Книга: Кости, гены и культура
Назад: Предки неандертальцев и денисовцев скрещивались с потомками древних евразийских эректусов
Дальше: Глава 4 Эти загадочные денисовцы
Главной причиной поздне-четвертичного вымирания были люди, а не климат
Ученые давно спорят о том, что было главной причиной массового вымирания крупных животных в конце плейстоцена – начале голоцена: первобытные охотники или изменения климата (книга 1, глава 6). В последние годы чаша весов явно склоняется в сторону антропогенного фактора.
До недавних пор исследования в этой области носили, как правило, частный характер, то есть опирались на данные по отдельным регионам или отдельным группам животных, а немногочисленные попытки глобального анализа имели низкое пространственно-временное разрешение. Но в последние годы ситуация стала меняться в связи с накоплением детальных данных и по распространению позднечетвертичных млекопитающих, и по климатическим изменениям, и по истории расселения человеческих популяций.
В 2014 году исследователи из Орхусского университета в Дании проанализировали данные по всем видам млекопитающих с массой тела 10 килограмм и более на всех континентах за период от 132 тыс. лет назад (что соответствует началу предпоследнего, рисс-вюрмского, межледниковья) до 1 тыс. лет назад (Sandom et al., 2014). Данные анализировались отдельно по регионам, большинство из которых соответствует современным странам (самые крупные страны были разделены на несколько регионов). Для каждого региона было подсчитано абсолютное число и процентная доля видов, исчезнувших за исследуемый период.
Уровень вымирания в каждом регионе сопоставлялся с данными по климату и по истории заселения региона представителями рода Homo. Амплитуда климатических изменений оценивалась по разности среднегодовых температур между ледниковым максимумом, который был примерно 21 тыс. лет назад, и современностью (мы живем в теплую межледниковую эпоху) опять же по отдельности для каждого региона. Кроме того, было подсчитано изменение количества осадков при смене ледниковых и межледниковых эпох. В ходе моделирования учитывалась не только амплитуда, но и скорость климатических изменений, а также ряд дополнительных факторов, таких как рельеф местности (теоретически на равнине климатические изменения должны сильнее влиять на биоразнообразие, чем в гористой местности, потому что в горах животные могут мигрировать вверх-вниз, подлаживаясь к колебаниям температуры, а на равнине эта опция отсутствует). Впрочем, как выяснилось, учет (или неучет) подобных тонкостей мало влияет на результаты.
По своей демографической истории все регионы были разделены на пять групп:
1) “Прародина человечества” – Африка к югу от Сахары. Здесь крупные звери имели больше всего времени, чтобы постепенно приспособиться к двуногим хищникам. У них на эту коэволюцию были миллионы лет. Ну а те виды, которые приспособиться не сумели, вымерли задолго до позднечетвертичной эпохи (см. раздел “Вымирание африканских хищников последние два миллиона лет направлялось эволюцией гоминид, а не климатом” в главе 1).
2) Регионы, заселенные древними Homo (эректусами) в ходе первой волны расселения людей за пределами Африки. Здесь у мегафауны тоже было достаточно времени для коэволюции, а наиболее уязвимые виды тоже вымерли до рассматриваемого интервала времени.
3) Регионы, заселенные представителями второй волны выходцев из Африки – неандертальцами и денисовцами.
4) Регионы Северной Евразии, где древние Homo вроде бы не жили и где не было эндемичных видов крупных млекопитающих. Все виды, что там водились, могли встречаться с древними Homo в южных частях своих ареалов.
5) Регионы, которые впервые были заселены людьми современного типа с их чрезвычайно эффективными приемами охоты на крупного зверя и оружием. Там у крупных млекопитающих не было возможности постепенно адаптироваться к новому беспощадному хищнику.
В общей сложности ученые насчитали 177 видов крупных млекопитающих, вымерших в интервале от 132 до 1 тыс. лет назад либо полностью, либо на одном из континентов (18 видов вымерло в Африке, 38 – в Азии, 26 – в Австралазии, 19 – в Европе, 43 – в Северной Америке, 62 – в Южной Америке). Самое интенсивное вымирание (как по абсолютному числу видов, так и по процентной доле) происходило на юге Южной Америки, на юго-востоке Северной Америки, в Европе и Австралии. Минимальный уровень вымирания отмечен в Африке к югу от Сахары и в Южной Азии.
Статистический анализ показал, что объяснить наблюдаемую географическую структуру вымираний одними лишь климатическими изменениями невозможно. В глобальном масштабе обнаруживается лишь слабая связь между амплитудой климатических изменений и уровнем вымирания. Причем эта связь достоверна только для Евразии и только для данных по температуре (но не по осадкам), а на остальных континентах вообще никакой корреляции между климатом и вымиранием не прослеживается. Например, в Южной Америке, где уровень вымирания был максимален, климат менялся вообще незначительно. В целом климат может объяснить лишь 20 % региональных различий по уровню вымирания. Гораздо большей предсказательной силой обладают модели, основанные на истории заселения регионов людьми (такие модели объясняют до 64 % различий). Наименьший уровень вымирания характерен для прародины человечества – Африки, промежуточный – для регионов, постепенно заселявшихся древними Homo, наибольший – для областей, где первопоселенцами были сапиенсы. Модели, комбинирующие историко-демографические факторы с климатическими, предсказывают уровень вымирания лишь ненамного лучше, чем чисто демографические, причем это улучшение заметно опять-таки только для Евразии.
Таким образом, имеется сильная зависимость между интенсивностью позднечетвертичного вымирания и продолжительностью периода коэволюции людей и крупных млекопитающих. Чем позже появились в регионе люди, тем более жестокому вымиранию подверглась мегафауна в позднем плейстоцене и голоцене. Пожалуй, еще важнее вывод об отсутствии значимой корреляции между уровнем вымирания и величиной климатических изменений на всех континентах, кроме Евразии (да и там корреляция слаба).
В статье приведено несколько показательных примеров. Так, многие вымершие южноамериканские крупные растительноядные, например гигантский ленивец Eremotherium laurillardi, представитель хоботных Stegomastodon waringi и представитель местных копытных Toxodon platensis, при необходимости легко переходили с одной диеты на другую (таких животных называют трофическими генералистами). Поэтому при смене ландшафтов они должны были просто изменить спектр питания. Их вымирание трудно объяснить чередованием влажных и засушливых периодов, как это пытались сделать некоторые исследователи. Тем более что эти климатические флуктуации в Южной Америке были уж точно не сильнее, чем в Африке, где уровень вымирания был гораздо ниже. В некоторых районах Северной и Южной Америки за рассматриваемый период не только не произошло существенных климатических изменений, но и растительность осталась практически такой же, как 130 тыс. лет назад. Однако крупные животные там все равно вымерли: и трофические генералисты, и травоядные, и хищники, охотившиеся на них. Например, в Калифорнии, где господствующие растительные комплексы за 130 тыс. лет почти не изменились, вымер 21 вид (53 %) крупных зверей. Среди них – многие виды, чьи предпочитаемые кормовые растения растут в изобилии по сей день, так что о подрыве кормовой базы говорить не приходится. Это относится, например, к мастодонту Mammut americanum, питавшемуся ветвями болотного кипариса, которого и сейчас сколько угодно в местах, где жил этот вымерший гигант. Вымерли растительноядные ленивцы Nothrotheriops shastensis, вымерли родственники современных вилорогов Capromeryx minor, вымер и ужасный волк, Canis dirus.
Полученные результаты – весомый аргумент в пользу антропогенного объяснения позднечетвертичных вымираний. Приходится признать, что характерное для последних веков и тысячелетий крайне низкое разнообразие или полное отсутствие крупных млекопитающих на большей части суши – вовсе не “естественное” состояние вещей, а прямой результат истребления мегафауны нашими предками. Исследователи подчеркивают (и с ними трудно не согласиться), что данный факт необходимо учитывать при планировании мероприятий по сохранению и восстановлению природных экосистем.
К сходным выводам привело ученых и другое масштабное исследование, основанное на совершенно ином подходе и других данных.
Важным косвенным аргументом против антропогенной версии позднечетвертичного вымирания, затронувшего в основном крупных животных, считается тот факт, что крупные животные вообще более уязвимы, поскольку их популяции имеют низкую численность. Так что различные природные катаклизмы, в том числе климатические, по идее, должны приводить к избирательному вымиранию крупных зверей. Результат будет выглядеть точно так же, как и при избирательной охоте, нацеленной на крупную дичь, ведь именно такую и предпочитали, скорее всего, первобытные охотники.
Если позднечетвертичное вымирание не связано с деятельностью человека, то мы вправе ожидать, что крупные виды млекопитающих вымирали чаще, чем мелкие, не только в конце четвертичного периода, но и в другие, более древние эпохи. В том числе и когда никаких людей еще в помине не было (а климат-то менялся в кайнозое многократно).
Можно проверить, насколько разнятся скорости вымирания мелких и крупных животных в течение кайнозоя, обратив особое внимание (естественно!) на поздний плейстоцен, когда сапиенсы начали хозяйничать повсюду. Такую проверку провели в 2018 году ученые из четырех американских университетов (Smith F. A. et al., 2018).
Исследователи проанализировали данные по пространственно-временному распространению всех видов нелетающих наземных кайнозойских млекопитающих, по которым есть достаточно точные датировки первых и последних находок. Таких видов набралось 2786. Для разных временных интервалов оценивалась скорость вымирания видов в зависимости от массы тела. Кайнозой поделили на отрезки длительностью в один миллион лет, и для каждого отрезка рассчитали степень избирательности вымирания в зависимости от размера животных.
Отдельная база данных была составлена по позднечетвертичным (от 125 тыс. лет назад до настоящего времени) и современным видам. Если вид существует сегодня, но не известен в ископаемом состоянии, то считалось, что 125 тыс. лет назад он уже существовал. Позднечетвертичное время поделили на четыре интервала, соответствующие разным этапам экспансии средне– и верхнепалеолитических охотников (рис. 3.8):
1) “Поздний плейстоцен” (125–70 тыс. лет назад). В это время в Европе процветали неандертальцы, в Азии – вероятно, денисовцы, а группы сапиенсов начали периодически выходить за пределы Африки.
2) “Конец плейстоцена” (70–20 тыс. лет назад). На этот период приходится быстрое расселение сапиенсов по Евразии, а также заселение Австралии.
3) “Терминальный плейстоцен” (20–10 тыс. лет назад). В это время сапиенсы заселили Америку.
4) Голоцен (10–0 тыс. лет назад).
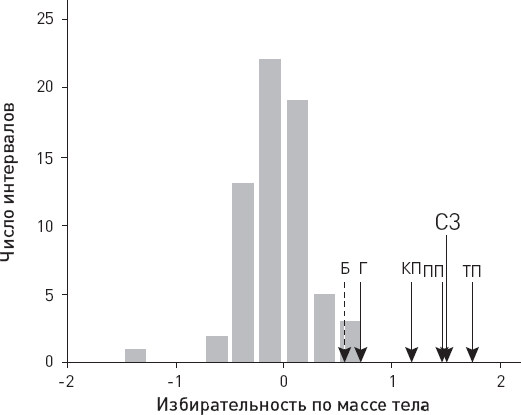
Рис. 3.8. Избирательность вымирания млекопитающих в зависимости от массы тела в течение кайнозоя. Значения на горизонтальной оси характеризуют зависимость вероятности вымирания вида от массы тела. Положительные значения соответствуют преимущественному вымиранию крупных животных, отрицательные – мелких, близкие к нулю – примерно одинаковой вероятности вымирания для крупных и мелких животных. Высота столбцов отражает количество интервалов кайнозойской эры (длительностью по 1 млн лет каждый), когда наблюдался данный уровень избирательности. Стрелками показаны уровни, характерные для позднечетвертичного времени, которое рассматривалось отдельно: ПП – “поздний плейстоцен” (125–70 тыс. лет назад), КП – “конец плейстоцена” (70–20 тыс. лет назад), ТП – “терминальный плейстоцен” (20–10 тыс. лет назад), Г – голоцен (10–0 тыс. лет назад), СЗ – среднее значение за всю позднечетвертичную эпоху (125–0 тыс. лет назад), Б – будущее, прогноз на ближайшие 200 лет (основан на предположении, что вымрут все виды, ныне находящиеся под угрозой). По рисунку из Smith F. A. et al., 2018.
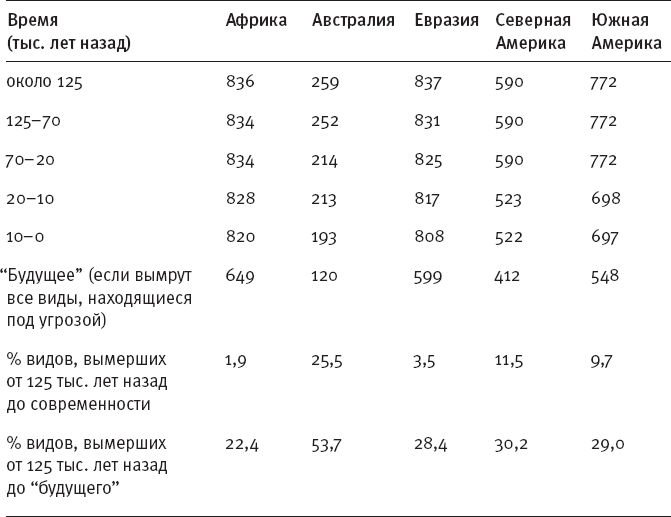
Общий масштаб позднечетвертичного вымирания отражен в таблице 1.
Оказалось, что в течение почти всей кайнозойской эры вымирание млекопитающих было неизбирательным по размеру. На рисунке 3.8 это выражается в том, что самые высокие столбцы расположены недалеко от нулевой отметки. Вымирали с равной вероятностью как мелкие, так и крупные виды. Около 29 млн лет назад, в олигоцене, был даже эпизод избирательного вымирания мелких млекопитающих, предположительно связанный с началом глобального похолодания, что привело к уменьшению площади лесов и распространению травяных биомов (степей и саванн).
Табл. 1. Число видов нелетающих наземных млекопитающих на разных континентах в течение позднего плейстоцена и голоцена. Нижние три строки показывают, на сколько процентов снизилось видовое разнообразие с начала позднего плейстоцена по настоящее время, а также ожидаемые потери в будущем, если вымрут все краснокнижные виды (Smith F. A. et al., 2018).
Принципиально иная ситуация сложилась в позднечетвертичную эпоху (стрелки на рис. 3.8). Уже в начале позднего плейстоцена, 125–70 тыс. лет назад, вымирание стало резко избирательным. Начиная с этого времени вымирали в основном крупные животные. Данная тенденция сохранялась и в дальнейшем – вплоть до настоящего времени. Правда, в голоцене избирательность немного сгладилась. Скорее всего, сглаживание связано с тем, что с развитием сельского хозяйства люди стали истреблять фауну другими способами. Охота отступила на второй план, а на первый вышло уничтожение местообитаний, от которого страдают не только крупные, но и мелкие животные.
Главный вывод состоит в том, что резко повышенная вероятность вымирания крупных животных, характерная для позднечетвертичного времени, вовсе не является универсальной закономерностью. В течение большей части кайнозоя такой избирательности не было. Это уникальная особенность именно позднечетвертичного вымирания, когда к издревле существовавшим причинам вымирания добавился новый суперхищник, овладевший эффективными методами охоты на крупных животных.
Избирательность позднечетвертичного вымирания выглядит уникальной не только на фоне предшествующей динамики разнообразия кайнозойских млекопитающих. Преимущественное вымирание крупных животных отмечается во время некоторых кризисов. Например, в конце девона крупные позвоночные, представленные в то время в основном всевозможными рыбами, пострадали, по-видимому, сильнее, чем мелкие. Но все же столь сильного перекоса в сторону вымирания крупных видов, как у млекопитающих в позднем плейстоцене и голоцене, во время других масштабных вымираний, судя по всему, не было.
Вывод о новом суперхищнике становится еще очевиднее при детальном рассмотрении полученных данных. С самого начала кайнозоя и вплоть до плейстоцена млекопитающие в среднем становились все крупнее. Росла как средняя, так и максимальная масса тела существующих видов. Однако в позднем плейстоцене тенденция сменилась на противоположную. За последние 125 тыс. лет фауна млекопитающих сильно измельчала, причем динамика этого процесса согласуется с историей расселения первобытных охотников. Так, в Евразии размер млекопитающих пошел на убыль уже 125–70 тыс. лет назад. Здесь нужно особо подчеркнуть, что в Евразии в то время охотились неандертальцы и денисовцы, а не сапиенсы. Сапиенсы в это время по большей части еще оставались в Африке, лишь изредка совершая небольшие экскурсии в Азию. Видимо, дело не в специфичности нашего вида, а в адаптации всех потомков гейдельбергских людей к охотничьей жизни. Неандертальцы и денисовцы, вероятно, были хорошими охотниками, так что при невысокой численности своих популяций они смогли оказать заметное влияние на популяции крупных животных.
В Австралии катастрофическое измельчание млекопитающих приходится на период от 70 до 20 тыс. лет назад, что совпадает с появлением там первых людей (около 50 тыс. лет назад). В этот период Северная и Южная Америка, куда люди еще не добрались, ненадолго стали подлинными заповедниками мегафауны: обитавшие там звери были крупнее, чем где бы то ни было на планете. Резкое измельчание американских млекопитающих произошло позже – в “терминальном плейстоцене” (20–10 тыс. лет назад), когда туда прорвались люди из Берингии (обо всех этих миграциях сапиенсов мы еще поговорим).
Что касается Африки, то на этом континенте позднечетвертичное вымирание мегафауны было выражено слабее всего. Это традиционно объясняют тем, что в Африке крупные звери коэволюционировали с гоминидами миллионы лет. Поэтому у них была возможность постепенно адаптироваться к уловкам плотоядных двуногих обезьян. Кроме того, не исключено, что в результате этой коэволюции размерное распределение африканских млекопитающих сместилось в сторону измельчания намного раньше, чем на остальных континентах. Африканские виды млекопитающих по своему среднему размеру заметно уступали евразийским и американским (и были сравнимы с австралийскими) уже в начале позднего плейстоцена.
Здесь нужно учитывать, что одним из факторов, способствующих эволюции крупной фауны, считается площадь континента. Этим объясняют, в частности, относительно более мелкие размеры представителей вымершей австралийской мегафауны по сравнению с плейстоценовыми млекопитающими Евразии и Америки. Тот факт, что уже 125 тыс. лет назад африканская фауна в среднем была ненамного крупнее австралийской, исследователи предположительно связали с влиянием древних гоминид – и это отлично согласуется с данными о вымирании крупных африканских хищников, о котором мы говорили в разделе “Вымирание африканских хищников последние два миллиона лет направлялось эволюцией гоминид, а не климатом” в главе 1.
Исследователи попытались также сделать прогноз на будущее, основанный на допущении, что вымрут все краснокнижные звери. Это приведет к еще большему измельчанию млекопитающих, а также к дальнейшему росту доли грызунов и снижению доли копытных в экосистемах.
Получается, что деятельность гоминид сильно влияла на фауну млекопитающих и структуру сообществ еще в доисторические времена. Антропогенное вымирание видов, происходящее в наши дни, является не более чем продолжением давней тенденции. Если дело и дальше так пойдет, то через пару веков фауна диких млекопитающих станет в основном фауной грызунов, а самым крупным наземным животным будет домашняя корова.
В этом исследовании есть один скользкий момент, связанный с неполнотой палеонтологической летописи. При анализе позднечетвертичного разнообразия учитывались данные по современным видам. Принималось, что все современные виды существовали уже в позднем плейстоцене, даже если по ним нет ископаемых находок. С одной стороны, это наверняка справедливо для большинства видов, поскольку средняя продолжительность существования видов у млекопитающих составляет 1–2 млн лет, так что если вид существует сегодня, то с большой вероятностью он существовал и 125 тыс. лет назад. С другой стороны, мелкие кости, как правило, хуже сохраняются в ископаемом состоянии, чем крупные. Иными словами, не исключено, что палеонтологическая летопись крупных животных в принципе полнее. Поэтому учет современных животных наравне с ископаемыми мог искусственно завысить долю мелкого зверья в позднечетвертичное время по сравнению с остальными эпохами кайнозоя, для которых использовались только палеонтологические данные. И даже не просто мелкого, а мелкого и не вымершего. Это могло привести к завышению уровня избирательности позднечетвертичного вымирания. Хорошо бы понять, насколько серьезно данный эффект повлиял на выводы.
Назад: Предки неандертальцев и денисовцев скрещивались с потомками древних евразийских эректусов
Дальше: Глава 4 Эти загадочные денисовцы

