимпатическими волокнами. Эта двойная иннервация имеет огромное значение для жизнедеятельности нашего организма. Она обеспечивает тонкую и точную регуляцию функций. Но в то время, как симпатические волокна проникают во все органы, парасимпатические разветвляются лишь в некоторых.
Влияние симпатических и парасимпатических нервов на физиологические процессы не одинаково, иногда даже противоположно. В одних случаях симпатический отдел: вегетативной нервной системы возбуждает деятельность, клеток, тканей, органов, а парасимпатический тормозит ее, в других — имеют место обратные соотношения.
Господствовавшее в XVIII в. представление о функции: симпатической нервной системы как о регуляторе деятельности органов, обеспечивающем «симпатии и согласие органов» (отсюда и название этой части нервной системы), давно уже имеет лишь историческое значение и заменено более точными анатомо-экспериментальными данными науки.
При возбуждении симпатических нервов учащается ритм сердечных сокращений, суживаются сосуды, повышается кровяное давление, пульс становится быстрым и напряженным, усиливается обмен веществ, улучшается питание тканей. В то же время гладкая мускулатура желудка и кишок расслабляется и венечные сосуды, снабжающие сердце кровью, расширяются. Симпатические нервы расширяют зрачок и сокращают мышцы волосяных мешочков, вследствие чего волосы взъерошиваются, как говорят, «становятся дыбом, ощетиниваются». Симпатические импульсы вызывают отделение густой, вязкой слюны и в то же время уменьшают секрецию желудочного сока. Под влиянием симпатических нервов утомленная, едва сокращающаяся мышца начинает работать энергичнее, бодрее.
В ней изменяются химические процессы, повышается обмен веществ, увеличивается образование тепла.
Многочисленные рецепторы нашего организма, в том числе кожные рецепторы, воспринимающие прикосновение, давление, боль, тепло и холод, иннервируются волокнами симпатической нервной системы. Она снабжает ими даже спинной мозг и головной мозг, в том числе его высшие отделы.
Несколько иную роль играет в организме парасимпатическая нервная система. Действие ее во многих отношениях противоположно действию симпатической. При возбуждении парасимпатических волокон ритм сердечных сокращений замедляется, сосуды расширяются, кровяное давление падает и одновременно суживаются венечные сосуды, что ведет к ухудшению кровоснабжения сердца. Импульсы, идущие по парасимпатическим нервам, вызывают отделение жидкой, обильной слюны, увеличение секреции желудочного и кишечного соков, усиление движений желудка и кишок, отложение резервов в организме. При возбуждении парасимпатических волокон зрачок резко суживается, увеличивается отделение слез, нарастает количество мочи.
Обычно принято считать, что в нормальном организме симпатическая нервная система способствует повышению общей активности организма, а парасимпатическая — восстановлению затрат, связанных с его деятельностью. Однако далеко не всегда симпатический и парасимпатический отделы нервной системы действуют противоположно.
Вряд ли в организме вообще возможен столь резко выраженный антагонизм. Понятие об антагонизме, т. е. о «противоборстве», «противодействии» между различными отделами вегетативной нервной системы уже давно подлежит пересмотру. Можно говорить об относительном антагонизме, вернее о единстве противоположностей, о едином механизме противоположно действующих факторов.
Между симпатическим и парасимпатическим отделами вегетативной нервной системы существуют тесные анатомические и физиологические связи. Они способствуют согласованной совместной деятельности и взаимодействию различных частей вегетативной системы. Но, по-видимому, нередко одни и те же процессы в организме возникают (или прекращаются) под влиянием симпатических нервов и прекращаются (либо возникают) под влиянием парасимпатических.
Вегетативная нервная система принимает самое деятельное участие в тончайшей игре механизмов, регулирующих физиологические процессы, возбуждающих одни клетки и тормозящих другие.
В течение многих лет вегетативную нервную систему рассматривали как автономное, независимое от головного и отчасти спинного мозга, нервное хозяйство, управляющее явлениями «растительной» жизни. К последним относили кровообращение, дыхание, пищеварение, мочеотделение, другими словами все непроизвольные процессы, не подчиняющиеся воле и сознанию.
Перелом наступил в начале XX в. Огромную роль сыграли в этом отношении работы русских и советских исследователей.
Оказалось, что «растительные» функции в такой же степени подчинены головному мозгу, как и «животные».
Об этом писал еще в 1875 г. выдающийся русский физиолог Василий Яковлевич Данилевский. Раздражая электрическим током лобные доли мозга, он вызывал изменение деятельности сердца, нарушение ритма дыхания. Интереснейшими работами казанской физиологической школы, возглавляемой Н. А. Миславским, было окончательно установлено, что в продолговатом мозгу находятся центры, регулирующие дыхание и кровообращение. За каких нибудь два-три десятилетия исследования современных физиологов полностью перестроили представление об автономности вегетативной нервной системы и показали, что она является неотъемлемой составной частью единого, целостного нервного аппарата.
Академик Л. А. Орбели создал и развил теорию адаптационно-трофической (т. е. приспособительной роли симпатического отдела вегетативной нервной системы в организме). Он считал, что последняя, изменяя обмен веществ, тем самым настраивает орган, приспособляет его к определенной деятельности. Такое влияние симпатическая нервная система оказывает на все органы и ткани, в том числе и на центральную нервную систему, включая ее высшие отделы.
Вегетативная нервная система в основном реализует импульсы, поступающие из высших отделов нервного аппарата. Ее волокна выходят из клеток, заложенных в центральной нервной системе, где находятся высшие и подчиненные вегетативные центры (рис. 17). Состояние организма, его реакция на внешние воздействия зависят как от анимальной, так и от вегетативной нервной системы.
Та часть нервной системы, которую мы называем вегетативной, необычайно чутко реагирует на все раздражения, поступающие из внешней и внутренней среды и обеспечивает приспособление организма к постоянно меняющимся внешним и отчасти внутренним условиям.
В то же время она находится под постоянным влиянием окружающей среды. Это влияние осуществляется корой головного мозга посредством сложной системы нервных и гуморальных регуляторов (см. стр. 104). Время дня, время года, атмосферное давление, погода, температура воздуха оказывают на вегетативную нервную систему свое закономерное влияние. Исключительное значение для реакций, осуществляемых вегетативной нервной системой, имеют внешние воздействия на органы чувств (свет, звук, запах).

Леон Абгарович Орбели
Нельзя делить единый нервный аппарат на независимые друг от друга отделы и участки, как и нельзя противопоставлять вегетативную нервную систему анимальной и наоборот.
В последние годы появились исследования, посвященные болевой чувствительности вегетативных узлов и волокон. Во время операций на людях было установлено, что шейная симпатическая цепочка чрезвычайно чувствительна к механическим и электрическим раздражениям. Прикосновение к ней скальпелем или пинцетом вызывает резкое, подчас мучительное болевое ощущение. Пациент отмечает боль в различных органах, иногда даже расположенных довольно далеко от места прикосновения инструмента. Так, например, раздражение верхнего шейного узла симпатической нервной системы вызывает сильные боли в зубах, в нижней челюсти, а иногда в ухе. Раздражение звездчатого узла сопровождается тяжелыми болями в плечевом поясе и в спине. Лериш утверждает, что распределение вегетативных волокон левого звездчатого узла полностью совпадает с распространением болей при грудной жабе. Он наблюдал возникновение сильнейших болей в области сердца при раздражении этого узла иглой или электрическим током.
Все это привело к мысли, что болевые раздражения могут передаваться в центральную нервную систему по симпатическим или даже парасимпатическим волокнам.
Хотя в литературе нет единства взглядов по этому вопросу, все же можно думать, что чувствительность внутренних органов связана с вегетативной нервной системой. Это тем более вероятно, что еще в 1896 г. выдающийся русский гистолог А. С. Догель описал в ней чувствительные нейроны. К сожалению, его работы были забыты, и лишь в недавнее время венгерский ученый Кисе подтвердил, что в симпатических цепочках существуют чувствительные волокна.
Для деятельности вегетативной нервной системы характерны особые виды рефлексов, так называемые аксон-рефлексы. В отличие от истинных рефлексов, аксон-рефлексы осуществляются без непосредственного участия нервных центров. Возбуждение, возникшее в периферическом рецепторе, не достигает нервной клетки, а переходит в точке разветвления нервного волокна с одной ветви на другую, вызывая таким образом тот или иной эффект.
Реакции подобного рода впервые наблюдал русский физиолог И. Соковнин и описал их еще в 1873 г., задолго до англичанина Ленгли, которому принадлежит название аксон-рефлекс. Однако надо полагать, что даже эти примитивные нервные реакции все же находятся под контролем центров головного и спинного мозга.
Существует немало химических соединений, усиливающих и ослабляющих деятельность вегетативной нервной системы. Симпатический отдел вегетативной нервной системы возбуждается при введении в кровь адреналина — гормона, вырабатываемого надпочечниками, железами внутренней секреции, расположенными в брюшной полости человека и животных. Достаточно ввести в кровь небольшое количество адреналина, чтобы усилить деятельность симпатических нервов и узлов. В обычных условиях адреналин постоянно поступает из надпочечников в кровь и стимулирует симпатическую нервную систему. Подобным же образом действуют и многие другие, так называемые «симпатикотропные», т. е. возбуждающие симпатический отдел вегетативной нервной системы, вещества. К ним относятся норадреналин, симпатол, синефрин, неоситаефрин (вазотон, мезатон), фенамин, эфедрин, тирамин, соли кальция и т. д.
Но попробуем ввести под кожу или в кровь незначительное количество другого вещества, широко распространенного в природе и имеющего огромное значение для жизни организма — ацетилхолина. Почти тотчас же после введения кровяное давление начинает снижаться, деятельность сердца резко замедляется, сосуды расширяются, зрачки суживаются. У человека усиливается потоотделение, отмечается тошнота. Это говорит о том, что ацетилхолин возбуждает парасимпатический отдел вегетативной нервной системы и в первую очередь волокна и окончания так называемого блуждающего нерва, который снабжает своими ветвями все внутренние органы. Почти такое же действие, как ацетилхолин, оказывают на организм и некоторые другие вещества, известные под названием «парасимпатотропных», например карбохолин, пилокарпин, ареколин, соли калия.
За последние годы накопилось много интересных фактов, показывающих, что деятельность вегетативной нервной системы в значительной степени зависит от состояния ретикулярной формации мозга.
Французские ученые — Делл и Бонвалле — установили, что адреналин и норадреналин усиливают тонус симпатической нервной системы, активируя определенные (адренергические) элементы ретикулярной формации. Некоторые отделы головного мозга (в особенности гипоталамическая область и ретикулярная формация) особенно богаты адреналином и норадреналином. По-видимому, они играют наиболее важную роль в осуществлении симпатических реакций. Некоторые отделы ретикулярной формации особо чувствительны к ацетилхолину. При их возбуждении усиливается деятельность парасимпатического отдела вегетативной нервной системы.
Вегетативная нервная система принимает самое широкое участие в осуществлении болевых реакций. Ее центральные и периферические отделы мобилизуются при болевых раздражениях и вызывают ряд сложных физиологических процессов, о которых будет идти речь в последующих главах.

