Книга: Кости, гены и культура
Назад: Больше хоббитов, хороших и разных
Дальше: Глава 3 Наследники питекантропов в Европе и Азии
Человек из Диналеди – еще один вид примитивных людей
В послужном списке южноафриканского антрополога Ли Бергера немало достойных открытий. О некоторых из них мы рассказали в первой книге. Например, в 2006 году Бергер переисследовал знаменитый череп юного австралопитека из Таунга и доказал, что ребенок стал добычей крупной хищной птицы (книга 1, глава 1, раздел “Австралопитеки”, подраздел “Юные австралопитеки были добычей хищных птиц”). В 2010 году последовало открытие нового вида австралопитеков (книга 1, глава 2, раздел “Australopithecus sediba – австралопитек, похожий на человека”).
Чем занимался этот неутомимый и азартный антрополог в течение последующих четырех с половиной лет, читатели узнали из двух публикаций в журнале eLife в 2015 году (Berger et al., 2015; Dirks et al., 2015). Как выяснилось, он вел раскопки в пещере Райзинг-Стар в 50 километрах от Йоханнесбурга (Южная Африка). В течение двух полевых сезонов – осенью 2013-го и весной 2014-го – его команда исследовала человеческие остатки, найденные двумя спелеологами в одной из камер пещеры. Пещера имеет очень сложную форму с несколькими залами и узкими вертикальными и горизонтальными тоннелями-“шкуродерами”, пройти которые полностью удалось, скорее всего, впервые.
Чтобы протиснуться в узкие лазы, имеющие в некоторых местах ширину всего 20–25 сантиметров, пришлось поискать для работы худых спелеологов (понятно, что для этого лучше всего подошли стройные юные девушки). Так или иначе, но пещерные тоннели были пройдены, и в последней камере, именуемой Диналеди, была найдена богатейшая коллекция человеческих остатков. Наверх для исследования было поднято около 1500 костей и зубов, все достойной сохранности, некоторые скелеты удалось собрать практически полностью. Почти все кости принадлежат гоминидам, среди них только 6 птичьих и несколько плохо определимых остатков грызунов.
Позже была обследована еще одна камера в другой части той же пещеры – камера Леседи, расположенная в 60 метрах от Диналеди по прямой (Hawks et al., 2017). Однако прямого сообщения между камерами нет, и проползти из одной в другую можно только кружным путем протяженностью в 145 метров. В Леседи, как и в Диналеди, гоминиды с большой вероятностью попадали не в виде разрозненных частей, а целиком. В этой камере найдено 133 костных фрагмента, сгруппированных в три скопления. Кости принадлежат как минимум трем особям разного возраста, но, вероятнее всего, их было больше. Кости в обеих камерах имеют одни и те же характерные признаки, так что нет сомнений в их единой видовой принадлежности. Ученые полагают, что люди из Диналеди и Леседи почти наверняка принадлежали к одной и той же популяции.
Кроме остатков гоминид, в Леседи найдены кости нескольких видов мелких млекопитающих. В основном это грызуны и землеройки. Есть также собачий зуб и остатки дикого кота – это самые крупные представители местной ископаемой фауны. Скорее всего, это связано с труднодоступностью камер Диналеди и Леседи. Судя по всему, в древности пробраться туда было не проще, чем сегодня.
По числу костных остатков, их концентрации и сохранности это местонахождение уникально. За прошедшие века захоронение не потревожили ни звери, ни птицы, ни туристы, и оно сохранилось в своем первоначальном состоянии, покрываясь постепенно тонкими наносами глинистой пыли.
Теперь Ли Бергеру и его коллегам нужно было понять, чьи эти кости, как они туда попали и из какого они времени.
Ответ на первый вопрос такой: все кости бесспорно принадлежат гоминидам одного вида, и это новый вид людей, получивший наименование Homo naledi (илл. VI на цветной вклейке). Отнесение к роду Homo основано на анализе морфологических признаков скелета и сравнении со всеми известными гоминидами и другими человекообразными обезьянами. Благо материала для сравнения было предостаточно – и он был тщательно изучен и педантично описан.
Люди из Диналеди имели небольшой рост (144–148 см) и вес (около 39–45 кг), некрупные зубы, не исключая и маленькие клыки. Жевательные поверхности коренных зубов относительно простые. Ходили эти люди на двух ногах, уже вполне человеческих, если не считать небольшого плоскостопия. На таких ногах, более длинных, чем у австралопитеков, можно было совершать дальние походы. Кисти рук у них тоже были больше похожи на человеческие – с удлиненным большим пальцем и подвижным запястьем, правда, пальцы были кривоваты. Удивительно при этом, что их ребра и плечи соответствуют австралопитековой морфологии, то есть годятся для лазанья. Искривленные фаланги пальцев тоже напоминают пальцы австралопитеков. Крепления бедер и вся тазовая часть также скорее напоминают австралопитеков. Так что перед нами вид гоминид, сочетающий черты людей и австралопитеков. Примерно такая же история была и с Australopithecus sediba, которого Бергер и его коллеги описали ранее, но тогда по “сумме баллов” черты австралопитеков перевесили человеческие.
Самая ценная находка из Леседи – неплохо сохранившийся череп крупной особи, к которому прилагается значительная часть посткраниального скелета. Данный индивид получил условное имя LES1. Объем его мозговой коробки составлял 610 см3, что заметно больше, чем у двух черепов из Диналеди, для которых ранее удалось реконструировать объем мозга (465 и 560 см3). Таким образом, самый мозговитый из трех представителей H. naledi решительно превосходит по данному показателю грацильных австралопитеков и хоббитов с Флореса, приближаясь к средним значениям, характерным для H. habilis (рис. 2.3).
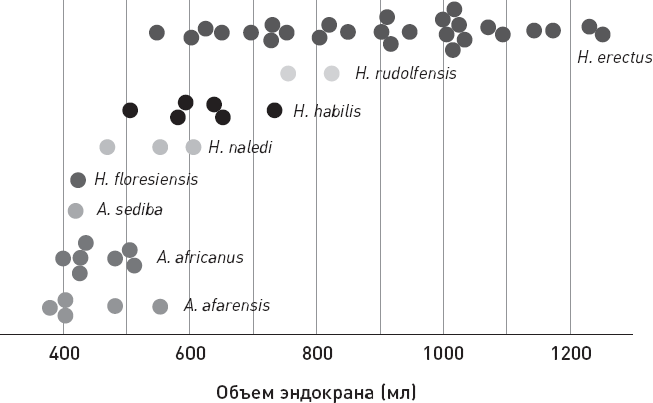
Рис. 2.3. Объем эндокрана Homo naledi по сравнению с другими гоминидами. По рисунку из Hawks et al., 2017.
При скромном размере мозга, сопоставимом с размером мозга хабилисов, у H. naledi оказались совсем небольшие зубы, какие характерны для людей современного типа. Раньше считалось, что тенденция к увеличению мозга тесно связана с уменьшением челюстей и зубов. Предполагалась примерно такая причинно-следственная связь: улучшенное питание (рост доли мяса в рационе, приготовление пищи на огне) → редукция жевательного аппарата → ослабление ограничений на рост мозга, накладываемых наличием могучих челюстей, зубов и жевательной мускулатуры. H. naledi не слишком хорошо укладывается в эту схему, как, впрочем, и A. sediba и H. floresiensis.
Ни детальный анализ морфологии, ни сравнение с другими видами ископаемых гоминид не позволили ученым сделать однозначного вывода о положении H. naledi на эволюционном дереве. Одни признаки сближают людей из пещеры Райзинг-Стар с ранними Homo (особенно с H. habilis) или с австралопитеками (особенно с A. sediba), другие – с эректусами, третьи своеобразны и могут свидетельствовать об “особом эволюционном пути”, четвертые (включая некоторые особенности зубов, кистей и стоп) напоминают поздних Homo. Но в целом H. naledi явно ближе по своей морфологии к ранним Homo с маленьким мозгом, чем к поздним большеголовым представителям рода. Хотя ко времени жизни людей из Диналеди первые давно вымерли, а вторые процветали (датировку образцов из Диналеди мы обсудим чуть позже).
Люди из Диналеди имели руки, способные к мелким и точным манипуляциям, стопы, приспособленные для двуногого хождения, а также небольшие зубы – признак, эволюционное формирование которого иногда связывают с пониженной внутригрупповой агрессией (книга 1, глава 1, раздел “Семейные отношения – ключ к пониманию нашей эволюции”). Но при этом мозг у H. naledi оставался маленьким. Означает ли это, что весь “человеческий комплекс” – двуногость, трудовая деятельность, социальная конформность – не требует большого мозга? И зачем тогда людям большой мозг?
Это исключительно важный вопрос, к которому мы вернемся в заключительных главах. А пока лишь заметим, что у флоресских хоббитов тоже маленький мозг (скорее всего, вторично уменьшившийся), но хоббиты при этом сохранили и кое-какие охотничьи навыки, и примитивную каменную индустрию, и даже огонь. Так что в целом понятно, что для подобной деятельности мозг не обязательно должен быть большим. Улучшить функциональность мозга, очевидно, можно не только путем простого наращивания его массы и числа кортикальных нейронов, но и путем структурной оптимизации. Другое дело, что увеличение объема мозга, возможно, является самым “простым” (высоковероятным с эволюционной точки зрения) способом этого добиться. Можно даже предположить, что быстрый рост мозга у наших предков в течение последних двух миллионов лет был своего рода “несчастным случаем”: отбор на улучшение когнитивных способностей был настолько сильным, что у эволюции попросту не хватало времени на тонкие структурные перестройки, и поэтому она пошла самым простым путем – безудержного разрастания, невзирая на все издержки (см. главу 12).
Но вернемся в пещеру Райзинг-Стар. Одна из главных загадок – как попали все эти люди в глубокую и труднодоступную пещеру. Геологи утверждают, что других входов в пещеру и следов обвалов нет. Это означает, что в древности, как и сегодня, во внутренние камеры пещеры попасть было нелегко. Тем не менее все признаки указывают на то, что останки (или еще живые люди) были доставлены в пещеру целиком, а не по частям. Так что версию с хищниками, таскавшими в укромное место свою трапезу, можно отбросить. Также пришлось оставить и гипотезу о лавинах и потоках, ставших причиной смерти и смывших мертвые тела в глубины пещеры. В таком случае на костях остались бы характерные следы.
Версия о том, что пещера была местом ритуальных захоронений, встречает серьезное возражение: в этом случае нужно было бы ожидать большую долю захоронений пожилых людей. Но среди умерших пожилым можно считать только одного человека. Остальные были моложе: трое детей, четверо подростков и четверо взрослых. Однако Бергер и его коллеги не исключают гипотезу о намеренном захоронении и даже всячески ее пропагандируют. Ведь детская смертность, напоминают они, во все времена была исключительно высока. Впрочем, всерьез обсуждать эту гипотезу явно преждевременно – экстраординарные утверждения требуют экстраординарных доказательств, а их пока нет.
Открытие Homo naledi стало громкой сенсацией в палеоантропологии, однако по-настоящему оценить значимость открытия было невозможно до тех пор, пока оставалось неизвестным чуть ли не самое главное – возраст находок.
Как мы помним, в морфологии H. naledi примитивные “австралопитековые” признаки сочетаются с продвинутыми “человеческими”. Если бы эволюция наших предков была линейной, как многие думали еще лет 30–40 назад, то есть представляла собой непрерывное прогрессивное развитие от австралопитеков до Homo sapiens, то H. naledi логично было бы поместить где-нибудь рядом с хабилисами. В таком случае ожидаемый возраст H. naledi составлял бы порядка полутора-двух миллионов лет.
Впрочем, на сегодняшний день твердо установлено, что эволюция гоминид вовсе не была линейной. На той ветви эволюционного дерева, что включает формы, более близкие к современному человеку, чем к шимпанзе (представителей этой ветви мы как раз и называем гоминидами, хотя многие авторы пользуются термином “гоминины”, что, на наш взгляд, менее благозвучно), было множество развилок и тупиковых веточек. Направления специализации в разных линиях могли сильно различаться, а “эволюционно продвинутые” (сильно отличающиеся от общего предка) виды нередко сосуществовали с “примитивными” (сохранившими больше предковых черт).
Из этого следует, что определить реальный возраст того или иного вида гоминид только по его морфологии просто-напросто невозможно. H. naledi мог оказаться древним представителем одной из веточек ранней радиации рода Homo. Он мог даже оказаться прямым предком эректусов и сапиенсов, хотя по комбинации признаков H. habilis лучше подходит на эту роль. С тем же успехом он мог оказаться тупиковой ветвью или архаичным потомком, сохранившим примитивные черты в те времена, когда другие представители человеческого рода уже далеко продвинулись по пути увеличения мозга и усложнения поведения. Подобные примеры были известны и ранее. Самый яркий из них – конечно, хоббиты с острова Флорес, имевшие мозг обезьяньего размера, но жившие относительно недавно, 90–60 тыс. лет назад (см. раздел “Новые данные о хоббитах с острова Флорес” выше).
Поэтому антропологи с нетерпением ждали, когда же появятся хоть какие-то датировки H. naledi – если, конечно, эти кости, найденные в необычном геологическом контексте, вообще удастся датировать. И вот наконец в мае 2017 года в журнале eLife появились большие статьи Ли Бергера и его коллег с бесценными новыми сведениями о H. naledi (Dirks et al., 2017; Berger et al., 2017).
Первая статья (Dirks et al., 2017) целиком посвящена датировкам. Как выяснилось, Бергер и его команда за четыре года, прошедшие с момента обнаружения костей H. naledi, проделали колоссальный объем работы. Для получения надежных дат ученые перепробовали буквально все существующие методы, которые в принципе можно было применить к имеющемуся материалу. Чтобы исключить влияние “человеческого фактора”, анализы проводились параллельно в разных лабораториях слепым методом: сотрудники лабораторий не знали ни о происхождении образцов, ни о результатах, полученных в других лабораториях.
Некоторые человеческие кости в камере Диналеди просто лежали на поверхности, изъятые из породы и очищенные естественными процессами, другие находились в толще пещерных отложений. Эти отложения сильно отличаются по своему строению от отложений в других пещерах данного района, где ранее были найдены ископаемые остатки нескольких видов гоминид. Район этот, кстати, включен в список ЮНЕСКО как памятник всемирного наследия под названием “Колыбель человечества”.
Отложения, содержащие кости H. naledi, сформировались внутри пещеры и представляют собой неконсолидированную (неокаменевшую, сыпучую) тонкозернистую породу с прослоями натечных образований. Известковые натеки удалось датировать при помощи уран-ториевого метода. Результаты, полученные в разных лабораториях, довольно точно совпали друг с другом. Анализ множества образцов, сформировавшихся как до попадания в пещеру костей H. naledi, так и после, позволил понять историю формирования пещерных отложений. В частности, стало ясно, что влажные периоды, когда образовывались натеки, чередовались с относительно сухими. Эти и другие детали, выявленные в ходе исследования, помогали ученым шаг за шагом подбирать и оттачивать аналитические подходы, постепенно сокращая неопределенность в главном вопросе о возрасте костей.
Уран-ториевый анализ зубов H. naledi, а также зуба павиана, попавшего в пещеру намного раньше, помог расшифровать многоэтапную историю привноса урана в ископаемые зубы, происходившего во влажные периоды. Привнос урана с дождевой водой или грунтовыми водами в кости после захоронения ведет к занижению датировок (омоложению), поэтому данный подход в итоге лишь показал, что зубы H. naledi наверняка древнее 70 тыс. лет и с большой вероятностью древнее 200 тыс. лет.
К тем же зубам был применен комбинированный метод урановых серий и электронного спинового резонанса, а к пробам вмещающей породы – метод оптически стимулируемого люминесцентного датирования. Эти подходы дали самые надежные результаты. Прослои натечных образований были также подвергнуты палеомагнитному анализу. Прочие методы датирования, которые исследователи пытались применить, в том числе радиоуглеродный и уран-свинцовый, оказались по тем или иным причинам непригодными для данного материала.
В итоге в распоряжении авторов оказалось множество датировок, полученных несколькими независимыми способами как для костей, так и для минералов из разных слоев пещерных отложений, одни из которых сформировались заведомо до образования костеносного слоя, а другие – после. Анализ всей совокупности данных привел исследователей к выводу, что возраст костей почти наверняка находится в интервале от 236 до 335 тыс. лет.
Таким образом, люди из Диналеди жили гораздо позже, чем можно было предположить по их морфологии. Для своего времени они были архаикой, “пережитками прошлого” – примитивными современниками продвинутых поздних представителей человеческого рода, не уступавших нам по размеру мозга, владевших огнем и изощренными (позднеашельскими и среднепалеолитическими) технологиями обработки камня.
До сих пор считалось, что в тот период (в начале так называемого африканского среднего каменного века, примерно соответствующего европейскому среднему палеолиту) в Африке жили представители только одной эволюционной линии поздних Homo – обладатели большого мозга, среди которых были и прямые предки современных людей. От этой линии ранее уже отделились и ушли в Евразию предки неандертальцев и денисовцев. Все прочие, более примитивные африканские гоминиды (австралопитеки, парантропы и ранние виды рода Homo) считались полностью вымершими к тому времени. Ну разве что в условиях островной изоляции где-то на краю света еще могли прозябать измельчавшие и деградировавшие потомки ранних эректусов – хоббиты. И вдруг выясняется, что такая точка зрения была лишь результатом неполноты палеонтологической летописи и ее недостаточной изученности. Теперь картина существенно усложнилась.
Оказывается, гоминиды оставались разнообразной группой не только в период диверсификации грацильных австралопитеков, парантропов и ранних Homo (примерно от 3 до 1,5 млн лет назад), но и значительно позже. В свете новых данных современная ситуация с единственным видом людей выглядит необычно. Homo sapiens фактически является реликтовым видом – последним уцелевшим представителем некогда разнообразной группы приматов. Новые данные окончательно поставили крест на старой идее о линейном антропогенезе – столбовой эволюционной дороге “от обезьяны к человеку”. Реальная эволюция гоминид имела мало общего с этой идеализированной схемой.
Палеонтологическая летопись африканских гоминид в промежутке от 700 до 200 тыс. лет назад вообще довольно скудна. Более того, из немногочисленных находок этого периода некоторые зубы и костные фрагменты, в принципе, могли бы принадлежать H. naledi или еще каким-нибудь примитивным гоминидам, хотя по умолчанию все находки этого возраста, в том числе фрагментарные и трудноопределимые, всегда относили к “нашей” эволюционной линии, то есть к поздним эректусам, “гейдельбергским людям в широком смысле” или архаичным сапиенсам. По-видимому, антропологам следует повнимательнее присмотреться к данному временному интервалу – вдруг там найдется еще что-то интересное.
Подводя итог сравнительному анализу морфологических признаков, Бергер и его коллеги заключили, что люди из пещеры Райзинг-Стар, скорее всего, представляют собой одну из ветвей ранней диверсификации рода Homo. В таком случае их последний общий предок с современными людьми жил, возможно, около 2 млн лет назад (вариант 1 на рис. 2.4), а продвинутые черты H. naledi – результат параллельной эволюции. Однако нельзя исключить и другие варианты (2 и 3 на рис. 2.4), предполагающие более близкое родство H. naledi с сапиенсами. В этом случае многочисленные примитивные черты H. naledi, включая маленький мозг, придется признать возвратом от продвинутого состояния к более примитивному (раньше такие эволюционные изменения называли “деградацией”, но сегодня так говорить невежливо).
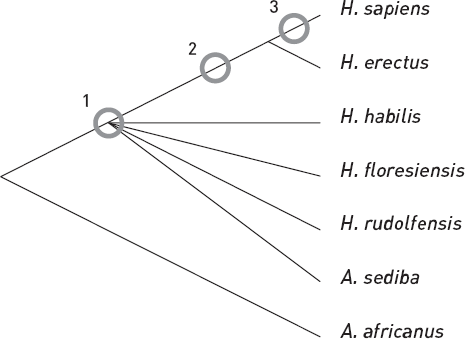
Рис. 2.4. Упрощенное филогенетическое дерево родов Australopithecus и Homo. Кружками показаны три возможных положения ветви, ведущей к H. naledi. По-видимому, наиболее вероятен вариант 1, хотя он предполагает независимое приобретение ряда продвинутых признаков черепа, стоп и кистей у H. naledi и у эректусов с сапиенсами. Из Berger et al., 2017.
Исследователи не исключают и возможность гибридного происхождения H. naledi. В этом нет ничего невероятного. Межвидовая гибридизация широко распространена у млекопитающих, в том числе у человекообразных обезьян. Например, разделившиеся два миллиона лет назад шимпанзе и бонобо впоследствии неоднократно скрещивались, как показал проведенный в 2016 году сравнительный анализ 65 полных геномов шимпанзе и 10 геномов бонобо. Из четырех подвидов шимпанзе наибольшая примесь генов бонобо (около 2,4 %) обнаружена у центрального подвида Pan troglodytes troglodytes, наименьшая – у западного подвида P. t. verus (De Manuel et al., 2016). По-видимому, эпизодическая гибридизация видов, разошедшихся сотни тысяч и даже миллионы лет назад, – не уникальная особенность рода Homo, а скорее общее правило для человекообразных.
Для формирования полной репродуктивной несовместимости между разделившимися видами млекопитающих, судя по всему, требуется около двух-четырех миллионов лет. Поэтому не исключено, что все плейстоценовые Homo могли скрещиваться друг с другом, а то и с австралопитеками. Судя по мозаичной морфологии H. naledi, этот вид мог бы быть гибридом между какими-нибудь продвинутыми Homo и поздними австралопитеками. Как проверить эту гипотезу, непонятно. Попытки извлечь ДНК из костей H. naledi пока не увенчались успехом.
По мнению исследователей, H. naledi должны были изготавливать каменные орудия. В пользу этого предположения свидетельствуют продвинутые черты строения их кистей и пальцев, сближающие их с неандертальцами и сапиенсами и отсутствующие у австралопитеков и хабилисов, а также маленькие зубы. Отсюда следует, что некоторые из орудий, до сих пор безоговорочно приписывавшихся африканским эректусам или архаичным сапиенсам, на самом деле могли быть изготовлены другими гоминидами.

