Книга: Кости, гены и культура
Назад: Денисовцы жили в Тибете 160 тысяч лет назад
Дальше: Глава 5 Тем временем в Африке
Данные по метилированию ДНК позволили воссоздать облик денисовского человека
Когда у ученых от всего денисовского племени была лишь одна косточка и один как следует прочитанный геном, то даже на таком более чем скудном материале были предприняты смелые попытки представить внешность этих людей. Как выглядела та девочка? Некоторые черты ее внешности удалось реконструировать. В геноме человека известны полиморфные нуклеотидные позиции (где у разных индивидов могут стоять разные нуклеотиды), по которым можно судить о таких фенотипических признаках, как цвет кожи, волос и глаз. Если соотношение между этими генетическими вариантами и внешними признаками у денисовцев было таким же, как у сапиенсов (что весьма вероятно, хотя и не известно наверняка), то девочка из Денисовой пещеры была брюнеткой с карими глазами и смуглой кожей без веснушек. Вот, пожалуй, и все. Но ученые на этом не остановились.
Научиться сколько-нибудь точно предсказывать фенотип по генотипу – важнейшая задача, стоящая перед современной биологией. Ее решение откроет прямо-таки фантастические перспективы перед медицинской генетикой, генной инженерией и другими дисциплинами – как теоретическими, так и самыми что ни на есть практическими. К сожалению, науке до этого еще далеко. Даже имея полный, качественно отсеквенированный геном, мы не так уж много можем сказать о фенотипе его обладателя.
Отдельные признаки, лежащие, что называется, на поверхности, – тот же цвет глаз, волос или кожи – еще кое-как можно реконструировать по характерным белок-кодирующим генам. Однако большинство интересных признаков (включая признаки скелета, такие как форма черепной коробки, размер зубов или пропорции костей таза) определяются не столько аминокислотными последовательностями отдельных белков, сколько нюансами регуляции активности (экспрессии) множества разных генов. Эти нюансы, в свою очередь, зависят от сложных сетей межгенных взаимодействий, чьи хитросплетения пока никто не умеет надежно расшифровывать, имея на руках лишь нуклеотидную последовательность генома. Теоретически задача должна быть решаемой – и когда-нибудь наука, будем надеяться, к этому придет. Но не сегодня и не завтра.
Однако можно поискать окольные пути. Например, было бы просто прекрасно, если бы удалось измерить уровень экспрессии генов в той же фаланге мизинца Denisova 3, из которой извлекли полный геном. Ведь уровень экспрессии – это некое приближение к оценке итогового результата работы всего запутанного клубка регуляторных взаимодействий. Чтобы измерить экспрессию, нужно было бы выделить из мизинца древнюю РНК. Вот только РНК разрушается еще быстрее, чем ДНК, и поэтому у нее меньше шансов сохраниться в древних костях. А ведь для оценки уровней экспрессии генов необходимо сравнивать количества молекул РНК, считанных с разных генов. Так что напрямую оценить экспрессию генов у древних гоминид на данный момент невозможно. И вряд ли когда-нибудь будет возможно. Впрочем, кто знает, от этих палеогенетиков всего можно ждать.
Но и здесь есть окольный путь, который палеогенетики недавно начали разведывать. Один из способов регуляции активности генов – это метилирование, присоединение метильной группы к цитозинам в промоторных областях генов, то есть там, откуда начинается считывание. Как правило, если у гена к промотору присоединено много метильных групп, то активность гена резко снижается. И мы можем посмотреть, какие цитозины в геноме метилированы, а какие нет. Это уже даст важную информацию об активности генов.
К счастью, метилирование цитозинов оставляет в древней ДНК различимые следы. Дело в том, что обычные, неметилированные цитозины в ходе посмертной деградации ДНК имеют обыкновение превращаться в урацилы, а метилированные – в тимины. На этой основе Давид Гохман из Еврейского университета в Иерусалиме и его коллеги из Израиля, Германии и Испании разработали метод реконструкции метиломов – профилей метилирования геномов ископаемых гоминид (Gokhman et al., 2014).
В исследовании, результаты которого были опубликованы в сентябре 2019 года в журнале Cell (Gokhman et al., 2019), ученые использовали метилом денисовского человека для реконструкции его облика (илл. IX на цветной вклейке). Метод, который они для этого разработали (рис. 4.3), основан на допущении, что фенотипические изменения, вызываемые сильным метилированием промотора, похожи на те, что возникают в результате мутаций, нарушающих работу данного гена или вовсе выводящих его из строя. Это правдоподобное допущение, потому что в обоих случаях функциональность соответствующего белка резко снижается.
В работе использовались метиломы денисовца (все той же девочки Denisova 3), двух неандертальцев (алтайского из Денисовой пещеры и хорватского из пещеры Виндия), пяти древних сапиенсов, живших от 45 до 7,5 тыс. лет назад, а также метиломы костей (это важно, ведь в разных тканях профили метилирования различаются!) пяти шимпанзе и пятидесяти пяти современных людей.
Первый этап исследования состоял в выявлении участков генома, уровень метилирования которых четко различается у разных видов (DMR, Differentially Methylated Regions). Учитывались только те участки, уровень метилирования которых мало зависит от возраста, пола, состояния здоровья и типа кости, а зависит только от видовой принадлежности. Такие DMR с большой вероятностью отражают именно эволюционные (а не возрастные, половые, тканеспецифичные или средовые) изменения уровня метилирования. Кроме того, рассматривались только самые сильные изменения. Отобранные для анализа участки должны были включать не менее 50 цитозинов, подходящих для метилирования (метилируются обычно цитозины, стоящие в ДНК перед гуанинами), и различаться у разных видов как минимум на 50 процентных пунктов: например, 80 % метилированных цитозинов у одного вида и лишь 30 % у другого. Для сравнения: колебания параметров среды обычно меняют уровень метилирования тех или иных участков генома не более чем на 10 %. Наконец, из всех DMR, удовлетворяющих этим условиям, для дальнейшего анализа были выбраны те, что расположены в промоторных областях белок-кодирующих генов (на расстоянии от 1 до 5 тысяч пар оснований от точки начала транскрипции), поскольку известно, что метилирование цитозинов именно на этих участках сильнее всего коррелирует с экспрессией гена.
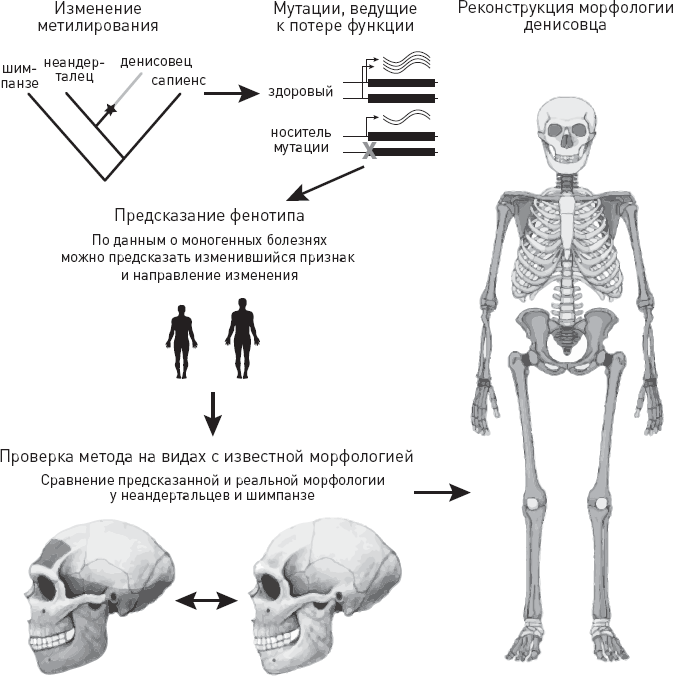
Рис. 4.3. Метод реконструкции фенотипа по данным о метилировании промоторных областей генов. Сначала выявляются гены, в промоторных областях которых у интересующего нас вида – в данном случае у денисовца – сильно изменился уровень метилирования по сравнению с ближайшими родственниками (неандертальцами и сапиенсами). Затем на основании данных по современным людям – носителям мутаций, выводящих данный ген из строя, делаются выводы, к каким фенотипическим изменениям могло привести изменение метилирования. При этом учитывается, что гиперметилирование промотора обычно ведет к снижению экспрессии гена, поэтому фенотипический эффект может быть похож на эффект мутации, ведущей к потере функции. Разработанный метод испытывается на видах с известной морфологией (неандертальцах и шимпанзе) и, наконец, используется для реконструкции морфологии денисовца. По рисунку из Gokhman et al., 2019.
После применения всех фильтров в изучаемой выборке DMR осталось 154 участка, уникальных для современных людей (то есть метилирование которых изменилось у сапиенсов по сравнению с другими гоминидами), 171 – метилирование которых изменилось у общих предков неандертальцев и денисовцев после их отделения от сапиенсов, 113 – специфичных для неандертальцев, 55 – специфичных для денисовцев, а также 2031 – по которым шимпанзе отличаются от всех видов людей. Для подавляющего большинства генов, в промоторах которых находятся отобранные DMR и по которым есть данные о корреляции между метилированием и экспрессией, эта корреляция отрицательна, то есть гиперметилирование соответствует пониженной экспрессии или полному отключению гена, а гипометилирование – высокой активности гена. Исключения из этого правила существуют, и они учитывались исследователями при разработке метода.
Чтобы на основании данных по DMR предсказывать морфологию, ученые использовали базу данных HPO (Human Phenotype Ontology), где содержится самая надежная и полная на сегодняшний день информация о том, как вредные мутации, ведущие к потере функции белка, сказываются на фенотипе. Из базы HPO были выбраны только те фенотипические изменения, которые, во-первых, затрагивают скелет, а во-вторых, имеют направленность. Например, “нарушение развития тазовых костей” не подходит под это условие, потому что не имеет направленности, а “укороченные подвздошные кости” – подходят. Всего в базе HPO удалось найти данные по 815 направленным изменениям скелета, которые наблюдаются у людей при поломке того или иного гена.
Эти данные опробовали на “предсказании” фенотипов двух видов с уже известной морфологией – неандертальцев и шимпанзе. Казалось бы, метод просто не может работать, ведь он основан на невероятно смелых допущениях и упрощениях! Тем не менее реконструкции шимпанзе и неандертальцев получились удивительно похожими на реальные. Например, неандертальцы отличаются от сапиенсов, как подсчитали авторы, по 107 признакам строения скелета. По метилому удалось “предсказать” больше половины (62) из этих отличительных признаков. В 46 случаях из 62 удалось предсказать не только сам признак, по которому неандертальцы отличаются от сапиенсов, но и направленность различий, причем в 36 случаях – правильно. Примерно такие же результаты получились и для шимпанзе.
Таким образом, метод оказался довольно эффективным, что вообще-то странно, учитывая, что метилирование промоторов – хотя и важный, но далеко не единственный способ регуляции активности генов. Отчасти успех может объясняться тем, что разные способы такой регуляции (включая метилирование ДНК, модификации гистонов и прикрепление регуляторных белков – факторов транскрипции – к регуляторным участкам ДНК) часто действуют согласованно, так что по одним признакам, указывающим на повышенную или пониженную активность гена, можно судить и о других. Иными словами, если мы видим, что у гена гиперметилирован промотор, то можно ожидать, что найдутся и другие признаки снижения активности гена, если их поискать.
Убедившись, что метод работает, ученые использовали его для реконструкции морфологии денисовского человека. Удалось выявить 56 отличий денисовцев (точнее, одного денисовца – все той же девочки Denisova 3) от неандертальцев и (или) современных людей. Направленность различий была предсказана в 32 случаях. Результаты по 18 признакам строения черепа (и 23 отличиям) суммированы на рисунке 4.4.
Получилось, что по большинству признаков, отличающих денисовца от современных людей, денисовцы были сходны с неандертальцами (мощные челюсти, низкий череп с широким основанием, низкий лоб, толстая зубная эмаль, широкий таз, крупная грудная клетка, расширенные кончики пальцев). При этом у денисовцев, в отличие от сапиенсов и неандертальцев, увеличен размер головки нижней челюсти, ширина черепа в теменной части и длина зубной дуги (последний факт согласуется с тем, что найденные денисовские зубы действительно очень крупные).
Когда это исследование было уже выполнено и статья проходила рецензирование, стали известны новые данные о тибетской челюсти, которая, как выяснилось, принадлежала денисовскому человеку (см. раздел “Денисовцы жили в Тибете 160 тысяч лет назад” выше). До этого момента из морфологически информативных денисовских образцов в распоряжении ученых имелись только зубы. Тибетская находка дала возможность проверить предсказания, связанные с четырьмя признаками нижней челюсти. Поскольку денисовец сравнивался по этим признакам с двумя другими видами (неандертальцами и сапиенсами), всего получается восемь предсказаний. Из них подтвердилось семь, что можно считать очень хорошим результатом. Единственное неподтвердившееся предсказание касается ширины “подбородка” (нижней челюсти в ее передней части): по метилому получилось, что денисовцы по этому признаку не отличаются от неандертальцев, а тибетская находка показала, что у денисовцев челюсть шире. Конечно, нужно иметь в виду, что тибетский денисовец имел полное право чем-то отличаться от денисовской девочки – как-никак он жил в другом регионе и на целую сотню тысячелетий раньше.
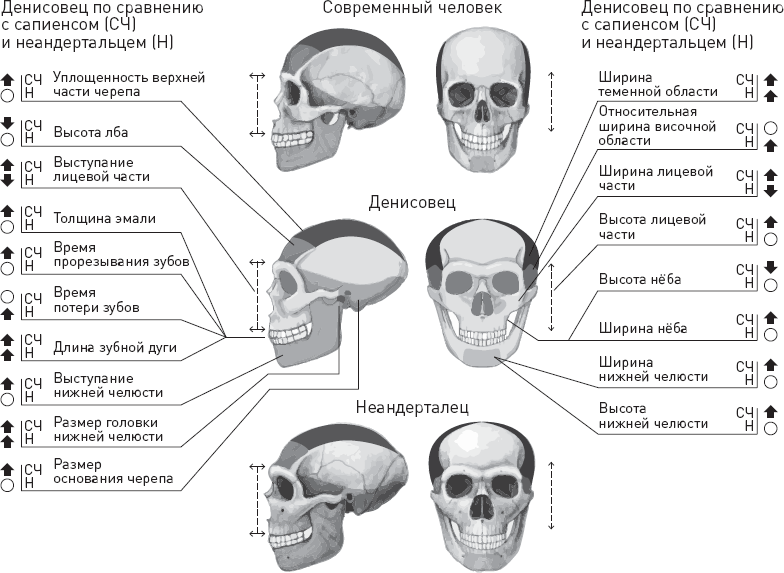
Рис. 4.4. Реконструкция черепа денисовского человека. Оттенками темно-серого цвета показаны реконструированные участки черепа и соответствующие им части черепов современного человека и неандертальца. Толстые стрелки, направленные вверх, означают, что данный признак у денисовца развит сильнее, чем у сапиенсов или неандертальцев (для прорезывания и утраты зубов это означает более раннее время), а направленные вниз – наоборот. Кружки означают отсутствие различий. Например, лоб у денисовца был ниже, чем у сапиенса, а от неандертальца по этому признаку отличий не было. По рисунку из Gokhman et al., 2019.
Получившаяся реконструкция особенностей скелета денисовского человека сильно напоминает некоторые спорные находки из Китая. Особенно много “денисовских” признаков у двух описанных в 2017 году черепов из Сюйчана возрастом от 100 до 130 тыс. лет (Li et al., 2017). Для тех частей черепа, которые сохранились у экземпляров из Сюйчана, по метилому предсказано семь отличий денисовцев от сапиенсов. Все эти семь отличительных особенностей (включая ширину теменной области, большую, чем у сапиенсов и неандертальцев) наличествуют у черепов из Сюйчана. Так что эти черепа с большой вероятностью принадлежат денисовцам.
Существенным ограничением предложенного метода реконструкции фенотипа по метилому является то, что он дает только качественные, но не количественные предсказания: можно предсказать, что какой-то признак будет выражен сильнее или слабее, но нельзя сказать насколько. Кроме того, во многих случаях даже направленность различий не удается предсказать, потому что метилирование разных генов, влияющих на один и тот же признак, направляет изменения в разные стороны – и никто не знает, какой из эффектов сильнее. Ничего нельзя сказать и о тех признаках, для которых пока нет данных о связи с какими-либо конкретными генами.
Что касается достоверности реконструкции денисовской морфологии, нужно помнить, что данные по метилому есть пока только для одного денисовца. Поэтому нельзя сказать наверняка, какие из реконструированных особенностей скелета характеризуют только девочку Denisova 3, а какие – всю денисовскую популяцию. Но все же есть основания полагать, что кое-какие предсказания справедливы для всех денисовцев. Во-первых, известно, что если мы возьмем случайно выбранного неандертальца и найдем у него какой-нибудь признак, по которому он отличается от всех сапиенсов, то в большинстве случаев этот признак позволит отличить от сапиенсов и всех остальных неандертальцев. Во-вторых, анализ был основан на участках генома, уровень метилирования которых в костях современных людей практически не зависит от возраста, пола, состояния здоровья и внешних воздействий и примерно одинаков у всех сапиенсов. Поэтому мы вправе надеяться, что степень метилирования этих участков у денисовцев тоже не зависела от всего перечисленного и что у других денисовцев она была примерно такой же, как у Denisova 3.
Еще одно интересное наблюдение состоит в том, что среди генов, чьи мутации связаны с известными наследственными болезнями человека, обнаруживается неожиданно много таких, которые как-то влияют на фенотипические признаки, недавно подвергавшиеся эволюционным изменениям. Показательно, что для морфологических отличий сапиенсов от неандертальцев в медицинских базах данных удалось найти “подходящие” гены в 70 % случаев (для 75 признаков из 107), тогда как для отличий человека от шимпанзе – только в 41 % случаев (для 83 признаков из 201). Возможно, это значит, что сбои в работе генов, недавно подвергавшихся действию отбора, с большей вероятностью приводят к медицинским проблемам по сравнению с генами, которые давно не подвергались эволюционным изменениям. Это согласуется с идеей о том, что стабилизирующему отбору нужно время, чтобы обеспечить устойчивость работы систем, подвергшихся действию движущего отбора. Те наши части и свойства, что развились недавно, – самые хрупкие.

