Книга: Эмоциональный мозг
Назад: Организация иерархии сосуществующих мотиваций — важная функция миндалевидного комплекса
Дальше: Взаимодействие мозговых структур, судя по пространственной организации их биоэлектрических процессов
Поведенческие функции гипоталамуса
Тесная связь функций гипоталамуса и миндалины продемонстрирована множеством экспериментов. По данным Т. Н. Ониани, эффекты стимуляции миндалины должны быть опосредованы через гипоталамус и другие структуры мезодиэнцефалона [Ониани, Мгалоблишвили, Чиджавадзе, 1978]. Миндалина оказывает регулирующее влияние на функции гипоталамуса, а в случае их выпадения компенсирует образовавшийся дефект. Эта компенсация происходит с участием структур новой коры. Так, после компенсации афагии у крыс, вызванной повреждением латерального гипоталамуса, корковая распространяющаяся депрессия вновь вызывает афагию, которая сохраняется в течение многих дней после декортикации. Повреждение ядер самой миндалины или полная нервная изоляция миндалевидного комплекса сопровождается менее выраженными поведенческими изменениями, чем аналогичные вмешательства на гипоталамусе.
В настоящее время имеется достаточно оснований рассматривать гипоталамус как сравнительно высокий уровень интегративной деятельности мозга. Очень трудно найти в гипоталамусе структуры, стимуляция которых вызвала бы вегетативные сдвиги без эмоциональных реакций. По-видимому, в гипоталамусе нет чисто «вегетативных центров», гипоталамус связан с организацией целостных поведенческих актов, в том числе — их вегетативных компонентов [Поляков, Талан, Черниговский, 1978]. Нейроны гипоталамуса очень быстро вовлекаются в условнорефлекторный ответ: активность нейронов латерального гипоталамуса изменялась через 150–200 мс после открывания заслонки для подачи пищи. Аналогичный ответ нейронов бледного шара наблюдался только через 300 мс, когда возникал двигательный компонент — изменения электромиограммы, связанные с лизанием [Rolls, Roper-Hall, Sanghera, 1977].
Реакция нейронов латерального гипоталамуса на вид и запах пищи зарегистрирована только у голодных обезьян, введение глюкозы устраняло этот эффект [Burton, Rolls, Mora, 1976]. Пачкообразная активность нейронов вентромедиального ядра гипоталамуса, возникающая у голодных животных, усиливается при поступлении пищи в рот и исчезает по мере наполнения желудка [Судаков, Журавлев, 1979]. Вместе с тем в гипоталамусе имеются и такие нейроны, которые будучи активированы голодом тормозятся сразу же с началом еды [Олдс, 1977]. При экспериментальном морфинизме у крыс нейроны латерального гипоталамуса активируются состоянием абстиненции (потребность) и тормозятся морфином (подкрепление). Нейроны медиального гипоталамуса, напротив, активируются подкреплением и тормозятся по мере актуализации потребности. Аналогичные данные получены для жажды и питья.
Многие авторы отмечают сравнительно узкую специализацию структур гипоталамуса. По данным Б. Оливьера, передний отдел медиального гипоталамуса у крыс контролирует пассивно-оборонительные реакции [Olivier, 1977]. Повреждение медиального гипоталамуса усиливает агрессивность крыс, вызванную болевым раздражением лап, и не влияет на территориальную агрессивность, которая нарушается только при разрушении латерального гипоталамуса [Adams, 1971]. Однако не меньшее количество фактов свидетельствует о зависимости последствий стимуляции и повреждения гипоталамуса от доминантного состояния животного и от стимулов окружающей среды [Isaacson, 1974].
Эффект стимуляции гипоталамуса одними и теми же параметрами тока при одной и той же локализации кончика электрода зависит от того, производится ли раздражение в пищевой или оборонительной ситуации опыта, у голодного или сытого животного [Беленков, Шалковская, 1978]. Стимуляция в области латерального гипоталамуса вызывает реакцию атаки на подчиненную крысу и не вызывает агрессивности по отношению к высокоранговому животному или самке [Koolhaas, 1978]. Под влиянием раздражения гипоталамуса макаки резусы нападают преимущественно на подчиненных самцов [Alexander, Parachio, 1973].
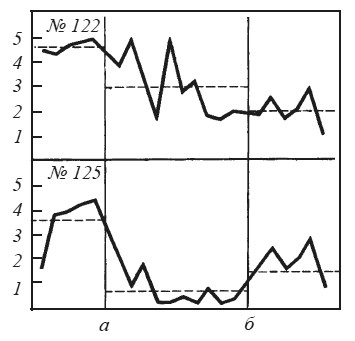
Рис. 35. Изменения реакции избегания у крыс № 122 и 125 после двустороннего повреждения латерального гипоталамуса Обозначения те же, что на рис. 34.
Для нас представляют особый интерес те случаи, где повреждение гипоталамуса дает эффект, противоположный амигдалэктомии. Так, после разрушений в гипоталамусе животные перестают реагировать на «тканевую жажду» и падение глюкозы в крови, но продолжают отвечать на условные сигналы воды и пищи [Олдс, 1977]. Их прошлый опыт приобретает известную самостоятельность, будучи изолирован от текущих нужд организма. Напомним, что при повреждениях миндалины нередко наблюдается прямо противоположное явление: например, после разрушения вентральной части медиального ядра у крыс нарушаются условные рефлексы, хотя потребление воды и пищи не претерпевает особых изменений [Korczynski, Fonberg, 1976]. Результат, противоположный последствиям амигдалэктомии, мы наблюдали и в опытах с реакцией избегания у крыс при болевом раздражении другой особи (рис. 35). Если амигдалэктомии нарушает баланс между конкурирующими мотивациями и выявляет доминирующую из них, то двустороннее повреждение латериального гипоталамуса, напротив, выравнивает силу мотиваций. Для этих крыс характерно «застревание» между педалью, включавшей ток, и открытым пространством камеры.
Итак, на уровне гипоталамуса конфликт между конкурирующими мотивациями однозначно решается в пользу одной из них за счет преобладания доминирующей в данный момент потребности. Участие миндалины делает этот процесс более пластичным, поскольку в конфликт вовлекаются эмоции, зависящие не только от силы потребностей, но и от вероятности их удовлетворения с учетом прошлого опыта и наличной ситуации. Благодаря миндалине возникает возможность сосуществования мотивов, их динамической иерархии. Эта возможность имеет огромное приспособительное значение: представим себе голодное животное, которое, стремясь к пище, прекращает наблюдение за сигналами потенциальной опасности. Вместе с тем функционирование миндалины способно осложнить поведение затрудненностью выбора между конкурирующими побуждениями. Подобно гиппокампу миндалину также можно назвать «органом колебаний и сомнений», но не в информационном, а в мотивационном смысле. Гиппокамп причастен к колебаниям, связанным с достижимостью целей, миндалина «взвешивает» их ценность.
Назад: Организация иерархии сосуществующих мотиваций — важная функция миндалевидного комплекса
Дальше: Взаимодействие мозговых структур, судя по пространственной организации их биоэлектрических процессов

