Книга: Кости, гены и культура
Назад: А у ос – наоборот!
Дальше: Глава 11 Социальное обучение и культурные традиции
Связь размера мозга и социальности у обезьян
Данные по нечеловеческим приматам в целом неплохо согласуются с идеей о том, что жизнь в больших, сложно организованных коллективах, где между особями складывается запутанная система личных отношений, была важным стимулом для развития мозга в эволюции обезьян, включая и предков человека.
В частности, широко известны пионерские работы британского антрополога Робина Данбара, который обнаружил у обезьян положительную корреляцию между характерным для данного вида размером социальной группы и относительным размером неокортекса. На этом основании Данбар даже пытался предсказать размеры групп вымерших гоминид – австралопитеков, хабилисов и так далее, опираясь на известные размеры их эндокранов (книга 2, глава 4).
Что показали дальнейшие исследования? Разумеется, то, что всегда показывают дальнейшие исследования: “все сложнее и не так однозначно”. Это настолько строгая закономерность, что даже скучно: если вы видите пионерскую работу с яркими результатами, будьте уверены – дальнейшие исследования обязательно покажут, что “все сложнее”.
Мы уже знаем, что у современных людей размер некоторых отделов мозга положительно коррелирует с общительностью, которую можно оценивать как по результатам специальных психологических тестов, так и просто по количеству друзей и знакомых (хотя этот вывод основан, как мы помним, на маленькой выборке и не может считаться окончательным, см. раздел “Коррелирует ли размер мозга с социальным интеллектом?” выше).
На обезьянах были получены похожие результаты, например, в исследовании, проведенном в 2011 году британскими нейробиологами на макаках-резусах (Sallet et al., 2011).
Теория социального мозга предсказывает, что жизнь в большом коллективе должна повышать нагрузку на определенные отделы коры. Следовательно, в этих отделах может возрасти объем серого вещества. Ведь объем серого вещества (он отражает размер тел нервных клеток), как и объем мышечной ткани, от постоянных тренировок может увеличиться. А может и не увеличиться – если теория надуманна.
Предположение проверили следующим образом. Взяли молодых (четырехлетних) обезьян и на год поместили их в новые коллективы разного размера (от 1 до 7 особей в группе). Размер группы, в которую попала та или иная обезьяна, не зависел ни от особенностей ее характера, ни от того, в какой группе она жила до начала эксперимента. А через год при помощи магнитно-резонансной томографии у этих уже пообвыкшихся в новом коллективе животных оценили объем серого вещества в разных отделах мозга.
Результаты совпали с теоретическими ожиданиями, основанными на теории социального мозга. У обезьян, живших в больших группах, объем серого вещества в нескольких отделах мозга к концу эксперимента оказался больше, чем у обезьян, живших в маленьких группах. Обратного эффекта, то есть меньшего объема серого вещества у обезьян из больших групп по сравнению с обезьянами из маленьких групп, ни в одном участке мозга замечено не было.
Наиболее сильная положительная корреляция между размером группы и объемом серого вещества характерна для нескольких участков височной доли неокортекса: середины верхней височной борозды, ростральной части верхней височной извилины, нижней височной извилины и височного полюса. Увеличение коллектива обезьян на одну особь (в пределах исследованного диапазона, то есть от одной особи до семи) приводит к увеличению объема серого вещества в этих отделах мозга в среднем на 5,42 %.
Перечисленные отделы мозга как у макак, так и у людей участвуют в восприятии лиц и жестов (верхняя височная борозда), в распознавании звуковых сигналов и хранении семантической памяти (верхняя височная извилина); повреждение височного полюса у макак ведет к нарушению эмоциональных реакций. Так что увеличение объема серого вещества в этих отделах, вероятно, отражает повышенную потребность в анализе мимики, жестов и звуковых сигналов многочисленных соплеменников.
У макак, живших в больших группах, увеличилась также амигдала – отдел мозга, играющий важную роль в эмоциональной регуляции поведения. У людей тоже выявлена положительная корреляция между размером круга общения (социальной сети, в которую вовлечен человек) и объемом амигдалы, однако для людей не удалось выяснить, где тут причина, а где следствие. Эксперимент на макаках показал, что причинно-следственная связь здесь имеется и что направлена она от размера коллектива к размеру амигдалы. Что, впрочем, не исключает существования связи, направленной в обратную сторону – от размера амигдалы к количеству особей, с которыми данный индивид взаимодействует.
Размер группы у подопытных макак положительно коррелировал также с объемом ростральной части префронтальной коры. У людей эта часть мозга (вместе с верхней височной бороздой и некоторыми другими участками) играет ключевую роль в оценке намерений других людей и прогнозировании их реакций, то есть в том, что называют “теорией ума” или “моделью психического состояния другой особи” (theory of mind). Скорее всего, у макак ростральная префронтальная кора выполняет сходные функции. Некоторые приматологи сомневаются, что у макак есть полноценная “теория ума”, но макаки, по крайней мере, могут предугадывать поступки соплеменников на основе того, что те видят или не видят. А это уже важный шаг на пути к приобретению данной способности. Например, известны эксперименты, когда обезьяны намеренно обманывали друг друга. Если одной обезьяне показать, где спрятана вкусная еда, скажем банан, то она в присутствии высокоранговой особи будет вести себя так, будто ни о каком банане не знает. И подойдет к тайнику только тогда, когда останется в одиночестве. Она своим поведением обманывает высокоранговую особь, предугадывая ее поведение, ведь та с большой вероятностью отберет припрятанное угощение. А если усложнить дело, спрятав огурец и банан (последний, очевидно, предпочтительнее), то обезьяна будет всем своим поведением, взглядами или движениями, подводить высокоранговую товарку к огурцу. Получается, что обманщица ведет себя в соответствии с построенной моделью поведения соплеменницы (Whiten, Byrne, 1988). Между прочим, данных по “тактическому обману” у разных видов приматов накоплено довольно много. Анализ этих данных еще в 2004 году показал, что частота случаев обмана у обезьян положительно коррелирует с объемом неокортекса (Byrne, Corp, 2004).
Логично предположить, что увеличенный объем серого вещества в участках мозга, связанных с социальными отношениями, способствует более результативному социальному поведению. У макак хорошей мерой этой результативности является социальный статус, или ранг, занимаемый особью в группе. Показано, что у самцов макак социальный ранг зависит от способности к формированию коалиций, которая, в свою очередь, определяется умением поддерживать товарищеские отношения с другими самцами. Ученые оценили социальный ранг 11 подопытных самцов (9 из которых жили в группах из 4 или 5 обезьян) по количеству побед в конфронтациях с соплеменниками. Оказалось, что общественное положение этих самцов положительно коррелировало с объемом серого вещества в ростральной префронтальной коре и нижней височной извилине. Таким образом, предсказания теории социального интеллекта снова подтвердились.
Исследователи также обнаружили, что у обезьян из больших коллективов повышена корреляция между уровнями активности верхней височной борозды и передней части поясной извилины. Последний отдел у макак, как и у людей, отвечает за оценку социально значимой информации, получаемой от соплеменников. Повышенная согласованность также отмечена в работе верхней височной борозды и тех участков мозга, из которых в нее приходит зрительная информация. Таким образом, с ростом коллектива не только увеличивается объем некоторых участков мозга, но и растет степень скоординированности их работы.
Исследование показало, что увеличение объема социально ориентированных отделов мозга может быть не только причиной (или базисом) участия индивида в сложной сети общественных взаимоотношений, но и следствием такого участия. Эти отделы можно натренировать подобно мышцам, а конечным результатом такой тренировки может стать рост общественного статуса индивида.
Все подобные исследования основаны на понимании того, что мозг позвоночных в целом имеет мозаичную (модульную) структуру: он состоит из множества довольно-таки специализированных отделов, различающихся как анатомически, так и функционально. Если в ходе эволюции общий объем мозга увеличивается или уменьшается, то пропорции его частей имеют обыкновение меняться предсказуемым образом (см. раздел “Опережающее развитие ассоциативных сетей” в главе 9). Это свидетельствует о неких онтогенетических или функциональных ограничениях, не позволяющих отделам мозга совсем уж произвольно менять свои пропорции. Однако на этом фоне все же существует значительная межвидовая (а также межродовая, межсемейственная и так далее) изменчивость по относительным размерам отделов мозга (Barton, Harvey, 2000).
Предполагается, что эта изменчивость во многом связана с адаптацией к различным условиям существования. Например, если мы видим, что у первых млекопитающих по сравнению с их предками цинодонтами (группа зверозубых ящеров) резко увеличились отделы мозга, связанные с обонянием и осязанием, то логично предположить, что это было напрямую связано с переходом к ночному образу жизни. Среди примеров такого рода – положительная корреляция между богатством песенного репертуара и относительным объемом вокального центра у певчих птиц (Devoogd et al., 1993) и увеличенный гиппокамп у пернатых, запасающих пищу впрок (Krebs, 1990). Также широко известно исследование, в котором изучались лондонские таксисты, вынужденные профессионально ориентироваться в одном из самых сложных для автомобилистов городов мира. Выяснилось, что у них заметно увеличена задняя часть гиппокампа по сравнению с людьми других профессий (Maguire et al., 2000), хотя в этом случае речь идет о пластических изменениях, то есть о результатах тренировки, а не эволюции.
В пределах отряда приматов изменчивость размера и пропорций мозга чрезвычайно высока – и столь же изменчивы экологические и социальные характеристики. Поэтому на приматах удобно смотреть, какие части мозга связаны с теми или иными экологическими и социальными адаптациями. В 2019 году американские приматологи сопоставили имеющиеся литературные данные по анатомии мозга приматов с несколькими социальными и экологическими факторами, а именно с диетой, ночным или дневным образом жизни и со сложностью социальной организации (DeCasien, Higham, 2019). Вид Homo sapiens был исключен из рассмотрения, поскольку он слишком резко отличается от остальных приматов как по строению мозга, так и по социоэкологическим особенностям.
В исследовании было учтено больше видов приматов и больше отделов мозга, чем в прежних публикациях на эту тему: 33 отдела мозга, для каждого из которых нашлись данные по 17–58 видам приматов. Учитывалось все, что удалось найти в литературе, так что информация для каждой комбинации “отдел мозга / вид” включала сведения по десяткам особей. Для внесения поправок на родство видов (чтобы минимизировать влияние так называемой филогенетической инерции) использовались новейшие уточненные эволюционные деревья.
Качество диеты оценивалось двумя альтернативными способами. В первом случае использовалось простейшее бинарное деление: всеядные и плодоядные виды относили к группе с “высококалорийной диетой”, а питание листьями считалось “низкокалорийной диетой”. Второй подход основан на вычислении индекса качества питания, который для приматов традиционно рассчитывают по формуле 1s + 2r + 3,5a, где s – доля структурных частей растений, r – доля репродуктивных частей растений, a – доля животной пищи в рационе. Оба подхода дали практически одинаковые результаты.
Сложность социальной организации тоже оценивалась двумя альтернативными способами – либо по среднему размеру группы, либо путем разделения всех видов на три категории: одиночки, живущие парами и живущие группами. Это тоже, как выяснилось, мало влияет на результаты.
Получилось, что обонятельные отделы мозга относительно крупнее у полуобезьян (Strepsirrhines) по сравнению с обезьянами (Haplorrhines), у видов с высококалорийной диетой, у ночных видов и живущих большими группами (рис. 10.3). Любопытно, что у последних по сравнению с видами, живущими парами или поодиночке, увеличены только дополнительные обонятельные луковицы, связанные с вомероназальным органом и восприятием социально значимых химических сигналов (феромонов), но не основные обонятельные луковицы, отвечающие за обычное обоняние. Это косвенно свидетельствует о важной роли химической коммуникации у социальных приматов, живущих большими группами (но не у моногамных видов и одиночек).
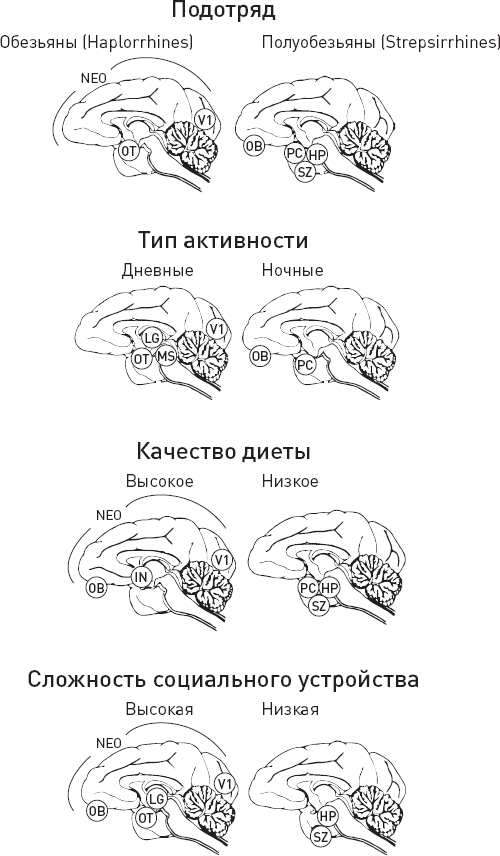
Рис. 10.3. Относительные размеры отделов мозга у приматов в зависимости от подотряда (Haplorrhines или Strepsirrhines) и социоэкологических факторов. Отмечены отделы мозга, относительно более крупные у данной группы по сравнению с другой: связанные со зрением (OT – зрительный тракт, V1 – первичная зрительная кора, LG – латеральное коленчатое тело таламуса, MS – средний мозг), с обонянием (OB – обонятельные луковицы), со вкусовым восприятием (IN – островок) и с пространственным мышлением (HP – гиппокамп, PC – палеокортекс, SZ – так называемый шизокортекс, включающий энторинальную кору и ряд других областей, тесно связанных с гиппокампом). Отдельно показан неокортекс (NEO), относительный объем которого увеличен у обезьян (по сравнению с полуобезьянами) и у видов со сложной социальной жизнью и высококачественной диетой (всеядных и плодоядных). По рисунку из DeCasien, Higham, 2019.
Зрительные отделы относительно крупнее у обезьян по сравнению с полуобезьянами, у дневных и сумеречных видов по сравнению с ночными, а также у видов с высококалорийной диетой и у живущих большими группами. Последний факт косвенно свидетельствует о важной роли визуальных сигналов в социальной жизни приматов.
Объем серого вещества в островковой коре оказался относительно больше у плодоядных по сравнению с листоядными, что может быть связано с улучшенным вкусовым восприятием, поскольку плодоядным приматам важно отличать на вкус спелые, сладкие фрукты от незрелых (ну а про листоедов и так понятно, что они не гурманы).
Относительный объем неокортекса больше у обезьян по сравнению с полуобезьянами, а также у видов, живущих большими группами, и у видов, потребляющих высококалорийную пищу. При этом калорийность диеты оказалась не менее, а даже чуть более надежным предиктором объема неокортекса, чем размер группы.
Отделы мозга, связанные с пространственным мышлением (это прежде всего гиппокамп и ряд прилегающих областей), относительно сильнее развиты у полуобезьян, а также у видов с низкокалорийной диетой и простой социальной организацией. Может быть, дело в том, что для высокосоциальных приматов, перемещающихся с места на место вместе с сородичами и живущих бок о бок с потенциальными половыми партнерами, не так критично умение ориентироваться в пространстве.
Некоторые подкорковые структуры и отделы ствола мозга (полосатое тело, бледный шар, таламус, гипоталамус, мозжечок, продолговатый мозг) относительно крупнее у полуобезьян. У видов с простой социальной структурой тоже сильнее развиты таламус, гипоталамус, мозжечок и продолговатый мозг, а также миндалевидные тела. Качество диеты положительно коррелирует с объемом субталамического ядра и отрицательно – с объемом мозжечка, гипоталамуса, эпиталамуса и продолговатого мозга.
Эти результаты в целом подтверждают идею о мозаичном характере эволюции мозга у приматов и о том, что особенности образа жизни, от которых зависит интенсивность отбора на те или иные сенсорные и когнитивные функции, могут приводить к изменению пропорций отделов мозга в ходе эволюции. Особенно хорошо это видно на примере зрительных и обонятельных отделов, относительные размеры которых явно зависят от того, насколько важную роль в жизни вида играют зрение или обоняние соответственно.
Тот факт, что высококалорийная диета, по-видимому, влияет на относительный объем неокортекса чуть ли не сильнее, чем сложность социальной организации, на первый взгляд, не очень хорошо согласуется с идеей о ведущей роли социальности в прогрессивном развитии мозга у приматов. Собственно, авторы исследования именно так и интерпретировали этот результат, полагая, что он ставит под сомнение теорию социального мозга.
Однако природа обнаруженной связи между диетой и объемом неокортекса может быть разной. С одной стороны, добывание высококалорийной пищи действительно может способствовать отбору на сообразительность. С другой стороны, хорошее питание ослабляет энергетические ограничения на рост неокортекса. То есть, возможно, оно не столько стимулирует эволюционный рост неокортекса напрямую, порождая отбор на когнитивные способности, сколько “разрешает” неокортексу увеличиться для выполнения иных, в том числе социальных, функций (книга 1, глава 2, раздел “Эректусы”, подраздел “Мозг, мясо и огонь”). А еще нужно помнить, что на свете живет очень много разных животных – и чем они только не питаются, однако мало кто достиг столь же впечатляющих когнитивных успехов, как обезьяны, тем более как человек. Вряд ли наши предки отрастили себе такой огромный мозг только потому, что предпочитали труднодобываемые деликатесы вроде мяса крупных животных в африканской саванне. Специфические пищевые предпочтения могли быть одним из важных факторов антропогенеза, но сами по себе они еще не дают положительной обратной связи в развитии мозга и когнитивных способностей, о которой говорилось выше.

