Книга: Эмоциональный мозг
Назад: Переключающая функция эмоций
Дальше: Компенсаторная (замещающая) функция эмоций
Подкрепляющая функция эмоций
Феномен подкрепления занимает центральное положение в системе понятий науки о высшей нервной деятельности, поскольку именно от факта подкрепления зависят образование, существование, угашение и особенности любого условного рефлекса. Под подкреплением «Павлов подразумевал действие биологически значимого раздражителя (пищи, вредоносного раздражителя и т. п.), которое придает сигнальное значение другому, сочетанному с ним несущественному в биологическом отношении раздражителю» [Асратян, 1971, с. 5]. Развивая представления И. П. Павлова, Э. А. Асратян полагает, что термин «подкрепляющий рефлекс» полнее и точнее отражает суть дела, чем «подкрепляющий раздражитель» (Там же, с. 6). Многочисленные факты показывают, что условный рефлекс можно выработать, сочетая и так называемые индифферентные раздражения, хотя скорость замыкания условной связи, ее стойкость и дальнейшая судьба в решающей мере зависят от видовых особенностей животного, интенсивности стимулов, их сенсорной модальности и порядка сочетаний [Schoenfeld, 1978]. Заметим, что с «индифферентностью» стимулов надо обращаться очень осторожно. Стимул, кажущийся безразличным для экспериментатора, может быть экологически важным для животного данного вида, не говоря уже о том, что исследовательская потребность (любопытство) чрезвычайно сильна у многих животных и делает «индифферентные» события «жизненно важными» [Berlyne, 1978].
Словно ощущая всю противоречивость и многозначность термина «подкрепление», И. П. Павлов в своих последних обобщающих работах (например, в статье «Условный рефлекс», написанной для «Большой советской энциклопедии») вообще не использует этот термин, предпочитая говорить только о совпадении индифферентного раздражителя с безусловным. В строгом смысле подкреплением следовало бы называть действие второго по порядку сочетания во времени стимула, благодаря которому первый стимул начинает вызывать ранее не свойственную ему реакцию. И все же биологическую значимость второго стимула нельзя сбрасывать со счетов, даже при моделировании условной связи путем прямого раздражения мозговых структур с целью изучения клеточно-синаптических механизмов замыкания. В опытах Л. Л. Воронина [1976] сенсорные стимулы (щелчок, свет) сочетались с прямым раздражением коры, вызывавшим движение соответствующей лапы. Присоединение к этой процедуре стимуляции эмоциональных структур гипоталамуса облегчало выработку условного рефлекса и делало его более стойким.
Необходимость вовлечения мозговых механизмов эмоций в процесс выработки условного рефлекса становится особенно демонстративной в случае инструментальных условных рефлексов, где подкрепление зависит от реакции субъекта на условный сигнал. Всесторонне проанализировав природу выработки инструментальных рефлексов, В. Вырвицка [Wyrwicka, 1975] пришла к выводу о том, что непосредственным подкреплением в этом случае является не удовлетворение какой-либо потребности, но получение желательных (приятных, эмоционально положительных) или устранение нежелательных (неприятных) стимулов. В зависимости от их интенсивности, функционального состояния организма и характеристик внешней среды приятными могут оказаться самые разнообразные «индифферентные» раздражители — световые, звуковые, тактильные, проприоцептивные, запаховые и т. п. С другой стороны, животные нередко отказываются от жизненно необходимых инградиентов пищи, если она невкусная. У крыс не удалось выработать инструментальный условный рефлекс при введении пищи через канюлю в желудок (то есть минуя вкусовые рецепторы), хотя такой рефлекс вырабатывается при введении в желудок морфина, который очень быстро вызывает у животного положительное эмоциональное состояние. Тот же морфин благодаря его горькому вкусу перестает быть подкреплением, если его вводить через рот [Cytawa, Trojiniar, 1976]. В другой серии опытов авторы вырабатывали инструментальный пищевой условный рефлекс у крыс, а после его упрочения заменяли натуральную пищу введением питательного раствора в желудок через носоглоточную канюлю. Рефлекс нажатия на рычаг при этом угасал, но сохранялся, если в желудок вводили 0,05 %-ный раствор морфина [Trojiniar, Cytawa, 1976].
Мы полагаем, что результаты этих опытов хорошо согласуются с данными Т. Н. Ониани [1975], который использовал прямое электрическое раздражение лимбических структур мозга в качестве подкрепления для выработки условного рефлекса. При сочетании внешнего стимула с раздражением структур мозга, вызывавшем у сытой кошки еду, питье, агрессию, ярость и страх, после 5-50 сочетаний удалось выработать только условную реакцию избегания, сопровождавшуюся страхом. Условных рефлексов еды и питья получить не удалось. Условнорефлекторный голод не удается получить и в натуральных условиях: обстановочные сигналы ситуации, в которой крыс заставляли голодать, вызывают у них не пищевое поведение, а страх и условную реакцию избегания [Могег, 1960]. Аналогичная реакция наблюдалась в опытах Т. Н. Ониани при подкреплении условного раздражителя стимуляцией «центров агрессии».
После 110 сочетаний звука и света с раздражением питьевой зоны гипоталамуса у коз условный сигнал не ведет к питьевому поведению, хотя подкрепляющая стимуляция сейчас же вызывает питье [Милнер, 1973]. С другой стороны, Э. Фонберг удалось выработать инструментальный условный рефлекс у собак, подкрепляя его стимуляцией, которая побуждала есть сытое животное. Автор полагает, что в этих опытах электрический ток активировал не структуры голода, а энграммы признаков вкусной пищи, что и заставляло собак продолжать еду, несмотря на состояние насыщения [Fonberg, 1967].
По мнению Т. Н. Ониани, подкреплением может быть раздражение только тех структур мозга, которые в естественных условиях активируются внешними факторами (страх), а не интероцептивными импульсами (голод, жажда). Возможность выработки условных рефлексов избегания, подкрепляемых стимуляцией «центров агрессии», Т. Н. Ониани объясняет тем обстоятельством, что формирование натуральных состояний агрессии имеет эндогенный компонент (гормональный при брачных боях, голодовой и т. п.).
С нашей точки зрения, результаты этих опытов еще раз свидетельствуют о решающей роли эмоций при выработке условных рефлексов. Страх имеет выраженную аверсивность для животного и активно минимизируется им путем реакции избегания. Раздражение пищевых и питьевых систем мозга у накормленных и не испытывающих жажды животных вызывает стереотипные акты еды и питья без вовлечения нервных механизмов эмоций, что исключает выработку условных рефлексов. Стимуляция центров агрессии генерирует при данной локализации электродов и параметрах тока эмоционально отрицательное состояние, которое так же, как в случае с эмоцией страха, ведет к минимизирующей реакции избегания. Если агрессивное поведение кошек сопровождается вовлечением эмоционально позитивных структур, то на базе их раздражения можно выработать условную реакцию самостимуляции, как это было показано А. В. Вальдманом, Э. Э. Звартау и М. М. Козловской [1976]. Эмоционально положительную окраску агрессивного поведения можно обнаружить и в натуральных условиях. Например, у мышей хорошо вырабатывается инструментальный условный рефлекс нажатия на рычаг, если это действие подкрепляется появлением в клетке другой мыши, немедленно атакуемой «исполнителем» [Connor, Watson, 1977].
Роль эмоций в замыкании условного рефлекса мы попытались схематически изобразить на рис. 6. С момента своего возникновения условно- рефлекторная теория предполагала конвергенцию двух возбуждений: от условного раздражителя и от стимула, вызывающего безусловный рефлекс, например — афферентацию из полости рта при поступлении в рот пищи (рис. 6, Г). Тогда же выяснилось значение «наличного функционального состояния», пищевой возбудимости, которую сегодня мы можем рассматривать как результат возбуждения мозговых структур, активизированных возникновением соответствующей потребности — состоянием голода (рис. 6, II). Однако ни афферентация из полости рта, ни голодовое возбуждение сами по себе не могут играть роль подкрепления, обеспечивающего формирование инструментального условного рефлекса. Только интеграция голодового возбуждения с возбуждением от фактора, способного удовлетворить данную потребность, то есть механизм, генерирующий положительную эмоцию, обеспечивает выработку условного рефлекса. При ином соотношении конвергирующих возбуждений, например, при поступлении пищи в рот перекормленного животного, активация механизмов отрицательной эмоции приведет к оборонительной реакции избегания (рис. 6, III).
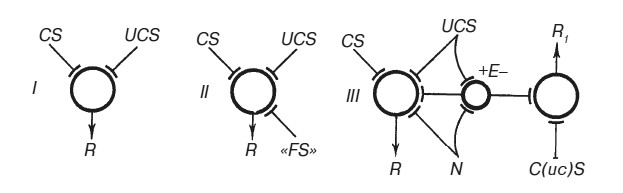
Рис. 6. Двух- (I), трех- (II) и четырехфакторные (III) схемы встречи возбуждений в пункте конвергенции при формировании условного рефлекса CS — условный стимул; UCS — безусловный стимул; R — реакция; FS — функциональное состояние; N — потребность; Е — эмоция.
Участие нервных механизмов эмоций в процессе выработки любого инструментального рефлекса делает весьма относительным выделение феномена так называемой эмоциональной памяти. По-видимому мы можем говорить только о большей или меньшей силе эмоциональной реакции на подкрепляющий стимул, как это имеет место при выработке условного рефлекса после одного сочетания или при выработке условной пищевой аверсии, когда подкрепление (отравление животного) происходит через несколько часов после восприятия условного сигнала. Что касается эмоций в момент воспроизведения условных связей, то они зависят от степени актуальности той потребности, на базе которой возникает данная эмоциональная реакция. Экспериментально показано, что дети 8 лет через 10 дней после первого опыта лучше воспроизводили тот словесный материал, который соответствовал мотиву, доминирующему в их личностной иерархии, будь то враждебность, лидерство, любознательность, привязанность и т. д. [Moore, Kagan, Haith, 1978].
Экспериментально установленная роль эмоций в процессе выработки классических и особенно — инструментальных условных рефлексов позволяет утверждать, что без учета и дальнейшей разработки нейрофизиологии эмоций мы вряд ли решим центральную проблему науки о высшей нервной деятельности: вопрос о механизмах замыкания условных связей.

