Книга: Река, выходящая из Эдема. Жизнь с точки зрения дарвиниста
Назад: Глава 2 Вся Африка и потомки ее
Дальше: Глава 4 Божественная функция полезности
Глава 3
Делай добро украдкой
Креационизм обладает неодолимой притягательностью, и за объяснением, почему это так, далеко ходить не надо. Дело тут — по крайней мере, в большинстве известных мне случаев — отнюдь не в приверженности ни к буквальному содержанию Книги Бытия, ни к легенде о происхождении мира еще какого-нибудь племени. Люди просто открывают для себя всю красоту и сложность мира живых существ, приходя к выводу, что он, «очевидно», был кем-то спроектирован. Те из креационистов, кто признает, что эволюция по Дарвину может хотя бы в некоторой степени служить альтернативой их священным теориям, зачастую прибегают к чуть более сложному доводу: отрицают возможность существования промежуточных форм. «X непременно был создан Творцом, — говорят они, — потому что половинка от X вообще не смогла бы работать. Все составные части X должны были оказаться на своих местах одновременно — они не могли эволюционировать постепенно». Вот пример: в тот самый день, когда я начал писать эту главу, мне пришло письмо от одного американского священника, который раньше был атеистом, но уверовал после того, как прочел статью в журнале National Geographic. Приведу отрывок из его письма:
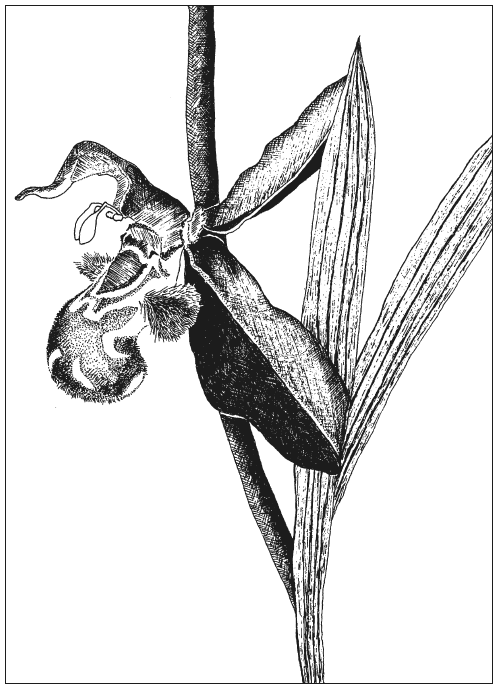
Речь в той статье шла о поразительных приспособлениях к окружающей среде, выработанных орхидными для успешного размножения. По мере чтения меня особенно заинтересовала стратегия одного вида, для которой требовалось участие самца осы. Внешне цветок очень напоминал самку того же вида ос, в том числе имел отверстие в подходящем месте — так чтобы самец, просто совокупившись с цветком, мог дотянуться до производимой тем пыльцы. При перелете на другой цветок этот процесс повторялся, благодаря чему происходило перекрестное опыление. Но в первую очередь цветок делало привлекательным для ос то, что он выделял феромоны [так называют особые химические аттрактанты, широко используемые насекомыми для привлечения половых партнеров — Р. Д.], идентичные феромонам данного осиного вида. Около минуты я с любопытством разглядывал сопутствующую иллюстрацию. Затем с чувством глубочайшего потрясения осознал: для того чтобы такая репродуктивная стратегия вообще могла работать, она изначально должна была быть совершенной. Ее невозможно объяснить никакими постепенными преобразованиями, ведь если бы орхидея не выглядела и не пахла, как самка осы, если бы она не обладала пригодным для копуляции отверстием с пыльцой, расположенной в пределах досягаемости полового органа самца, вся стратегия потерпела бы полный провал.
Никогда не забуду чувства беспомощности, охватившего меня, поскольку в тот момент мне стало ясно, что некий Бог должен в какой-то форме существовать и находиться в непрерывной связи с любыми процессами становления чего бы то ни было. Что, проще говоря, Бог-творец — это не какой-нибудь допотопный миф, а нечто реальное. И также, с крайней неохотой, я внезапно осознал, что должен больше разузнать об этом Боге.
Несомненно, другие приходят к религии иначе, но наверняка многие прошли через опыт, сходный с тем, что изменил жизнь этого священника (чью личность я не раскрываю из соображений благопристойности). Им довелось либо увидеть какое-то из чудес природы, либо прочесть о нем. Если говорить в общем, то это наполнило их благоговейным трепетом, перешедшим в преклонение. Если же конкретнее, то они, подобно моему корреспонденту, решили, что данное явление природы — будь то паучья сеть, или орлиный глаз, или орлиное же крыло, или что угодно другое — не могло возникнуть в результате поэтапной эволюции, так как промежуточные, недооформившиеся стадии были бы ни на что не пригодны. Цель настоящей главы — опровергнуть этот довод: будто сложным приспособлениям, чтобы хоть как-то функционировать, необходимо быть совершенными. Кстати говоря, орхидные были одним из любимых примеров Чарльза Дарвина, посвятившего целую книгу тому, чтобы показать, с каким триумфом принцип плавной эволюции путем естественного отбора проходит суровое испытание объяснением «различных приспособлений, при помощи которых орхидеи опыляются насекомыми».
Суть аргументации нашего священника заключена в утверждении, что «для того чтобы такая репродуктивная стратегия вообще могла работать, она изначально должна была быть совершенной, ее невозможно объяснить никакими постепенными преобразованиями». Тот же довод можно было бы также высказать — и многократно высказывали — насчет эволюции глаза, к чему я в этой главе еще вернусь.
Всякий раз, когда я слышу заявления подобного рода, меня неизменно поражает их безапелляционность. Как, хочу я спросить у написавшего мне священника, можете вы быть настолько уверены в том, что цветок, подражающий осе (или глаз, или еще какой-либо орган), не смог бы функционировать, если бы все его части не были совершенными и идеально подогнанными друг к другу? Задумывались ли вы вообще хотя бы на долю секунды, о чем говорите? Обладаете ли хотя бы начальными знаниями об орхидеях, или об осах, или о глазах, которыми осы смотрят на самок и на орхидеи? Что дало вам право утверждать, будто осы настолько проницательны, что одурачить их можно только сходством, идеальным во всех отношениях?
Припомните, когда в последний раз вы сами были обмануты случайным сходством. Вам, вероятно, приходилось снимать шляпу на улице перед незнакомкой, которую вы приняли за известную вам личность. В кинематографе используют каскадеров, чтобы те падали с лошадей и прыгали со скал вместо актеров и актрис. Сходство каскадера с подменяемой им кинозвездой обычно крайне поверхностно, однако при быстрой смене кадров этого достаточно, чтобы обмануть зрителей. Человеческие самцы испытывают вожделение при виде картинки из журнала. А ведь это всего лишь чернила на бумаге. Картинка двух-, а не трехмерна. Величина ее не более нескольких дюймов. Она может быть не правдоподобным изображением, а грубым наброском из нескольких линий — и все равно вызывать у мужчины эрекцию. Возможно, мимолетный силуэт самки — это все, что оса, летящая на большой скорости, ожидает увидеть, перед тем как приступить к копуляции. Возможно, самцы ос вообще обращают внимание только на некоторые ключевые стимулы.
Имеются все основания полагать, что осу еще проще ввести в заблуждение, чем человека. Колюшку определенно проще, а ведь у рыб мозг крупнее и глаза лучше, чем у ос. У самцов колюшки красное брюшко, и они ведут себя агрессивно по отношению не только к другим самцам, но и к грубо сработанным макетам с красным «брюшком». Мой старый учитель Нико Тинберген, этолог и нобелевский лауреат, рассказывал знаменитый случай, как все самцы колюшки в аквариумах бросились с угрожающим видом в сторону окна лаборатории, когда мимо проезжал красный почтовый фургон. Брюшко самки колюшки, готовой метать икру, заметно раздувается. Тинберген обнаружил, что чрезвычайно грубые, слегка продолговатые серебристые подделки, не имеющие, на наш с вами взгляд, ничего общего с колюшкой, но обладающие округлым «брюшком», провоцируют полноценное брачное поведение со стороны самцов. Более поздние эксперименты исследователей, принадлежащих к основанной Тинбергеном научной школе, показали, что так называемая секс-бомба — штуковина грушевидной формы, воплощенная пузатость, но не продолговатая и совершенно не похожая на рыбу, как ни напрягай (человеческое) воображение, — пробуждает похоть в колюшках мужского пола даже еще эффективнее. Эта «секс-бомба» представляет собой классический пример сверхнормального стимула — то есть такого, который действует сильнее, чем подлинный. В качестве другого примера Тинбергеном было опубликовано фото кулика-сороки, сидящего на яйце размером со страусовое. Мозг у птиц больше, а зрение лучше, чем у рыб (и уж тем более чем у ос), и однако же кулику-сороке явно «кажется», будто огромное, как у страуса, яйцо — превосходный объект для высиживания.
У чаек, гусей и прочих гнездящихся на земле птиц имеется стандартная реакция на яйцо, выкатившееся из гнезда. Они вытягивают шею и закатывают его назад нижней частью клюва. Тинберген и его ученики выяснили, что чайки поступают так не только со своими собственными яйцами, но и с куриными и даже с деревянными цилиндрами, а также с оставшимися после туристов жестянками из-под какао. Птенцы серебристой чайки получают пищу, выпрашивая ее у родителей: они тычутся в красное пятно на родительском клюве, чем стимулируют родителя отрыгнуть немного рыбы из набитого зоба. Тинберген совместно с одним коллегой показал, что примитивная картонная модель родительской головы успешно провоцирует выпрашивающее поведение у птенцов. Все, что действительно необходимо, — это красное пятно. С точки зрения птенца серебристой чайки красное пятно и есть родитель. Вполне возможно, что птенец видит и остальные части родительского тела, но, судя по всему, значения это не имеет.
Столь явно ограниченный взгляд на вещи присущ не только чаячьим птенцам. Взрослых озерных чаек можно отличить по их характерным темным «маскам». Чтобы изучить важность этого признака для других взрослых особей, студент Тинбергена Роберт Мэш изготавливал и раскрашивал искусственные чаячьи головы из дерева. Каждая голова закреплялась на конце деревянной палки, которую Мэш подсоединял к находившемуся внутри коробки электромотору таким образом, чтобы при помощи дистанционного управления передвигать эту голову вверх и вниз, а также поворачивать ее вправо и влево. Коробку он зарывал поблизости от гнезда чайки, так чтобы деревянная голова оказывалась вне поля зрения, под песком. Затем он изо дня в день приходил в укрытие возле гнезда и наблюдал реакцию находившихся в гнезде чаек на муляж головы, когда тот поднимался из песка и поворачивался в ту или иную сторону. Птицы реагировали так, как если бы имели дело с настоящей чайкой, хотя это была не более чем подделка без туловища, ног, крыльев и хвоста, не издававшая никаких звуков и не совершавшая никаких движений, за исключением откровенно безжизненных, роботоподобных поднятия, вращения и опускания. По-видимому, в восприятии озерной чайки подозрительный сосед — это едва ли многим более, чем бестелесная черная маска. Ни в туловище, ни в крыльях, ни в чем-либо еще необходимости, судя по всему, нет.
Да просто для того, чтобы попасть в свое укрытие, Мэш, как и многие поколения орнитологов до и после него, пользовался давно известным изъяном птичьей нервной системы: птицы не слишком сильны в математике. В укрытие входят двое, а выходит только один. Без этой уловки птицы были бы настороже, поскольку «знали» бы, что там кто-то есть. Но, увидев, что один человек ушел, они «полагают», будто ушли все. Раз птица не в состоянии заметить разницу между одним человеком и двумя, так ли уж будет удивительно, если самец осы окажется обманут орхидеей, чье сходство с самкой не вполне абсолютно?
Еще одна птичья история в том же ключе, на сей раз трагедия. Индюшки — отважные защитницы своих птенцов. Им приходится оборонять их от таких расхитителей гнезд, как ласки и крысы. Золотое правило индюшачьей матери, при помощи которого она распознает грабителей, пугающе грубое: нападай на любой движущийся поблизости от твоего гнезда объект, если он не кричит, как индюшонок. Это открытие сделал австрийский зоолог по имени Вольфганг Шляйдт. Когда-то у него была индюшка, которая беспощадно убивала всех своих птенцов. Происходило это по прискорбно простой причине: она была глухой. Индюшачьей нервной системой хищник определяется как движущийся объект, не кричащий подобно птенцу. Несчастные индюшата, несмотря на то что они выглядели, как индюшата, двигались, как индюшата, и доверчиво бежали к своей маме, как индюшата, пали жертвой ее чрезмерно узкой трактовки понятия «хищник». Она защищала своих детей от самих себя и уничтожала их всех.
С трагической историей индюшки перекликается и следующий факт из жизни насекомых. Некоторые из чувствительных клеток на усиках медоносной пчелы реагируют только на одно вещество — олеиновую кислоту. (У пчел есть и другие чувствительные клетки, восприимчивые к другим соединениям.) Олеиновая кислота выделяется пчелиным организмом при разложении и включает у пчел «погребальное поведение» — удаление мертвых сородичей из улья. Если экспериментатор нанесет немного олеиновой кислоты на живую пчелу, то несчастное брыкающееся, упирающееся и, вне всяких сомнений, живое создание будет выброшено вместе с покойниками.
Мозг у насекомых намного меньше, чем у индейки или человека. Глаза насекомых, даже крупные фасеточные у стрекоз, обеспечивают лишь малую долю той зоркости, какая свойственна нашим с вами глазам и глазам птиц. Известно также и то, что насекомые видят мир совершенно иначе, чем мы. Великий австрийский зоолог Карл фон Фриш открыл в молодости, что они неспособны видеть красный свет, но видят — и воспринимают как отдельный оттенок — ультрафиолетовый свет, который неспособны видеть мы. Глаза насекомых придают большое значение так называемому мельканию, которое, судя по всему (по крайней мере, у быстро передвигающихся насекомых), отчасти заменяет им то, что мы воспринимаем как «форму». Было замечено, как самцы бабочек «ухаживают» за порхающими на ветру опавшими листьями. Мы видим самку бабочки как два больших крыла, машущих вверх-вниз. Самец же, находясь в полете, воспринимает ее как некое сосредоточение «мелькания». Вы можете обмануть его при помощи стробоскопической лампы, которая вообще не двигается, а только мигает. Если подобрать правильную частоту вспышек, он примет лампу за другую бабочку, бьющую крыльями с этой частотой. Для нас с вами полосы — это неподвижный рисунок. А летящему насекомому они видятся как «мелькание» и могут быть сымитированы стробоскопом, мигающим с нужной частотой. Мир, воспринимаемый глазами насекомых, настолько чужд нам, что любые заявления, делаемые на основании собственного опыта, насчет того, насколько «совершенно» орхидея должна подражать самке осы, — это не более чем человеческая самонадеянность.
Осы тоже стали объектом одного классического опыта, впервые проделанного великим французским натуралистом Жаном-Анри Фабром и многократно воспроизводившегося другими исследователями, в том числе и из школы Тинбергена. Самка роющей осы возвращается в свою норку, принося туда ужаленную и парализованную жертву. Прежде чем затащить добычу в норку, она оставляет ее у входа и залезает внутрь — по-видимому, чтобы проверить, все ли в порядке. Во время ее отсутствия экспериментатор отодвигает добычу на несколько дюймов в сторону. Вылезая на поверхность, оса видит, что жертва пропала, но быстро ее находит. Затем она подтаскивает добычу обратно ко входу в свою норку. С тех пор, как она проверила, все ли там в порядке, прошло лишь несколько секунд. Кажется, у осы нет никакой реальной причины, чтобы не перейти к следующему этапу своей привычной процедуры: затащить добычу в норку и покончить с этим делом. Однако ее программа сброшена на предыдущую команду. Подчиняясь, оса послушно оставляет добычу возле норки и забирается внутрь, чтобы снова все проинспектировать. Экспериментатор волен повторять свою проделку хоть сорок раз, пока ему не надоест. Оса ведет себя подобно стиральной машине, чью программу перевели на предыдущую стадию и которая не «знает», что уже постирала это белье сорок раз подряд. Для обозначения такого упрямого, бездумного поведения выдающийся информатик Дуглас Хофштадтер предложил новое прилагательное — «сфекский». (Sphex — название одного типичного рода роющих ос.) Следовательно, по крайней мере в некоторых отношениях осы легко поддаются обману. Это совсем не тот обман, каким пользуются орхидеи. Но тем не менее следует остерегаться человеческой интуиции, подталкивающей нас к выводу, будто «для того чтобы такая репродуктивная стратегия вообще могла работать, она изначально должна была быть совершенной».
Быть может, я перестарался, убеждая вас в том, что одурачить осу, по всей видимости, нетрудно. В вас могло зародиться подозрение, почти противоположное тому, какое испытывал мой преподобный корреспондент. Если зрение у насекомых такое слабое, а осы — и впрямь простофили, зачем тогда орхидее утруждать себя, делая свой цветок настолько похожим на осу? Ну, вообще-то зрение у ос не всегда так уж ни на что не годится. Бывают ситуации, в которых, судя по всему, осы видят вполне неплохо: скажем, когда они определяют местонахождение своей норки после продолжительной ночной охоты. Тинберген изучал это на примере роющей осы Philanthus, называемой также пчелиным волком. Он ждал, когда оса заберется к себе в норку, и поспешно, пока она не вылезла, размещал у самого входа какие-нибудь «ориентиры» — допустим, веточку и шишку. Затем отходил в сторону и ждал, когда оса вылетит. Дождавшись, он видел, как она делала в воздухе пару-тройку кругов, будто бы мысленно фотографируя местность, после чего улетала за добычей. Во время ее отсутствия Тинберген передвигал веточку и шишку на несколько футов. Когда оса возвращалась, она не находила свою норку, а вместо этого пикировала в песок — в точку, расположенную относительно веточки и шишки точно так же, как прежде вход. Да, в каком-то смысле осу опять «одурачили», но на сей раз мы вынуждены отдать должное ее зрению. Похоже, она и в самом деле «мысленно фотографировала местность», когда кружила над норкой, перед тем как улететь. По-видимому, она распознала образ, или «гештальт», веточки с шишкой. Тинберген многократно повторял этот опыт, используя различные ориентиры — например, гирлянды из шишек, — и всегда с неизменным результатом.
А вот эксперимент, который провел ученик Тинбергена Герард Берендс и результат которого разительно отличается от фабровской «стиральной машины». Объектом для Берендса служил вид роющих ос (изучавшийся также и Фабром) Ammophila campestris. Эта оса необычна тем, что является «регулярным снабженцем». По большей части роющие осы заполняют свою норку провизией, откладывают яйцо, а затем запечатывают вход, предоставляя юной личинке питаться самостоятельно. Ammophila же поступает иначе. Она, подобно птице, ежедневно возвращается к норке, проверяя, все ли у личинки в порядке, и снабжая ту пищей по мере необходимости. Все это само по себе не так уж примечательно. Однако каждая самка Ammophila всегда занимается двумя-тремя норками одновременно. В одной из них находится относительно крупная, почти выросшая личинка, в другой — маленькая, недавно отложенная, а в третьей, допустим, личинка промежуточного возраста и средних размеров. Мать ухаживает за ними сообразно их потребностям в пище, которые, естественно, различаются. В серии трудоемких экспериментов, включавших в себя и обмен содержимым между норками, Берендс сумел показать, что осы-матери действительно учитывают индивидуальные пищевые потребности каждого гнезда. Это выглядит разумным, но Берендс обнаружил, что это вместе с тем и неразумно, причем неразумно в очень странной, инопланетной манере. Каждое утро осиная мать первым делом обходит с инспекцией все свои действующие норки. Интересует ее исключительно состояние каждого гнезда на момент утреннего обхода, и именно этим состоянием определяется ее запасающее поведение на весь оставшийся день. После утренней инспекции Берендс мог менять содержимое гнезд местами столько раз, сколько ему заблагорассудится, — на запасающем поведении мамаши-осы это никак не отражалось. Она как будто бы включала свой гнездооценивающий прибор только во время утреннего обхода, а на остаток дня выключала, экономя электричество.
С одной стороны, из этой истории следует, что в голове у осы имеется сложное оборудование для подсчетов, измерений и даже вычислений. Теперь в самом деле становится нетрудно поверить в то, что обмануть осиный мозг можно только тщательно выверенным сходством цветка орхидеи с самкой осы. Но, с другой стороны, опыт Берендса указывает на наличие у ос как некой склонности к избирательной слепоте, так и некоторой недальновидности — вполне в духе эксперимента со «стиральной машиной», — позволяющих поверить и в то, что самого грубого сходства между орхидеей и самкой будет вполне достаточно. Главный урок, который нам следует извлечь из всего этого, таков: никогда нельзя подходить к оценке подобных явлений с человеческой меркой. Никогда не говорите сами и не воспринимайте всерьез того, кто говорит: «Не могу поверить, будто то-то и то-то возникло путем постепенного отбора». Этой разновидности заблуждения я дал название «убеждение личным недоверием». Из раза в раз обращение к такой аргументации оказывается на поверку интеллектуальным аналогом наступания на банановую кожуру.
Вот довод, который я подвергаю здесь критике: «Плавная эволюция того-то и того-то не могла иметь места, поскольку „очевидно“, что то-то и то-то, чтобы вообще как-то функционировать, должно быть совершенным и целостным». До сих пор в своих возражениях я напирал на тот факт, что осы и прочие животные видят мир совершенно иначе, чем мы, и что в любом случае даже нас с вами нетрудно одурачить. Но теперь я собираюсь изложить и другие аргументы, даже еще более убедительные и более общие. Давайте по отношению к таким устройствам, которым, чтобы хоть как-то работать, необходимо быть совершенными, будем использовать термин «хрупкий». Таковы, по мнению моего корреспондента, подражающие осам орхидеи. На самом деле подыскать безоговорочно хрупкий механизм довольно трудно, и я нахожу это показательным. Самолет не хрупок, потому что, хотя мы и предпочли бы доверить свои жизни такому «Боингу-747», все мириады составных частей которого работают абсолютно исправно, в действительности, даже лишившись наиважнейших деталей своей оснастки — например, одного или двух двигателей, — он все равно будет лететь. Микроскоп не хрупок, поскольку даже в плохонький, дающий расплывчатое и тусклое изображение микроскоп разглядывать маленькие объекты все же лучше, чем вовсе без него. Не является хрупким и радио: в каком-то смысле оно не лишено недостатков, так как качество сигнала может падать, а звуки — становиться металлическими и искаженными, но вы все равно сумеете разобрать смысл передаваемых слов. Я десять минут стоял, уставившись в окно, пытаясь подобрать хотя бы один действительно удачный пример хрупкого приспособления, созданного людьми, и единственное, что пришло мне в голову, — это арка. Она определенно близка к тому, чтобы называться хрупкой в этом смысле: когда две ее стороны уже соединены, арка обладает большой прочностью и устойчивостью, но пока стороны не соединены, ни одна из них не будет стоять вообще. Чтобы построить арку, нужно использовать какие-то подмости, обеспечивающие временную поддержку до тех пор, пока вся конструкция не будет готова. Затем их можно убрать, а арка останется крепко стоять на долгие времена.
В человеческих технологиях нет ничего, что в принципе не позволяло бы устройству быть хрупким. Инженеры вольны проектировать на своих чертежных досках такие механизмы, которые, не будучи полностью готовыми, не смогут работать вообще. И тем не менее даже в области технологий найти подлинно хрупкий аппарат не так-то просто. Думаю, что по отношению к живым аппаратам это справедливо в еще большей степени. Давайте посмотрим на некоторые якобы хрупкие приборы из мира живой природы, которые креационистская пропаганда так любит использовать в качестве примеров. История с орхидеей и осой — только частный случай восхитительного явления мимикрии. Многие животные и некоторые растения получают преимущество благодаря своему сходству с другими объектами — зачастую с другими животными или растениями. Мимикрия может как положительно, так и отрицательно влиять практически на любой аспект жизнедеятельности организмов: добывание пищи (тигры и леопарды, подкрадывающиеся к своим жертвам, почти что невидимы в испещренном солнечным светом лесу; рыба-удильщик похожа на морское дно, лежа на котором она привлекает добычу при помощи длинной «удочки» с приманкой, напоминающей червяка; самки светляков, эти «роковые женщины», имитируют световые брачные сигналы других видов, заманивая таким образом самцов, чтобы съесть их; аспидонт подражает другим видам рыб, являющихся чистильщиками для более крупной рыбы, и, получив привилегированный доступ к своим «клиентам», отъедает куски от их плавников); избегание опасности быть съеденным (животные, рискующие стать добычей хищников, на различный лад прикидываются древесной корой, прутиками, сочными зелеными или сухими опавшими листьями, цветками, шипами роз, слоевищами водорослей, камнями, птичьими экскрементами, а также другими животными, заведомо ядовитыми или опасными); отвлечение хищников от молоди (шилоклювки и многие другие птицы, гнездящиеся на земле, своими позами и походкой изображают, будто у них сломано крыло); обеспечение заботы о будущем потомстве (яйца кукушек воспроизводят специфическую окраску яиц вида-хозяина; на боках у самок некоторых видов рыб, вынашивающих икру во рту, есть узор, напоминающий икринки и служащий для привлечения самцов, чтобы те набрали себе в пасть настоящую икру и заботились о ней).
Вне зависимости от примера, всегда есть искушение полагать, будто мимикрия не сможет быть эффективной, не будучи идеальной. В конкретном случае с похожей на осу орхидеей я делал основной акцент на свойственных осам и другим жертвам мимикрии несовершенствах восприятия. Если говорить начистоту, то, на мой взгляд, сходство орхидей с изображаемыми ими осами, пчелами или мухами отнюдь не выглядит таким уж впечатляющим. Сходство листовидки с листом кажется моим глазам намного более точным — возможно, потому, что мои глаза больше похожи на глаза тех хищников (предположительно, птиц), против которых данная мимикрия направлена.
Но есть и более общее соображение, почему мнение, будто мимикрии, чтобы вообще иметь смысл, необходимо быть совершенной, неверно. Сколь бы острым ни было зрение, скажем, какого-нибудь хищника, условия видимости далеко не всегда идеальны. Причем практически неизбежно должен существовать некий непрерывный спектр этих условий: от наихудшей видимости к наилучшей. Подумайте о каком-нибудь действительно хорошо знакомом вам предмете — таком, который вы ни за что не перепутаете ни с каким другим. Или о каком-нибудь человеке — например о близкой подруге, которую вы так знаете и любите, что никогда в жизни не примете за кого-то еще. Ну а теперь представьте себе, что этот человек приближается к вам с дальнего расстояния. Непременно должны существовать как дистанция, с которой вы вообще не будете его видеть, так и дистанция, с которой вам будут видны каждая черта, каждая ресница, каждая пора. И на промежуточных расстояниях никакого внезапного перехода от одного к другому не происходит: имеет место плавное увеличение или уменьшение узнаваемости. В военных пособиях по стрельбе об этом говорится открытым текстом: «За двести ярдов все части тела отчетливо видны. За триста ярдов контуры лица становятся расплывчатыми. За четыреста ярдов лица не видно. Начиная от шестисот ярдов голова представляет собой точку, а очертания тела сходят на нет. Вопросы есть?» Конечно, постепенно приближающуюся подругу вы можете узнать и сразу. Но в этом случае дистанция создает градиент вероятности мгновенного узнавания.
Так или иначе, расстоянием обусловливается градиент видимости — плавный по своей природе. Для любой степени сходства между моделью и ее имитатором — как ошеломляющего, так и практически отсутствующего — должно существовать и такое расстояние, на котором глаза хищника будут обмануты, и такое, чуть короче, на котором они ошибутся с несколько меньшей вероятностью. Следовательно, в ходе эволюции естественный отбор может благоприятствовать постепенно совершенствующемуся сходству, так чтобы необходимое для обмана критическое расстояние плавно сокращалось. Под «глазами хищника» я подразумеваю глаза кого угодно, кого нужно одурачить. В конкретных случаях это могут быть глаза добычи, приемного родителя, самки и так далее.
Я демонстрировал это явление на своих лекциях перед детской аудиторией. Мой коллега, доктор Джордж Макгейвин из Музея естественной истории Оксфордского университета, любезно изготовил по моей просьбе модель «лесной подстилки», покрытую ветками, сухими листьями и мхом. На ней он искусно разместил несколько десятков засушенных насекомых. Некоторые из них, например бронзовки, были довольно заметны; другие, подобно палочникам и подделывающимся под листья бабочкам, оказались великолепно замаскированы; а третьи, вроде коричневых тараканов, занимали в этом смысле промежуточное положение. Детям предлагалось вставать с места, медленно приближаться к макету, выискивать взглядом насекомых и сообщать о каждом замеченном. Находясь достаточно далеко, дети не могли разглядеть даже бросающихся в глаза насекомых. По мере приближения они видели сначала самых заметных, затем таких, как таракан, средней степени различимости, и, наконец, хорошо закамуфлированных. Насекомым, замаскированным наилучшим образом, удалось избегнуть обнаружения даже при близком разглядывании, и когда я указывал на них детям, те восхищенно ахали.
Расстояние — не единственный градиент, к которому применима подобная аргументация. Другой пример — освещенность. В глухую ночь не видно практически ничего и даже самая грубая подделка сойдет за чистую монету. А в самый полдень только очень тщательный и точный имитатор сможет уйти от разоблачения. Между тем и другим временем суток — на рассвете и на закате, в сумерки и просто в серый пасмурный день, в туман и в ливень — складывается плавный и непрерывный спектр качества видимости. И снова естественный отбор благоприятствует постепенному увеличению точности подражания, потому что для любой конкретной степени сходства существует такой уровень видимости, при котором именно такое сходство окажется достаточным и решающим. В ходе эволюции непрерывно возрастающая точность имитирования будет давать преимущество для выживания, поскольку критический порог освещенности, необходимой для обмана, будет таким образом неуклонно повышаться.
Похожий градиент обеспечивается и углом зрения. Иногда хищник видит насекомое, обладающее мимикрией — как качественной, так и не очень, — только краем глаза. А иногда жертва оказывается под прицелом беспощадного взгляда в упор. Непременно должен существовать настолько периферический угол зрения, что даже самый бездарный имитатор не будет узнан. Должен существовать и настолько центральный угол зрения, что даже самый блистательный подражатель не будет в безопасности. Между двумя этими крайностями лежит плавный градиент точек обзора — непрерывный спектр углов зрения. Для любой степени сходства будет существовать такой критический угол, при котором самое незначительное улучшение или ухудшение качества имитации окажется решающим. В ходе эволюции отбор будет благоприятствовать постепенному усилению сходства, так как критический угол зрения, необходимый для успешного обмана, будет плавно смещаться в сторону центра.
В качестве еще одного градиента можно рассматривать совершенство строения вражеских глаз и мозгов, на что я уже намекал выше. Какова бы ни была степень сходства между оригиналом и подделкой, с высокой вероятностью отыщется и такой глаз, который будет обманут, и такой, который не будет. В ходе эволюции опять-таки отбор способствует плавному усилению сходства — так, чтобы оставлять в дураках хищников со все более и более сложно устроенными глазами. Я сейчас говорю не о том, что зрение хищников может эволюционировать в сторону улучшения параллельно с повышением качества мимикрии, хотя это вполне возможно. Я хочу сказать просто, что бывают хищники как с хорошим зрением, так и с плохим. Все они представляют опасность. Плохой подражатель обманет только хищников с плохими глазами. Хороший — практически всех хищников. Между двумя этими крайностями — непрерывный спектр промежуточных вариантов.
Раз уж речь зашла о плохом и хорошем зрении, не могу не вспомнить «неразрешимую задачку», которую так любят задавать креационисты. Какая польза от половинки глаза? Каким образом естественный отбор мог благоприятствовать не вполне идеальному органу зрения? Мне уже приходилось сталкиваться с этим вопросом раньше, и тогда в качестве ответа я представил подборку рисунков, изображавших глаза, находящиеся на всех возможных промежуточных ступенях совершенства, какие встречаются в реальности у представителей различных типов животного царства. Сейчас же я собираюсь поместить зрение в выстраиваемый мною ряд теоретических градиентов. Существует некий градиент — непрерывный спектр — задач, для выполнения которых может быть использован глаз. В настоящий момент я использую свои глаза для распознавания появляющихся на компьютерном экране букв. Эта задача требует хорошего, острого зрения. Я уже достиг такого возраста, что больше не могу читать без помощи очков — пока еще не очень сильных. По мере моего старения мне будут выписывать очки со все более и более мощными линзами. Постепенно и неуклонно мне будет становиться все труднее и труднее видеть мелкие подробности без очков. Вот вам и еще один континуум — возрастной.
У любого нормального человека, как бы стар он ни был, зрение лучше, чем у насекомого. Существуют задачи, которые могут успешно выполняться людьми с относительно слабым зрением, даже почти полностью слепыми. Вы сможете играть в теннис, даже если видимый вами мир довольно расплывчат, потому что теннисный мяч — это крупный предмет, местоположение и передвижение которого заметны и тогда, когда глаза не наведены на резкость. Глаза стрекозы, неважные по нашим меркам, хороши для насекомого, и стрекоза способна ловить других насекомых на лету — задача не менее сложная, чем попадать ракеткой по мячу. А для выполнения таких задач, как «не врезаться в стену», «пройти по краю обрыва» или «перейти реку вброд», сгодится зрение и намного слабее. Еще более никудышные глаза сумеют различить нависшую тень, которая может быть отброшена проплывающим облаком, но может и предвещать приближение хищника. И даже еще менее совершенные глаза помогут вам отличить день от ночи, что полезно в числе прочего для синхронизации периодов размножения и для того, чтобы знать, когда пора отправляться ко сну. Существует непрерывный спектр задач, для выполнения которых можно применить глаз, так что для любого конкретного качества зрения, от изумительного до отвратительного, существует такой уровень сложности задачи, когда ничтожнейшее улучшение зоркости может оказаться решающим. Следовательно, нет ничего трудного в том, чтобы представить себе постепенную эволюцию глаза: начиная от примитивных и грубых начальных этапов, минуя плавный континуум промежуточных звеньев и заканчивая тем совершенством, какое мы наблюдаем у ястреба или у человека в его молодые годы.
Таким образом, креационистская загадка — «Какой прок от половинки глаза?» — это вопрос плевый, выеденного яйца не стоящий. Половинка глаза ровно на 1 % лучше 49 % глаза, которые при этом лучше 48 %, и разница тут существенна. Более затруднительным может показаться следующее неизбежное продолжение дискуссии: «Рассуждая как физик, я не могу поверить, что на возникновение из ничего такого сложного органа, как глаз, хватило бы времени. Вы действительно полагаете, будто его было достаточно?» Оба вопроса — как этот, так и предыдущий — проистекают из «убеждения личным недоверием». Публике тем не менее хотелось бы услышать ответ, и обычно в таких случаях я просто призывал на помощь громадную протяженность геологического времени. Если представить себе один век как шаг, то все, что мы называем нашей эрой, окажется короче, чем поле для крикета. А чтобы добраться в этом масштабе до возникновения первых многоклеточных организмов, вам пришлось бы полностью протопать расстояние от Нью-Йорка до Сан-Франциско.
Теперь же выясняется, что апеллировать здесь к сокрушительно огромной геологической шкале времен — значит палить из пушек по букашкам. Аналогия с изнурительной прогулкой от одного побережья к другому наглядно иллюстрирует количество времени, имевшегося для эволюции глаза. Однако двое шведских ученых, Дан Нильссон и Сусанна Пельгер, в своей недавней работе показали, что с лихвой хватило бы и малой доли от этого. Кстати, когда говорят «глаз», обычно подразумевают глаз позвоночных, хотя пригодные к использованию и формирующие изображение органы зрения возникали с нуля во многих различных группах беспозвоночных от сорока до шестидесяти раз независимо. В ходе этих более чем сорока не связанных друг с другом эволюционных процессов было изобретено по меньшей мере девять четко различимых конструкций, в том числе глаз, устроенный по принципу камеры-обскуры, две разновидности глаза-«фотоаппарата», глаз, функционирующий как криволинейный отражатель («спутниковая тарелка»), и несколько моделей фасеточных глаз. Нильссон и Пельгер сосредоточили свое внимание на глазах, устроенных по типу камеры с линзой-хрусталиком, то есть таких, что хорошо развиты у позвоночных и осьминогов.
Как подступиться к тому, чтобы оценить продолжительность времени, необходимого для некоего количества эволюционных преобразований? Для выяснения величины каждого эволюционного шага нам потребуется единица измерения, и представляется разумным определить ее как некую долю от того, что имелось прежде. В качестве такой единицы измерения для эволюции анатомических показателей Нильссон и Пельгер использовали постепенное изменение признака на 1 %. Это просто условная единица — вроде калории, по определению представляющей собой количество энергии, необходимое для выполнения некой работы. Данную однопроцентную единицу проще всего применять, когда преобразования происходят только в одном измерении. Если, к примеру, сделать неправдоподобное предположение, будто естественный отбор благоприятствует бесконечному удлинению хвоста у райской птицы, то сколько потребуется шагов, чтобы длина хвоста проэволюционировала от одного метра до одного километра? Увеличение длины хвоста на 1 % — это то, чего обычный орнитолог и не заметит. И тем не менее, чтобы удлинить хвост до километра, таких шагов потребуется на удивление мало: менее семисот.
Удлинение хвоста от одного метра до одного километра — это, конечно, прекрасно (и, конечно, нелепо), но как разместить на этой шкале эволюцию глаза? Проблема в том, что в случае с глазом множество параметров должно меняться одновременно в самых разных направлениях. Целью Нильссона и Пельгер было разработать такие компьютерные модели эволюционирующего глаза, которые отвечали бы на два вопроса. Первый вопрос примерно тот же, каким мы с вами здесь уже неоднократно задавались, только они рассматривали его более методично, с использованием компьютера. Вопрос этот таков: «Существует ли непрерывный градиент преобразований от гладкой кожи до полноценного глаза позвоночного животного, так чтобы каждый следующий промежуточный этап представлял собой усовершенствование?» (В отличие от инженеров-людей, естественный отбор не способен временно понижать планку качества — даже если это сулит возможность поднять ее значительно выше впоследствии.) Второй вопрос — тот, с которого мы начали данный раздел: «Сколько времени займут необходимые эволюционные изменения?»
В своих компьютерных симуляциях Нильссон и Пельгер не делали попыток моделировать процессы, происходящие внутри клеток. В их работе одиночная светочувствительная клетка — не будет большим грехом назвать ее фотоэлементом — изначально считалась уже возникшей. Неплохо было бы как-нибудь в будущем создать еще одну компьютерную модель — уже на внутриклеточном уровне, — чтобы показать, как самый первый живой фотоэлемент получился в ходе пошаговых преобразований некой клетки-предшественницы, имевшей более общее назначение. Но с чего-то нужно начинать, и Нильссон с Пельгер начали с появления светочувствительной клетки. Они работали на уровне тканей — то есть не отдельных клеток, а состоящего из них материала. Кожа — это ткань, так же как и выстилка кишечника, мышца или печень. Ткани могут самым различным образом видоизменяться под влиянием случайных мутаций. Клеточные пласты становятся обширнее или компактнее, толще или тоньше. В специфическом случае прозрачных тканей — наподобие хрусталика — мутации могут менять показатель преломления (степень отклонения падающего света от первоначального направления) на том или ином участке.
Моделировать глаз, в отличие, скажем, от лапы бегущего гепарда, приятно тем, что его эффективность легко оценить при помощи элементарных законов оптики. Глаз можно представить в виде двумерного поперечного среза, после чего компьютер без затруднений рассчитает нам остроту зрения (пространственное разрешение) в виде определенного вещественного числа. Прийти к подобному численному выражению для эффективности конечности или позвоночника гепарда было бы намного сложнее. Отправной точкой в модели Нильссона и Пельгер была плоская сетчатка, лежащая поверх плоского пигментного слоя и покрытая плоским защитным слоем прозрачной ткани. Этот прозрачный слой мог локально подвергаться действию мутаций, менявших его показатель преломления. Также модели позволялось претерпевать деформации в любых случайных направлениях, но при соблюдении того условия, чтобы каждое изменение было небольшим и непременно в лучшую сторону по сравнению с тем, что имелось до этого.
Результаты были получены незамедлительно — и оказались однозначными. По мере того как форма моделируемого глаза преображалась на компьютерном экране, плавное увеличение остроты зрения уверенно вело от исходной плоской поверхности через промежуточный этап мелкой выемки ко все более углубляющейся чаше. Прозрачный слой утолщался, заполняя чашу и образуя легкое выпячивание с наружной стороны. А дальше происходил почти что фокус: участок этого прозрачного содержимого уплотнялся в обособленную сферическую область с более высоким показателем преломления. И не просто более высоким, а плавно меняющимся от периферии к центру таким образом, чтобы данная сферическая область функционировала как превосходная градиентная линза. Градиентные линзы плохо известны мастерам рукотворной оптики, но внутри живых глаз они — дело обычное. Люди производят линзы, шлифуя стекло, чтобы придать ему определенную форму. Мы изготавливаем составные линзы (вроде дорогих линз с фиолетовым отливом, используемых в современных фотоаппаратах), соединяя несколько линз вместе, но каждая из этих простых линз сделана из стекла, однородного по всей своей толщине. У градиентных же линз, напротив, показатель преломления составляющего их вещества варьирует от участка к участку, возрастая, как правило, по направлению к центру. Хрусталик глаза рыб — градиентная линза. Физикам давно известно, что меньше всего искажений градиентная линза дает при достижении определенной теоретически оптимальной пропорции между фокусным расстоянием и радиусом, называемой соотношением Матиссена. Компьютерная модель Нильссона и Пельгер безошибочно пришла именно к этому соотношению.
Что же касается вопроса, как много времени могли занять все эти эволюционные перемены, то, чтобы ответить на него, Нильссону и Пельгер пришлось сделать некоторые допущения насчет генетики популяций в естественных условиях. Им требовалось ввести в свою модель правдоподобные значения различных количественных величин, таких как «наследуемость». Наследуемость — это мера того, в какой степени изменчивость обусловлена генами. Удобный способ определения наследуемости — сравнивать сходство признака у однояйцевых (то есть «идентичных») близнецов по сравнению с обычными двойняшками. Так, в одном исследовании было показано, что наследуемость длины ног у мужчин равняется 77 %. Если бы она равнялась 100 %, это означало бы, что вы можете измерить ногу одного из идентичных близнецов и получить точное значение длины ноги другого, даже если близнецы не росли вместе. А наследуемость, равная 0 %, означала бы, что нижние конечности однояйцевых близнецов не более похожи друг на друга, чем ноги двух любых случайных представителей данной популяции, обитающих в тех же условиях. Вот характерные для человека значения наследуемости некоторых других признаков: поперечный диаметр головы — 95 %, высота в положении сидя — 85 %, длина рук — 80 %, рост — 79 %.
Наследуемость часто превышает 50 %, и потому Нильссон и Пельгер со спокойной совестью ввели в свою модель глаза 50-процентное значение наследуемости. Это было консервативное, или «пессимистическое», допущение, которое завышало окончательную оценку времени, необходимого для эволюции глаза, по сравнению с более реалистичным значением, скажем, в 70 %. Шведские исследователи предпочли ошибиться в сторону завышения, поскольку выводы, будто для эволюции такого сложно устроенного объекта, как глаз, требуется не так уж много времени, вызывают у нас интуитивный скепсис.
По этой же причине Нильссон и Пельгер сделали выбор в пользу пессимистических значений коэффициента вариации (показателя того, насколько велика изменчивость в популяции) и интенсивности отбора (параметра, отражающего пользу зрительных усовершенствований для выживания). В своих допущениях они пошли даже на то, чтобы считать, что в каждом новом поколении видоизменяться может только какой-то один составной элемент глаза. Одновременные преобразования сразу нескольких участков, значительно ускорившие бы эволюцию, были объявлены вне закона. Но даже несмотря на все эти осторожные оценки, время, затраченное на возникновение глаза рыбы из гладкого участка кожи, было ничтожным: менее четырехсот тысяч поколений. Для тех мелких животных, о которых мы ведем тут речь, можно отвести по году на поколение, так что, по-видимому, на эволюцию полноценного глаза, устроенного по типу камеры с линзой, нужно меньше полумиллиона лет.
В свете результатов Нильссона и Пельгер не приходится удивляться тому, что пресловутый «необъяснимый» орган зрения независимо возникал в животном царстве не менее сорока раз. Он мог бы полторы тысячи раз подряд возникать из ничего на протяжении любой отдельно взятой линии, ведущей от предков к потомкам, — времени было достаточно. Если исходить из типичного для мелких животных интервала между поколениями, то время, необходимое для эволюции глаза, не только не выходит за рамки правдоподобия своей громадной продолжительностью, но и оказывается слишком кратким для того, чтобы геологи могли его измерить! Это геологическое мгновение.
Делай добро украдкой. Главная особенность эволюции — ее постепенность. Это вопрос не столько факта, сколько принципа. На некоторых своих этапах эволюция вполне может принимать (а может и не принимать) неожиданный оборот. Возможны стремительные эволюционные рывки и даже скачкообразные макромутации — резкие изменения, делающие детеныша не похожим ни на одного из родителей. Несомненно, случаются и внезапные вымирания — по-видимому, вследствие глобальных природных катаклизмов вроде столкновения Земли с кометой, — освобождающие место для дублеров, которые начинают оперативно совершенствоваться, как это было с млекопитающими, сменившими динозавров. Весьма вероятно, что на самом деле эволюция не всегда постепенна. Но она должна быть постепенной в тех случаях, когда с ее помощью пытаются объяснить возникновение сложных, будто заранее спроектированных, объектов, таких как глаз. Здесь, не будучи плавной, она вообще перестает объяснять что-либо и возвращает нас к такому понятию, как чудо, служащему просто-напросто синонимом отсутствия какого бы то ни было объяснения.
Причина, почему нас так восхищают глаз и опыляемая осами орхидея, состоит в том, что они невероятны. Шансы их случайной самопроизвольной сборки слишком ничтожны, чтобы иметь отношение к реальному миру. Постепенная эволюция, движущаяся мелкими шажками, каждый из которых — счастливая, но не слишком большая случайность, — вот ключ к отгадке. Если же эволюция не постепенна, то никакой отгадки нет, а есть все та же загадка, только иначе сформулированная.
Бывает трудно себе представить, каковы могли быть промежуточные этапы такой плавной эволюции. Это испытание для нашей сообразительности, но, если она не в состоянии его выдержать, тем хуже для нее. Отсюда не следует, что промежуточных этапов не было. Одним из самых суровых испытаний, каким подвергается наша изобретательность в придумывании промежуточных форм, мы обязаны прославленному «языку танца» у пчел, открытому в классической работе Карла фон Фриша, которой этот ученый более всего знаменит. Конечный продукт эволюции кажется здесь настолько замысловатым и хитроумным, настолько не похожим на все то, чего мы в принципе могли бы ожидать от насекомых, что вообразить предшествовавшие ему промежуточные стадии непросто.
Медоносные пчелы сообщают друг другу о местонахождении цветущих растений при помощи строго закодированного танца. Если пища находится совсем близко от улья, они исполняют «круговой танец». Это просто побуждает других пчел устремиться наружу и прочесывать окрестности улья. Ничего особенно примечательного. Но крайне примечательно то, что происходит, когда пища находится несколько дальше. Обнаружившая ее пчела-разведчица исполняет так называемый виляющий танец, фигура и продолжительность которого информируют других пчел как о расстоянии от улья до добычи, так и о том, в каком направлении та находится. «Виляющий танец» исполняется внутри улья на вертикальной поверхности сот. В улье темно, так что другие пчелы не видят танца. Они чувствуют его, а также слышат, поскольку танцующая пчела сопровождает свое выступление ритмичным свистящим шорохом. Фигура танца представляет собой сплющенную восьмерку с поперечным отрезком посередине. Именно направление этого прямолинейного пробега и является искусно зашифрованным сообщением о том, в какую сторону следует лететь за взятком.
Прямолинейный пробег в танце не указывает непосредственно дорогу к пище. Это невозможно, ведь танец исполняется на вертикальной поверхности сот, а их положение неизменно и не зависит от местонахождения добычи. Пищу следует искать где-то на горизонтальной местности. А вертикальные соты больше смахивают на карту, прикрепленную к стене. Линия, проведенная по карте, не показывает нужного направления напрямую, но оно считывается благодаря произвольно выбранным условным обозначениям.
Для понимания условных обозначений, принятых у пчел, необходимо в первую очередь знать, что пчелы, как и многие другие насекомые, используют солнце в качестве компаса. Мы тоже так делаем, хотя и с меньшей точностью. У этого способа есть два минуса. Во-первых, солнце часто бывает скрыто за облаками. Пчелы решают данную проблему при помощи чувства, у нас с вами отсутствующего. И снова человеком, открывшим, что они видят направление поляризации света и это позволяет им понять, где находится солнце, даже когда самого солнца не видно, был фон Фриш. Второй недостаток солнечного компаса заключается в том, что в течение дня солнце перемещается по небосклону. Пчелы справляются с этим благодаря имеющемуся у них внутреннему хронометру. Невероятно, но фон Фриш обнаружил, что у пчел, запертых в улье в течение многих часов после возвращения из разведывательной экспедиции, отрезок прямолинейного пробега в танце медленно поворачивается, как если бы он был часовой стрелкой на 24-часовом циферблате. Находясь внутри улья, они не могут видеть солнце, но, чтобы не отставать от него, плавно меняют направление танца в соответствии со своими внутренними часами. Восхитительно, что породы пчел из Южного полушария проделывают то же самое в обратную сторону — именно так, как и следовало ожидать.
Теперь перейдем к собственно коду. Если, танцуя, пчела пробегает по прямой линии точно вверх по стенке сот, значит, пища находится в том же направлении, что и солнце. Точно вниз — это говорит о строго противоположном направлении. Промежуточные углы обозначают именно то, что вы и предполагаете. Пятьдесят градусов влево от вертикали соответствуют 50° влево от направления к солнцу на горизонтальной плоскости. Впрочем, информация передается через танец отнюдь не с точностью до градуса. Да и с чего бы — ведь это мы, люди, условились делить окружающее пространство на 360°. А пчелы делят его на свои восемь пчелиных градусов. Собственно говоря, приблизительно так же поступают и люди, если они не профессиональные штурманы. Свой повседневный компас мы привыкли делить на восемь секторов: С, СВ, В, ЮВ, Ю, ЮЗ, З, СЗ.
Расстояние до пищи также закодировано в танце. Точнее, различные характеристики танца — скорость кружения, частота виляний брюшком, частота, с какой пчелы повторяют звук, похожий на шорох, — коррелируют с удаленностью добычи, и, следовательно, любая из этих характеристик, так же как и любое их сочетание, может служить другим пчелам для оценки расстояния. Чем ближе пища, тем быстрее танец. Нетрудно запомнить, если принять в соображение, что пчела, обнаружившая еду поблизости от улья, должна быть более воодушевленной и менее усталой по сравнению с пчелой, принесшей свои вести издалека. И это не просто удобная памятка — как мы увидим, здесь кроется ключ к пониманию эволюции пчелиного танца.
Итак, пчела-разведчица находит хороший пищевой ресурс. Нагруженная нектаром и пыльцой, она возвращается в улей и передает свою ношу пчелам-приемщицам, после чего приступает к танцу. На любой вертикальной стенке сот — где именно, значения не имеет — она упорно и беспрерывно выписывает приплюснутую восьмерку. Другие рабочие особи скапливаются вокруг нее, сосредоточившись на своих ощущениях и прислушиваясь. Они оценивают частоту шороха, а также, возможно, скорость кружения. Пока танцовщица виляет своим брюшком, они определяют, под каким углом относительно вертикали расположен прямой участок траектории танца. Затем направляются к выходу и бросаются из царящей в улье тьмы на дневной свет. Заметив, как расположено солнце — не высоту его, а компасное направление к нему на горизонтальной плоскости, — они летят по прямой линии, отклонение которой от этого направления соответствует тому, под каким углом относительно вертикали танцевала на сотах пчела-разведчица. Выбранного курса они придерживаются не сколько им заблагорассудится, а преодолевая расстояние, (обратно) пропорциональное частоте шороха, издававшегося танцовщицей (точнее, логарифму этой частоты). Занятно, что, если разведчица набрела на источник пищи окольными путями, танцем она не показывает проделанный ею маршрут, а реконструирует компасное направление.
В историю о танцующих пчелах трудно поверить, и некоторые действительно отказывались верить в нее. К этим скептикам и к недавним экспериментам, доказавшим все окончательно и бесповоротно, мы вернемся в следующей главе. А здесь мне хотелось бы обсудить плавную эволюцию пчелиного танца. Как могли выглядеть промежуточные этапы его становления и как он функционировал, не будучи завершенным?
Кстати говоря, формулировать вопрос таким образом не вполне правильно. Ни одно существо не проживает свою жизнь в качестве некой «незавершенной», «промежуточной» стадии. Древние, давным-давно исчезнувшие пчелы, чьи танцы можно сегодня, задним числом, рассматривать как стадии формирования танца современной медоносной пчелы, жили совсем неплохо. Они проживали свои полноценные пчелиные жизни и понятия не имели о том, что находятся «на пути» к чему-то «лучшему». Более того, нынешний, «современный» пчелиный танец может быть отнюдь не последним словом — ничто не мешает ему со временем проэволюционировать во что-то еще более впечатляющее, когда ни нас, ни наших пчел уже не будет на свете. Все это, однако, никоим образом не отменяет стоящей перед нами загадки, каким образом этот танец в его нынешнем виде мог возникнуть при помощи постепенных преобразований. На что могли быть похожи промежуточные этапы его эволюции и как они работали?
Уделил внимание данному вопросу и сам фон Фриш. Он подошел к нему через генеалогию — изучая дальних родственников медоносной пчелы. Они не могут быть ее предками, поскольку живут в одно время с ней. Но они могли сохранить предковые признаки. Медоносная пчела — это насекомое умеренного пояса, вынужденное гнездиться в укрытиях вроде дупел и пещер. Ее ближайшая родня — пчелы тропических широт, имеющие возможность устраивать свои гнезда под открытым небом и свешивающие свои соты с древесных сучьев или обнаженных скал. Следовательно, они могут видеть солнце во время танца и им нет необходимости прибегать к условным символам, где вертикаль «обозначает» местонахождение солнца. Солнце вполне может обозначать само себя.
Одна из таких тропических родственниц — карликовая пчела Apis florea — танцует на горизонтальной верхней поверхности сот. Прямолинейный пробег в ее танце совершается непосредственно в направлении к источнику пищи. В картографических обозначениях нет нужды — ведь можно указать путь напрямую. Это, несомненно, выглядит правдоподобной промежуточной стадией эволюции наших медоносных пчел, но нам все равно необходимо задуматься и о других стадиях — предшествовавших этой и следовавших за ней. Что могло служить предтечей танцу карликовой пчелы? Зачем пчеле, недавно обнаружившей пищу, остервенело выписывать напоминающую восьмерку фигуру, прямой участок которой указывает в сторону добычи? Предполагают, что это ритуализированная форма разбега. По гипотезе фон Фриша, до возникновения танца пчела-разведчица, выгрузив взяток, просто улетала обратно в сторону источника пищи. Готовясь подняться в воздух, она должна была повернуться в нужном направлении и сделать несколько шагов. Естественный отбор мог благоприятствовать любой тенденции увеличивать как время, так и длину разбега, если это подстрекало других пчел следовать за разведчицей. Быть может, танец представляет собой своего рода ритуально повторяемый разбег. Такое предположение убедительно, поскольку пчелы — независимо от того, используют они танцы или нет, — часто прибегают к менее замысловатой тактике простого следования друг за дружкой в полете за добычей. Еще один факт, добавляющий правдоподобия этой идее, состоит в том, что пчелы во время танца слегка расправляют крылья, как будто готовясь взлететь, а мускулы их крыльев вибрируют — недостаточно интенсивно для того, чтобы подняться в воздух, но достаточно для произведения шума, являющегося важной частью танцевального кода.
Очевидный способ увеличить время и длину разбега — повторить его, то есть вернуться на исходную позицию и еще раз проделать несколько шагов в направлении еды. Чтобы возвратиться к старту, есть две возможности: свернуть с дороги вправо или влево. Если вы постоянно сворачиваете только в одну сторону, то неясно, какое направление вашего движения соответствует подлинной дорожке разбега, а какое — возвращению на начало этой дорожки. Лучший способ избежать такой двусмысленности — поворачивать попеременно то влево, то вправо. Вот почему естественный отбор предпочел рисунок в виде восьмерки.
А как эволюционировала взаимосвязь между расстоянием до пищи и скоростью танца? Если бы между ними имелась прямая зависимость, объяснить ее было бы непросто. Но, как вы помните, в действительности все обстоит как раз наоборот: чем расстояние меньше, тем танец быстрее. И это сразу же позволяет представить правдоподобную версию плавной эволюции. Прежде чем танец полностью сформировался, разведчицы, вероятно, исполняли свое ритуализированное повторение разбега перед взлетом, не придерживаясь какой-то определенной скорости. Та могла быть какой угодно в зависимости от их самочувствия. А теперь скажите: если вы пролетели несколько миль, будучи под завязку нагружены нектаром и пыльцой, бойко ли вам будет плясаться на сотах? Вряд ли — скорее всего, вы будете обессилены. А вот если вы только что обнаружили богатый источник пищи вблизи от улья, то после недолгой обратной дороги будете чувствовать себя еще свежо и бодро. Несложно представить себе, как исходное случайное влияние дальности расстояния на замедленность танца было закреплено и преобразовано в строгий, надежный код.
Ну и самый затруднительный переход из всех. Каким образом древний танец, прямой участок фигуры которого был направлен непосредственно в сторону еды, смог превратиться в танец нынешний, где угол к вертикали указывает на расположение источника пищи относительно солнца? Такая трансформация была необходима отчасти потому, что внутри улья темно и солнца не видно, а отчасти — потому что, танцуя на вертикальной поверхности, невозможно указать прямое направление на источник пищи, если только эта поверхность уже случайно не повернута в его сторону. Но продемонстрировать необходимость такого преобразования недостаточно. Мы обязаны также объяснить, как это сложное превращение могло правдоподобно осуществиться через плавную вереницу промежуточных форм.
Задача кажется неразрешимой, но нам на выручку приходит одна замечательная особенность нервной системы насекомых. С самыми различными их представителями, от жуков до муравьев, был проделан следующий выдающийся опыт. Допустим, у нас есть жук, ползущий по горизонтальной доске при электрическом освещении. В первую очередь докажем, что насекомое ориентируется с помощью «светового компаса». Передвиньте лампу — и оно соответствующим образом сменит курс. Скажем, если насекомое двигалось под углом 30° к свету, оно так изменит свою траекторию, чтобы продолжать движение под тем же углом относительно нового местоположения лампы. В сущности, вы можете управлять жуком, как вам вздумается, используя световой луч вместо руля. Это свойство насекомых известно давно: они используют солнце (или луну, или звезды) в качестве компаса и их нетрудно перехитрить при помощи электрической лампочки. Пока что все просто. А теперь про выдающийся опыт. Выключите свет и одновременно поверните доску вертикально. Нимало не смутившись, жук продолжает свой путь. И — о чудо! — меняет направление таким образом, чтобы двигаться относительно вертикали под тем же углом, под каким он прежде двигался относительно света (в нашем примере это 30°). Никто не знает, в чем тут причина, но факт остается фактом. По-видимому, так проявляется какой-то случайный дефект в работе нервной системы насекомых — обман чувств, замыкание проводов между ощущением гравитации и зрением (быть может, нечто сродни тому, как мы, ударяясь головой, видим вспышку). В любом случае это, вероятно, и обеспечило необходимый мостик для эволюции условного соглашения «вертикаль символизирует солнце» в танце медоносных пчел.
Показательно, что, если внутри улья включить освещение, пчелы забывают о гравитации и пользуются непосредственно направлением света для обозначения солнца в своем коде. Этот давно известный факт лег в основу одного из самых хитроумных экспериментов, когда-либо проводившихся, — эксперимента, окончательно подтвердившего истинное назначение пчелиного танца. Я еще вернусь к этому в следующей главе. Между тем мы с вами нашли правдоподобную серию плавных переходов, благодаря которым танец современных медоносных пчел смог бы развиться из простейших зачатков. Не исключено, что эта история в том виде, как я вам ее рассказал, опираясь на идеи фон Фриша, не соответствует действительности. Но что-то подобное, несомненно, имело место. Мой рассказ был ответом на естественный человеческий скептицизм — на «убеждение личным недоверием», возникающее всякий раз при встрече с по-настоящему замысловатыми и труднопостижимыми явлениями природы. В таких случаях скептик говорит: «Я не в состоянии вообразить правдоподобный ряд промежуточных форм, а значит, его не было и наблюдаемый феномен возник самопроизвольно, чудом». Фон Фриш представил нам убедительный пример такого ряда. Даже если это и не вполне тот же ряд, что имел место в действительности, самого факта его правдоподобия достаточно, чтобы выбить у «убеждения личным недоверием» почву из-под ног. То же справедливо и для всех прочих рассмотренных нами примеров: от подражающих осам орхидей до глаза-«фотокамеры».
Те, кто скептически относится к дарвинистскому градуализму, могут прикрываться любым из множества необычных и занимательных явлений природы. Например, меня как-то попросили обосновать постепенную эволюцию живых существ, обитающих в глубоких впадинах Тихого океана, где нет света, а давление воды порой превышает 1000 атмосфер. Вокруг горячих вулканических жерл в глубоких тихоокеанских впадинах возникла целая экосистема. В бактериях там работает совершенно иная, альтернативная биохимия, использующая вулканическое тепло, а вместо кислорода потребляющая серу. Сообщество более крупных организмов зависит в конечном итоге от этих серобактерий, подобно тому как обычная жизнь существует за счет зеленых растений, улавливающих энергию солнца.
Все животные, что входят в состав серных экосистем, приходятся родственниками менее странным животным, обитающим в других местах. Как они эволюционировали, через какие промежуточные этапы? Ну что же, аргументация все та же. Для объяснения нам нужен хотя бы один естественный градиент, а по мере погружения в морскую пучину градиентов будет хоть отбавляй. Давление в 1000 атмосфер чудовищно, но оно только количественно превышает 999 атмосфер, которые лишь количественно больше 998, и так далее. Морское дно обеспечивает градиент глубины от 0 до 33 000 футов, со всеми промежуточными значениями. Давление плавно меняется от 1 атмосферы до 1000. Освещенность непрерывно варьирует от яркого дневного света у поверхности до полнейшего мрака, оживляемого лишь редкими скоплениями люминесцентных бактерий да светящимися органами глубоководных рыб. Нет никаких резких перепадов. Для любого уровня давления и темноты, к которому животное уже приспособлено, всегда возможно такое устройство организма, только незначительно отличающееся от уже существующих, которое позволит выжить в условиях на одну морскую сажень глубже, на один люкс темнее. Для любого… Впрочем, эта глава и так уже получилась слишком длинной. Вам известны мои методы, Ватсон. Применяйте их.

