Книга: Эмоциональный мозг
Назад: Влияние эмоций на сердце
Дальше: Влияние эмоции на деятельность (на примере процессов восприятия)
Изменения электроэнцефалограммы при эмоциональных реакциях человека
Степень эмоционального напряжения нередко рассматривают как феномен, идентичный уровню бодрствования. Согласно этим представлениям эмоциональное возбуждение занимает в континууме бодрствования крайнее положение, диаметрально противоположное глубокому медленноволновому сну и коме. Подобный взгляд на природу эмоционального возбуждения, подкрепленный открытием функций ретикулярной формации мозга, получил свое наиболее законченное воплощение в «активационной теории эмоций» Дональда Б. Линдсли [1960]. Однако факты демонстрируют многочисленные примеры явной диссоциации между степенью эмоционального возбуждения, если о нем судить по сдвигам вегетативных функций, и уровнем бодрствования, диагностированным по суммарной электрической активности мозга. Ожидание болевых раздражений в опытах Р. Фроста с соавторами приводило к сдвигам частоты сердцебиений и электросопротивления кожи без достоверных изменений альфа-ритма электроэнцефалограммы — ЭЭГ [Frost, Burish, Holmes, 1978]. В состоянии страха, при восприятии эмоционально окрашенных слов, во время сдачи экзаменов были зарегистрированы не угнетение, а усиление альфа-ритма, повышение его амплитуды, возрастание альфа-индекса.
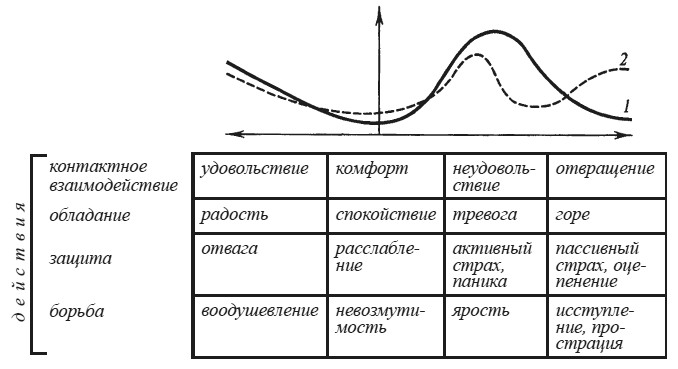
Рис. 14. Схема взаимодействия симпатического (1) и парасимпатического (2) отделов вегетативной нервной системы по мере роста эмоционального напряжения
Вопрос о соотношении вегетативных и ЭЭГ признаков эмоционального возбуждения был подвергнут систематическому анализу в экспериментах М. Н. Русаловой [1979]. На рис. 15 хорошо видно, что эмоциональное напряжение, о котором судили по учащению сердцебиений, может сопровождаться у одного и того же субъекта как угнетением, так и экзальтацией альфа-ритма. Чем различаются эти две ситуации? В первом случае субъект ожидал реальное болевое раздражение кожи предплечья. Во втором — ему была дана инструкция мысленно представить болевое раздражение в конце падающего отсчета, записанного на магнитофон.
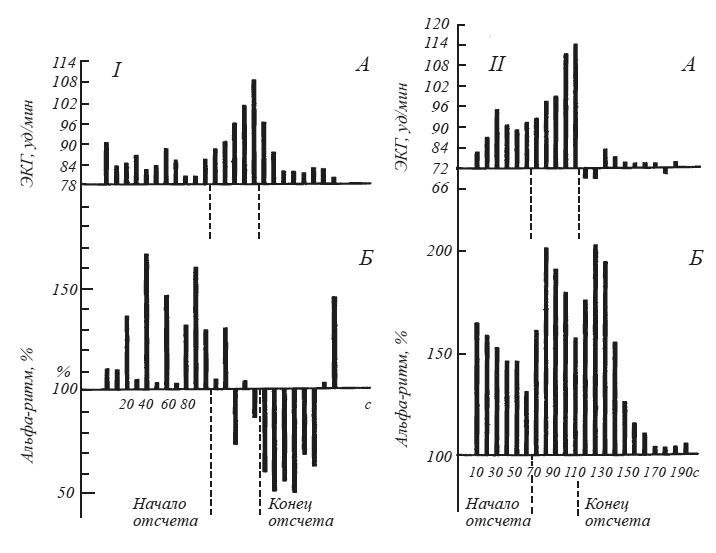
Рис. 15. Динамика изменений амплитуды (Б) альфа-ритма (по показанию пера интегратора) и частоты сердечного ритма (Д) после инструкции, в период отсчета и после отсчета
Альфа-ритм (11 Гц) выражен в процентах к фону, принятому за 100; I — действительное ожидание боли; II — воображаемое ожидание боли (по М. Н. Русаловой).
Выше мы говорили, что эмоциональная реакция может возникнуть как при поступлении информации из внешней среды, так и при извлечении следов ранее полученных впечатлений из памяти. Многочисленные эксперименты, поставленные М. Н. Русаловой (Валуевой), равно как и данные, почерпнутые в литературе, убеждают в том, что депрессия альфа-ритма характерна для ситуаций, в которых внимание субъекта обращено к окружающей его внешней среде. Если субъект преимущественно сосредоточен на следах, хранящихся в памяти, даже высокая степень эмоционального возбуждения не ведет к угнетению альфа-ритма, а зачастую сопровождается его экзальтацией. Подобный вывод хорошо согласуется с представлением об альфа-ритме как электрофизиологическом корреляте механизма, квантующего поток поступающей в мозг афферентации [Walter, 1954]. Нуну и Осселтон показали, что короткие зрительные стимулы не воспринимаются, если они подаются во время определенных фаз альфа-ритма. По мнению авторов, альфа-ритм коррелирует с деятельностью «нервной задвижки», которая периодически прерывает поступление информации в мозг [Nunn, Osselton, 1974].
Может возникнуть вопрос: почему же при сильном потоке афферентации, когда необходимость в работе «задвижки» особенно велика, альфа-ритм исчезает, депрессируется? Здесь не следует отождествлять деятельность механизма квантования с выраженностью его электрофизиологических коррелятов. Хорошо регистрируемый альфа-ритм при закрытых глазах, тишине и покое — это «холостой ход» квантующего механизма, который, кстати, существует и у лиц без выраженного альфа-ритма в ЭЭГ. Сказанное справедливо и для динамики тета-ритма, о котором сейчас пойдет речь.
Одним из ЭЭГ, симптомов эмоционального возбуждения служит усиление тета-ритма с частотой колебаний 4–7 Гц. С помощью анализаторов частотного спектра ЭЭГ тета-ритм был зарегистрирован у здоровых людей при отрицательных [Суворова, 1975] и положительных [Walter, 1953; Валуева, 1967] эмоциях. У детей, занятых деятельностью, окрашенной положительными эмоциями, тета-ритм лучше выражен в правом полушарии большого мозга, особенно в лобных его областях [Денисова, 1978]. Для понимания функционального значения тета-ритма интересны данные Брауна [Brown, 1971], который демонстрировал субъекту альфа-, бета- и тета-полосы его собственной ЭЭГ в виде различной цветовой окраски экрана. Испытуемых спрашивали, какому их субъективному состоянию соответствует тот или иной цвет. Оказалось, что для тета-ритма характерны обдумывание планов, неопределенность мысленно анализируемой ситуации, изменение темы размышлений, решение технических и житейских проблем, «сны наяву». Нарастание процентного содержания тета-ритма в спектре ЭЭГ совпадает со снижением бдительности человека-оператора к внешним сигналам [Beatty, Greenberg, Deibler, O’Hanlon, 1974].
Для того чтобы оценить функциональное значение изменений ЭЭГ при эмоциях, остановимся на гипотетическом механизме ритмических колебаний биопотенциалов мозга. Согласно гипотезе П. Андерсена и Дж. Экклса, важную роль в формировании этих колебаний играют тормозные нейроны [Andersen, Eccles, 1962]. Это дает основание применить к взаимодействию возбужденных и тормозящих их деятельность нервных микроструктур теорему В. Вольтерра [1976] о закономерных изменениях амплитуды и частоты осцилляций в системе, состоящей из двух типов функционально антагонистических элементов (табл. 3).
Таблица 3. Изменение электроэнцефалограммы, предсказываемое теоремой Вольтерра
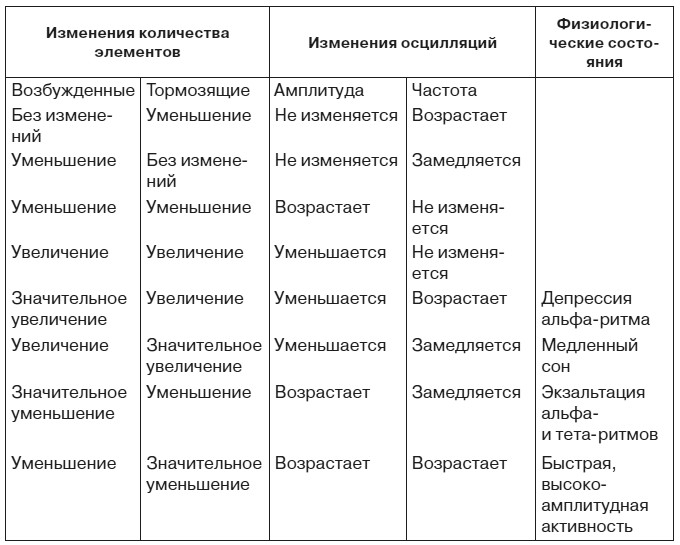
В какой мере картина, представленная в таблице, совпадает с физиологическими фактами? Наиболее бесспорен, пожалуй, механизм классической депрессии альфа-ритма, которая определяется нарастанием числа возбужденных и тормозящих элементов с преимущественным увеличением первых из них. Весьма неожиданным может показаться тот факт, что картина, характерная для медленного сна, также обусловлена увеличением возбужденных и тормозящих элементов, только число последних нарастает в большей мере, чем количество первых. Впрочем, данные последних лет не позволяют отвергнуть этот вывод. Усиление активности нейронов коры и ретикулярной формации во время сна побуждает рассматривать сон скорее как результат перестройки взаимодействия нервных элементов, чем как состояние «разлитого торможения» коры и подкорки в старом понимании этого термина. Ряд авторов придают важное значение нарастанию активности механизмов возвратного торможения в генезе медленного сна.
Высокоамплитудные медленные колебания свидетельствуют об уменьшении количества и возбужденных, и тормозящих элементов с относительным преобладанием последних. Уменьшение количества возбужденных элементов, но с преимущественным уменьшением тормозящих микроструктур имеет место и при быстрой высокоамплитудной активности. Применение теоремы Вольтерра к ритмической биоэлектрической активности мозговых структур не только создает предпосылки для математического моделирования ЭЭГ, но и побуждает по-новому взглянуть на внутренний нейрональный механизм ряда типичных изменений электроэнцефалограммы.
Если в генезе альфа-ритма решающее значение имеют кортикоталамические отношения, то тета-ритм связан с кортико-лимбическим взаимодействием. Появление тета-ритма в ЭЭГ человека, находящегося в коматозном состоянии, можно рассматривать как результат активности филогенетически древних лимбических структур на фоне деактивации менее устойчивых неокортикальных образований [Гриндель, Брагина, Добронравова, Доброхотова, 1974].
Итак, альфа-ритм — это «сенсорный» ритм, связанный с поступлением информации из внешнего мира. Не случайно он лучше всего выражен в зрительных отделах новой коры, поскольку у человека зрение является основным каналом получения информации об окружающей внешней среде. «Лимбический» по своему происхождению «эмоциональный» тета-ритм, по-видимому, имеет какое-то отношение к деятельности «оценивающих» механизмов мозга. С таким предположением хорошо согласуются временные параметры альфа- и тета-ритмов. Согласно данным психофизики восприятия первые 100 мс необходимы для отражения в структурах мозга физических характеристик стимула, для формирования его «нервной модели». На протяжении следующих 100 мс мозг производит сопоставление модели с энграммами, извлеченными из памяти. Между 200–300 мс выясняется результат сличения и начинается подготовка к реакции, если наличный стимул требует ответного действия [Ломов, Иваницкий, 1977]. Продолжительность первого этапа совпадает с временными характеристиками альфа-ритма. Продолжительность процессов сличения и «принятия решения» (150–250 мс) идентична частоте тета-ритма (рис. 16).
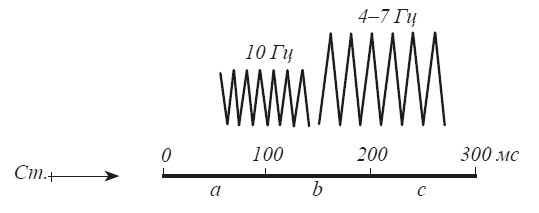
Рис. 16. Схема сопоставления временных характеристик альфа- и тета-ритма ЭЭГ с процессом восприятия внешнего стимула (Ст.)
а — формирование нервной модели; b — сравнение модели с энграммами, извлеченными из памяти; с — начало формирования реакции в зависимости от результатов сравнения.
Мы полагаем, что усиление тета-ритма при эмоциях не сводится к простой активации неокортекса со стороны лимбических структур. Характерная для эмоций ситуация прагматической неопределенности требует усиленной мобилизации хранящихся в памяти энграмм, их сопоставления с сигналами, поступающими из внешней среды. Весьма вероятно, что тета-ритм является электрофизиологическим коррелятом механизма, квантующего поток извлекаемых из памяти энграмм, подобно тому, как альфа-ритм связан с механизмом, квантующим поток информации из внешней среды. Подобное предположение хорошо объясняет, почему тета-ритм так характерен для электрической активности гиппокампа у многих видов животных, о чем специально мы будем говорить в следующей главе.
Сказанное выше не означает, что каждый раз, регистрируя тета-ритм, мы наблюдаем процесс извлечения энграмм: у роженицы во время родов, у коматозного больного и т. д. Особенно это относится к случаям патологии. Когда у автомашины повреждено рулевое управление и нет двух колес, продолжающий работать двигатель уже не является источником перемещения в пространстве. С другой стороны, поступление внешней информации и актуализация, извлечение имеющейся представляют два фундаментальных процесса функционирования любой нервной сети. Вот почему мы вправе считать, что наличие альфа- и тета-ритмов во многих мозговых образованиях означает не только сходный уровень их возбуждения, но и указывает на сходство протекающих в них рабочих процессов. О последнем свидетельствует функциональное значение пространственной синхронизации по тета-ритму, установленное в лаборатории М. Н. Ливанова [1972].
Информационная теория, вскрывшая сложную внутреннюю структуру эмоциональных состояний, наличие в этой структуре потребностно-мотивационных и информационных компонентов, позволяет более адекватно анализировать изменения ЭЭГ при эмоциях, не отождествляя эти изменения со шкалой колебаний уровня бодрствования. Если первоначально эти колебания объясняли активирующим влиянием одной лишь ретикулярной формации мозгового ствола, то постепенно число активирующих (а равно и деактивирующих) систем стало нарастать. Сложнейшую конструкцию и взаимодействие активирующих систем уже невозможно сопоставлять с «целостным», далее не расчлененным представлением об эмоции, все еще кочующим по страницам учебников и руководств.
Назад: Влияние эмоций на сердце
Дальше: Влияние эмоции на деятельность (на примере процессов восприятия)

