Часть III
Главная теория биологии
В главе 5 мы уже упоминали максиму выдающегося эволюционного генетика Феодосия Добржанского – “Ничто в биологии не имеет смысла, кроме как в свете эволюции”. Эти слова были написаны в 1975 году, и за прошедшие с тех пор почти полвека не появилось ни фактов, ни теорий, которые поколебали бы их справедливость. А поскольку, как мы видели в предыдущей части книги, в понимании эволюции и ее законов у дарвинизма сегодня просто не осталось соперников, это автоматически делает его центральной теорией всей совокупности наук о жизни.
Сказанное, разумеется, не означает, что чем бы ни занимался ученый той или иной биологической специальности, он непременно занимается вопросами эволюции. Различные дисциплины предоставляют множество задач и тем, которыми можно успешно заниматься, не прибегая к эволюционным представлениям. Если, допустим, исследователь изучает изменения в мозге, возникающие при формировании алкогольной или наркотической зависимости, взаимодействие разных типов клеток в ходе иммунной реакции, запаховые сигналы насекомых или зависимость продуктивности (скорости прироста биомассы) луга от присутствия и обилия на нем определенного вида муравьев, он вполне может не думать об эволюции. (Один биохимик с некоторой даже гордостью сообщил автору этих строк, что за десятилетия научной работы ему ни разу не понадобилось привлекать эволюционные соображения.) Тем более это справедливо для прикладных исследований. Но как только исследователь переходит от вопросов типа “как это устроено?” и “как это работает?” к вопросу “почему это устроено и работает именно так?” – ему без эволюции уже никак не обойтись.
Такое исключительное положение эволюционной теории, естественно, требует, чтобы она плодотворно взаимодействовала со всеми областями биологии. Слово “плодотворно” здесь ключевое: важно не просто уметь истолковать любые добытые факты с эволюционно-селекционистской точки зрения, но предложить такой взгляд на проблемы той или иной дисциплины и на изучаемые ею явления, который даст новое знание – или (что было бы даже лучше) укажет пути, на которых можно получить такое знание.
Но на то, чтобы рассмотреть “в свете эволюции” все области современной биологии, не хватит и нескольких таких книг. К тому же выше мы уже довольно подробно раскрыли отношения дарвинизма с генетикой и слегка коснулись его взаимодействия с иммунологией и науками о поведении. Поэтому в этой части книги мы ограничимся всего несколькими областями фундаментальной биологии, взаимоотношения которых с дарвиновской теорией, как мне представляется, больше всего заслуживают научно-популярного рассказа.
И начать нам придется с области исследований, настолько связанной с теорией эволюции, что многие даже интересующиеся наукой люди их не различают. Я говорю о проблеме происхождения жизни.
Глава 12
В начале было… что?
От оппонентов эволюционной теории – как заведомо предубежденных, так и добросовестных – часто приходится слышать такое возражение: ну хорошо, мол, допустим, эта теория более-менее убедительно описывает эволюцию уже существующей жизни. Но откуда эта самая жизнь взялась? Как могла возникнуть система, способная к такой эволюции посредством естественного отбора? Ведь отбор может действовать лишь на то, что способно к самовоспроизведению. А даже самые простые из известных нам самовоспроизводящихся систем все-таки слишком сложны, чтобы возникнуть в результате чисто случайных событий. В конце концов, будь иначе, мы бы регулярно видели самозарождение простейших живых существ из каких-то неживых компонентов. Однако, как доказал еще Луи Пастер, такого не происходит. Если жизнь не может возникнуть сама из неживой материи, но при этом существует – не означает ли это, что ее кто-то сотворил?
Строго говоря, данный вопрос не относится к теме этой книги. Ее задача – рассказать о биологической эволюции, а не обо всех современных представлениях о природе и ее развитии. Если уж на то пошло, механика Ньютона тоже не объясняла, откуда взялись планеты и другие небесные тела – достаточно было того, что они несомненно существуют, а она успешно описывает их движение и взаимодействие. Вопрос же о происхождении Солнечной системы и составляющих ее тел получил более-менее удовлетворительное решение много позже и в рамках совсем других теорий и дисциплин. С другой стороны, тем, кого действительно интересует проблема происхождения жизни, можно посоветовать прочесть вышедшую в 2016 году прекрасную книгу Михаила Никитина “Происхождение жизни: от туманности до клетки” – полное, ясное, связное и максимально беспристрастное изложение того, что знает и думает современная наука об этой проблеме.
Но ограничиваться таким ответом я не хочу. И не только потому, что это оставляло бы впечатление, что проблема происхождения жизни действительно является уязвимым пунктом – если не теории эволюции, то современной научной картины мира в целом, – но еще и потому, что успехи исследований последних десятилетий в этой области не только подкрепляют и дополнительно обосновывают эволюционную теорию, но и демонстрируют неожиданную плодотворность в ней дарвинистского подхода – хотя, казалось бы, он может вступать в действие только с момента, когда жизнь уже начала существовать. Именно поэтому глава, посвященная вопросам происхождения жизни, помещена в ту часть книги, где речь идет о роли эволюционной теории в разных областях фундаментальной биологии.
Разумеется, излагать во всех подробностях современное состояние проблемы происхождения жизни я не буду: во-первых, потому, что это уже сделал Никитин (и сделал гораздо глубже и компетентнее, чем мог бы это сделать я), во-вторых, потому, что такое изложение требует соответствующего объема: в книге Никитина более 500 страниц, не считая списка литературы. Поэтому я позволю себе коснуться лишь некоторых аспектов этой проблемы – тех, которые вызывают больше всего вопросов и недоразумений, а также наиболее тесно связаны с собственно эволюционной проблематикой.
Трижды невозможная жизнь
Прежде всего нужно разобраться с тем, почему сейчас мы не видим самозарождения живого из неживого – и почему, тем не менее, мы полагаем, что оно было возможно в далеком прошлом, около четырех миллиардов лет назад. Самозарождение жизни на сегодняшней Земле невозможно, видимо, потому, что… на ней уже есть жизнь. Это не шутка и не софизм: любые сценарии происхождения жизни предполагают, что в самом начале этого процесса на Земле уже имелся широкий спектр простых органических веществ, возникших чисто химическим путем из неорганических предшественников. Эксперименты, воспроизводящие условия на поверхности древней Земли, показывают, что такое вполне возможно и даже неизбежно. Более того: многие органические вещества, включая даже некоторые аминокислоты, обнаруживаются (пусть и в следовых количествах) в составе метеоритов, комет и космической пыли, так что само по себе появление простейшей органики без участия живых организмов не представляет особой теоретической проблемы.
Дело, однако, в том, что в наше время любая органика (в том числе и абиогенного происхождения), оказавшись в водах Мирового океана или внутренних водоемов, на поверхности земли, в почве или в атмосфере, быстро станет добычей вездесущих бактерий и других микроорганизмов. Что, естественно, лишит ее возможности чисто химической эволюции, отдаленным результатом которой могло бы стать появление “жизни-2”. Хотя мы не знаем, сколько времени занял путь от простейших органических соединений до первых самокопирующихся молекул, предполагается, что на это ушел не один миллион лет (а скорее – не одна сотня миллионов лет), в то время как любая попавшая в современную биосферу органика будет сожрана гораздо быстрее. Даже такие химически инертные и “неудобные” для вовлечения в метаболические процессы органические вещества, как углеводороды, уничтожаются бактериями довольно быстро: уже через несколько лет после катастрофических нефтяных разливов, при которых в море выливаются сразу сотни тысяч тонн нефти, свободные углеводороды обнаруживаются лишь в следовых количествах (причем в основном в телах животных-фильтраторов и в других “убежищах”, где они недоступны бактериям). Конечно, в подземных нефтеносных пластах те же углеводороды могут сохраняться миллионами лет – но там нет физических факторов, за счет которых возможно самопроизвольное усложнение молекул: солнечного излучения, неравномерного или переменного во времени нагрева и т. п. Так что приходится признать: первейшее и непременное условие зарождения жизни – отсутствие таковой.
Из этого вывода, кстати, следует еще одно простое и довольно грустное соображение. Как мы знаем, у всех известных нам живых организмов фундаментальная биохимическая основа одна и та же: полинуклеотиды и полипептиды (белки). Причем в этих соединениях используется один и тот же набор мономеров: 20 основных аминокислот для всех белков и по 4 типа нуклеотидов для каждой из нуклеиновых кислот. И даже генетический код, то есть правила “перевода” нуклеотидной последовательности в аминокислотную, у всех живых организмов один и тот же, с незначительными разночтениями: ген любого организма может быть прочтен молекулярными машинами любого другого организма – и прочтен правильно. Трудно представить, чтобы этот набор нуклеотидов и аминокислот был единственно пригодным для формирования простейших самовоспроизводящихся систем. И уж точно ни из каких законов природы или расчетов энергетической выгоды не следует, что правила соответствия между тройками нуклеотидов и аминокислотами должны быть именно такими, каковы они есть, а не иными. Генетический код достаточно произволен – если не абсолютно, то в значительной степени, – что и делает возможным существование отдельных “разночтений” в нем. Его принципиальное единство у всех живых организмов свидетельствует лишь о том, что все они – потомки только одной линии “преджизни”, самовоспроизводящихся молекул. Вполне возможно, что на ранних стадиях становления жизни были и другие подобные системы. Но по каким-то существенным параметрам (скорость и точность самокопирования, устойчивость к тем или иным повреждающим агентам и т. п.) они уступали “нашей” линии – возможно, совсем чуть-чуть, но этого хватило, чтобы в условиях ограниченности ресурсов они в конце концов сошли со сцены и сами стали ресурсом для победителей. А поскольку распавшиеся и утилизированные белки и нуклеиновые кислоты (а тем более вышедший из употребления генетический код) не оставляют различимых следов, мы никогда не сможем точно узнать даже о самом факте существования этих альтернативных вариантов жизни – не говоря уж о том, сколько именно их было и в чем конкретно заключались их отличия от состоявшейся жизни.
Но вернемся к происхождению той единственной жизни, которая нам известна. И для начала оговорим исходные условия. Сегодня мы знаем, что на поверхности древней Земли, по крайней мере, в некоторых местах, не только мог, но и должен был возникать обширный набор простых органических молекул – сахарá, аминокислоты, азотистые основания и многое другое. (О том, что это были за места, какие факторы обеспечивали абиогенный синтез органики и в каких именно реакциях она возникала, можно прочитать в книге Никитина.) Но от таких органических “кирпичиков” до самых примитивных клеток – дистанция огромного размера. Причем этот разрыв надо было не просто чем-то заполнить (хотя бы мысленно), но еще и как-то решить или обойти целый ряд теоретических проблем, среди которых особенно непреодолимыми выглядели три.
Во-первых, самопроизвольное усложнение и упорядочение молекул как-то плохо согласовывалось с классической термодинамикой – особенно с ее знаменитым Вторым законом. Перипетии отношений эволюционной биологии (и биологии вообще) с термодинамикой мы будем специально рассматривать в главе “Эволюция и энтропия”, здесь же скажем лишь, что прямого запрета на усложнение химических структур термодинамика на самом деле не налагает. Но из этого следует лишь принципиальная возможность такого усложнения – но не его конкретные причины и механизмы. В конце концов, те же законы термодинамики допускают существование паровоза, но это не означает, что в мире, где действуют эти законы, паровозы могут появляться сами собой. Они появились лишь тогда, когда человек их придумал и начал делать. Так что для окончательного разрешения этой трудности нам нужно указать фактор или факторы, под действием которых органические молекулы постепенно усложнялись.
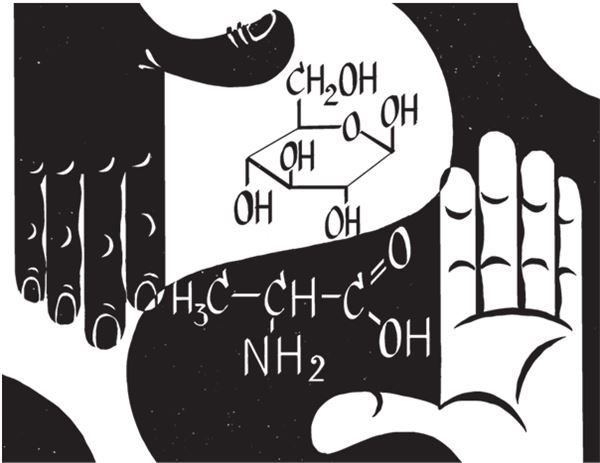
Второй трудностью стала проблема хиральной чистоты. Дело в том, что многие органические молекулы (все, в которых хотя бы один атом углерода соединен с четырьмя разными атомами или атомными группами) существуют в двух вариантах, каждый из которых – как бы зеркальное отражение другого, как русское И и латинское N. Попробуйте, не поднимая букву И с поверхности, повернуть ее так, чтобы получилась N – у вас ничего не получится. А с молекулами, содержащими асимметричный атом, этого не удастся сделать и в трехмерном пространстве. Это свойство молекул называют хиральностью (от древнегреческого “хейр” – “рука”: молекулы разной хиральности точно отражают друг друга, но не могут быть совмещены никакими поворотами в пространстве, как кисти правой и левой руки). В живых клетках каждое такое вещество представлено молекулами только одной хиральности: все входящие в состав белков аминокислоты – “левые”, все сахара – “правые”. В то же время когда такие вещества образуются в “обычных” (протекающих без участия ферментов) химических реакциях, получается смесь из одинакового числа “правых” и “левых” молекул. В биохимических реакциях такого не бывает никогда: их катализируют белки-ферменты, а каждый фермент может работать только с молекулами одной хиральности. Но сами ферменты, как и все белки, состоят только из “левых” аминокислот. Значит, первые живые системы сумели как-то разделить “зеркальные” молекулы еще до того, как обзавелись ферментами. Как же им это удалось и почему это вообще произошло?
Наконец, третья загадка – это проблема возникновения белково-нуклеиновой кооперации. Как мы помним, белки в природе образуются только в живых организмах в результате сложного процесса белкового синтеза, для которого необходима матричная РНК – носитель информации о последовательности аминокислот в будущем белке, – а также транспортные и рибосомные РНК. Никаким иным путем, без участия нуклеиновых кислот, белки возникать не могут. Сами же нуклеиновые кислоты синтезируются матричным образом друг на друге – но они могут это делать только с помощью белков. Процесс синтеза белка тоже требует участия уже готовых белков – ферментов аминоацилтрансфераз и белков рибосомы. Но не могли же белки и нуклеиновые кислоты появиться одновременно в случайных химических реакциях – сразу готовые взаимодействовать друг с другом! А если один из этих классов веществ появился раньше другого – как же он обходился без него? Нетрудно видеть, что по сути дела это старая схоластическая проблема “что появилось раньше – курица или яйцо?”, сформулированная в понятиях молекулярной биологии и оказавшаяся неожиданно актуальной для науки о происхождении жизни.
У теорий, пытавшихся как-то объяснить возникновение жизни из неживых химических компонентов, возникали и другие трудности. Известно, например, что соединение относительно простых органических молекул в основные биополимеры (аминокислот – в белки, нуклеотидов – в нуклеиновые кислоты) идет с выделением воды. Элементарные принципы общей химии гласят, что в водных растворах, где вода заведомо в избытке, равновесие таких реакций будет сдвинуто в сторону не полимеризации, а гидролиза полимеров – то есть самопроизвольное возникновение сколько-нибудь длинных (хотя бы в несколько десятков звеньев) цепочек в таких условиях совершенно невероятно. Между тем у всех известных нам форм жизни эти процессы протекают исключительно в водной среде. Там их обеспечивают опять-таки специальные ферменты, затрачивая на это энергию. Фактически присоединение каждого звена представляет собой целую цепочку промежуточных реакций, точно состыкованных между собой. Но как и за счет чего это могло происходить на древней Земле?
Еще одна чисто химическая трудность состояла в том, что фосфор в неживой природе встречается только в виде химически инертных и почти не растворимых соединений вроде фосфата кальция. Между тем этот элемент абсолютно необходим для всех известных нам форм жизни: он входит в состав ДНК и РНК, его соединения используются как источник энергии для любых реакций, требующих энергетических затрат и т. д. Откуда брали его ранние живые или “протоживые” системы?
Трудности на пути теории абиогенеза (так стали называть все версии возникновения жизни, основанные на предположениях, что это происходило на Земле и исключительно посредством естественных процессов) были настолько велики, что многие – в том числе и выдающиеся умы – обратились мыслями к альтернативной возможности: жизнь не возникла на Земле, а была занесена на нее извне. Сторонниками этой гипотезы, получившей название теории панспермии, были Сванте Аррениус, Владимир Вернадский, Фрэнсис Крик; особенно горячо защищал ее выдающийся британский астрофизик Фред Хойл (с которым нам еще предстоит встретиться в следующей части этой книги). А ученик и постоянный соавтор Хойла Чандра Викрамасингхе и созданная им научная школа до сих пор пытаются найти хоть какие-то свидетельства внеземного происхождения жизни.
Не вдаваясь в подробности аргументации сторонников панспермии, заметим только, что она не объясняет возникновение жизни, а лишь переносит его с древней Земли (о которой нам известно хоть что-то) в абсолютно неведомое место с неизвестными условиями. При этом легко видеть, что ни одной из упомянутых выше трудностей теории абиогенеза теория панспермии не решает. Термодинамическая невероятность самопроизвольного усложнения химических систем, хиральная чистота химических “кирпичиков” жизни, происхождение белково-нуклеиновой кооперации и даже полимеризация белков и нуклеотидов в водной среде выглядят на комете, на Марсе, на спутниках Юпитера, в газо-пылевых скоплениях, в окрестностях нейтронных звезд и черных дыр и в любых других условиях, которые только может нарисовать человеческое воображение, столь же загадочно и необъяснимо, как и на Земле.
Некоторые ученые (включая Вернадского) за неимением лучшего предположили, что жизнь вообще не возникала – это, мол, изначальное и неотъемлемое свойство вечно существующей материи. Но в середине ХХ века в космологии возобладало представление, что сама Вселенная имеет начало – Большой взрыв. И сразу после ее возникновения физические условия были явно несовместимы с существованием жизни – по крайней мере, той, которая нам известна. Так что эта жизнь все-таки должна была когда-то и где-то возникнуть.
Ситуация выглядела настолько тупиковой, что в последние десятилетия ХХ века сама тема происхождения жизни стала восприниматься как не вполне научная. Некоторые журналы отказывались принимать какие-либо публикации на эти темы. Стали слышны даже разговоры о том, что эта проблема, мол, вообще не может быть изучена научными методами – наука, дескать, занимается только явлениями повторяющимися и воспроизводимыми, а жизнь нам известна только одна и она явно возникла один раз. И даже если мы когда-нибудь сможем предложить процесс, приводящий к возникновению жизни из неживых компонентов – это докажет лишь, что жизнь могла возникнуть таким путем, но мы никогда не сможем быть уверены, что именно таким путем она и возникла…

Отборные молекулы
Однако, как ни странно, именно в этот период начались первые подвижки в преодолении вышеупомянутых принципиальных трудностей. В начале 1980-х годов Томас Чек в Университете Колорадо и независимо от него Сидни Олтмен в Йельском университете обнаружили, что некоторые молекулы РНК обладают ферментативной активностью, то есть способны катализировать те или иные химические реакции. С легкой руки Чека и его сотрудников такие РНК стали называть рибозимами. Это уже давало надежду на выход из замкнутого круга “нуклеиновые кислоты – белки”: если рибозимы могут катализировать какие-то реакции (в том числе и с участием других молекул РНК), то, может быть, они могут катализировать и копирование самих себя? И действительно, среди рибозимов (которых вскоре открыли и синтезировали довольно много) нашлись и способные катализировать репликацию РНК. Правда, рибозимы, которые могли бы копировать сами себя (или друг друга) непосредственно из отдельных нуклеотидов, до сих пор не найдены и не созданы, но принципиальная возможность их существования уже выглядит вполне правдоподобной. Так, в январе 2020 года группа Джеральда Джойса из Солковского института в Ла-Хойе (Калифорния) сообщили о получении рибозима из примерно двух сотен нуклеотидов, способного синтезировать собственного “предка” – вдвое более короткую молекулу РНК, содержащую участок, образующий каталитический центр, и тоже проявляющую рибозимную активность. Следует отметить, что исходный рибозим был получен путем “направленной эволюции”, т. е. размножения молекул и отбора наиболее эффективных из них.
Надо сказать, гипотезу о том, что РНК может обладать каталитической активностью, выдвигали еще в 1967 году Фрэнсис Крик, Лесли Оргел и Карл Вёзе. Вскоре Вёзе постулировал существование “РНК-мира” – мира самокопирующихся рибозимов. Предполагалось, что некоторые из небольших молекул РНК, образовавшихся в результате чисто случайного соединения абиогенно синтезированных нуклеотидов, обладали рибозимной активностью. Она была весьма разнообразной, но те РНК, которые могли хоть немного содействовать матричному копированию самих себя, росли в числе. Таким образом, естественный отбор начал работать еще на этом этапе. Который можно считать рубежом становления жизни или относить еще к “химической эволюции”, но в любом случае “демон Дарвина” взялся за работу задолго до появления даже самых примитивных клеток и вообще всего, в чем мы могли бы опознать живой организм. Со временем РНК передали свои каталитические функции белкам, а длительное хранение наследственной информации возложили на свою модифицированную форму – ДНК. Еще позже они обросли системами обмена веществ, закутались в липидные пузырьки-мембраны (позволяющие поддерживать нужную концентрацию реагентов и промежуточных продуктов), создали механизм фотосинтеза… в общем, дальше это была уже знакомая нам биологическая эволюция.
Разумеется, РНК-мир – это всего лишь гипотеза. В современных работах по проблеме происхождения жизни (с наступлением XXI века начался настоящий бум исследований в этой области, как теоретических, так и экспериментальных) можно найти и альтернативные сценарии перехода от химической эволюции к биологической (например, подход “сначала метаболизм”, согласно которому первым шагом к жизни были не самовоспроизводящиеся молекулы, а самовоспроизводящиеся циклы химических реакций, в которых участвуют вещества, выступающие одновременно реагентами, продуктами и катализаторами), и дополнительные гипотезы. Сама концепция РНК-мира сегодня далеко ушла не только от догадки Карла Вёзе, но и от той куда более проработанной версии, которую придал ей в 1986 году другой известный молекулярный биолог – Уолтер Гильберт. Сегодня это глубоко проработанная теория, в рамках которой, в частности, получили изящные объяснения некоторые трудные моменты.
Как мы помним, в водном растворе свободные нуклеотиды должны быть не склонны к соединению в цепочки. Чтобы их соединить, нужно затратить энергию. Если раствор подвергается воздействию солнечного света, эта реакция может идти за счет энергии ультрафиолетовых квантов. Но ультрафиолет с таким же успехом может и разрушить молекулу нуклеотида, поглотившего квант. Так вот, оказалось, что соединение рибонуклеотидов в цепочки повышает их устойчивость к ультрафиолету: даже в коротких цепочках по 3–5 звеньев нуклеотиды устойчивее, чем в свободном состоянии, и эта устойчивость растет (правда, с постепенным замедлением) с ростом числа звеньев в цепочке. Но этот эффект возникает только в том случае, если все нуклеотиды в цепочке – одной хиральности (все “правые” или все “левые”). Поскольку устойчивость достигается за счет распределения вызванного квантом возбуждения электрона по “стопке” расположенных параллельно друг другу плоских азотистых оснований, а для такого их расположения нужно, чтобы цепочка нуклеотидов закручивалась в правильную спираль.
Это позволяет разом ответить на два трудных вопроса: о причинах роста цепочек (вопреки требованиям химического равновесия) и о причинах их хиральной чистоты. Под действием ультрафиолета нуклеотиды постоянно соединяются друг с другом – и тот же ультрафиолет постоянно рвет возникающие цепочки и разбивает на части сами нуклеотиды. В этих постоянных перестановках преимущественно “выживают” те композиции молекул, которые, однажды возникнув, труднее всего разрушаются – небольшие, хирально чистые молекулы РНК.
Один из ведущих специалистов по проблеме происхождения жизни, известный биофизик Армен Мулкиджанян считает, что это не единственное селективное действие ультрафиолетового излучения на предбиологические системы. Возможно, уже сам наш генетический “алфавит” – набор азотистых оснований, входящих в состав ДНК и РНК и служащих непосредственными носителями генетической информации, – тоже выбран для нас ультрафиолетом. В принципе соединений такого типа существует довольно много, немало из них может входить в состав нуклеотидов, объединяться в цепочки и образовывать уотсон-криковские пары с другими основаниями. Но именно те, которые используются в наших “молекулах наследственности”, – аденин, гуанин, цитозин и тимин/урацил – при захвате ультрафиолетового кванта разрушаются реже всех других известных типов нуклеотидов. Устойчивость к ультрафиолету повышается также при образовании комплементарных пар. Поскольку именно этот процесс лежит в основе матричного синтеза нуклеиновых кислот (свободные нуклеотиды образуют пары с соответствующими им нуклеотидами одноцепочечной ДНК или РНК, выстраиваясь таким образом во вторую цепочку), то можно предположить, что и удвоение древних РНК могло начаться еще до того, как какая-то из них обрела способность катализировать эту реакцию – просто за счет того, что двухцепочечные РНК оказывались более устойчивы к разрушающему действию ультрафиолета, чем такие же одноцепочечные. Конечно, и по скорости, и по точности соблюдения соответствия между парами такой механизм, вероятно, сильно уступал рибозимам-полимеразам (не говоря уж о современных полимеразах-белках), но сам феномен самовоспроизводящихся молекул вполне мог начаться с него.
Я думаю, читатель уже понял, к чему я клоню. Да, все вышеописанное – это явная (хотя и несколько своеобразная) версия дарвиновского отбора. Который, оказывается, начал действовать не с момента появления жизни (хотя бы даже и в виде молекул, способных копировать себя), а гораздо раньше. И по сути дела создал эту жизнь, собрав ее из абиогенно синтезированных малых органических молекул и наделив свойствами, выходящими за пределы собственно химии.
Конечно, это всего лишь гипотеза, причем даже возможности ее проверки довольно ограничены. Но сегодня это, пожалуй, самое изящное и убедительное объяснение перехода от малых органических молекул к самокопирующимся биополимерам и дальнейшей эволюции жизни. Она обходится без привлечения “излишних сущностей” (будь то “разумный замысел”, инопланетяне-экспериментаторы, неведомые физико-химические условия на экзотических космических объектах или никем не найденные чудо-катализаторы), рассматривая только те факторы, наличие которых на древней Земле не вызывает сомнений. И при этом не противоречит никаким известным фактам.
Заметим, что уже сам факт существования такой модели (независимо от того, подтвердят ли ее дальнейшие исследования) прямо опровергает дежурную претензию креационистов и всякого рода мыслителей широкого профиля – “дарвинизм бессилен объяснить возникновение жизни”. Можно (если, конечно, есть серьезные аргументы) оспаривать предложенное объяснение, но ни о каком “бессилии объяснить” говорить уже не приходится.
Здесь, впрочем, надо сделать важную оговорку. Объяснение происхождения жизни подразумевает решение целого ряда проблем, среди которых проблема возникновения самокопирующихся молекул РНК – безусловно важная, одна из ключевых и, возможно, самая принципиальная, но все же не единственная. Чтобы можно было без натяжки сказать, что загадка возникновения жизни разгадана, нужно выяснить, где разыгрывались события, приведшие к ее возникновению (имеется в виду, конечно, не “в какой географической точке?”, а “в какой среде?”, “в каких локальных физико-химических условиях?”), как соединились воедино самокопирование молекул и метаболические циклы, как возникли белки и ДНК, генетический код и рибосомы, как ансамбли взаимодействующих молекул оделись мембранами и как научились управлять делением мембранных пузырьков, и многое-многое другое. Ученые выдвигают разные, порой противоречащие друг другу гипотезы и модели, спорят, экспериментируют. Контуры моста над пропастью, разделяющей живое и неживое, уже вырисовываются, но никто не может сказать, когда по этому мосту можно будет пройти от одного края до другого, ни разу не прибегнув к крыльям воображения.
Но, как я уже сказал в начале этой главы, у меня нет ни возможностей, ни нужды в полноценном рассказе об этой безусловно интереснейшей области исследований. Желающим узнать о ней подробнее я опять-таки рекомендую книгу Никитина. Здесь же я хотел лишь показать, что и в этом весьма специфическом направлении исследований идея эволюции на основе естественного отбора присутствует не только как некое абстрактное универсальное объяснение, но и как актуальная рабочая теория, источник возможных решений для конкретных (и притом важнейших) исследовательских задач.

Поэтому из всего необозримого множества тех проблем, над которыми бьются ученые, занимающиеся происхождением жизни, я позволю себе сказать пару слов только еще об одной. Во-первых, потому, что она была упомянута в начале этой главы (как известно, если в первом акте на сцене висит ружье, до конца пьесы оно должно выстрелить), во-вторых, потому, что здесь мы вновь встречаем пример успешного применения селекционистского подхода.
Итак, открытие рибозимов и создание концепции РНК-мира разорвало замкнутый круг “белки не могли возникнуть без нуклеиновых кислот, а нуклеиновые кислоты – без белков”. Но если нуклеиновые кислоты смогли возникнуть и какое-то заметное время успешно существовать без всяких белков – как и почему эти два типа биополимеров начали взаимодействовать между собой?
Когда ученым удалось узнать кое-что об эволюции рибосомы (об этой замечательной работе мы еще поговорим подробнее в главе 20), оказалось, что самая древняя часть рибосомальной РНК – пептидилтрансферазный центр. Этот участок непосредственно пришивает аминокислоты к растущей полипептидной цепочке. Но в его составе нет никаких структур, которые проверяли бы соответствие присоединяемой аминокислоты тому, что значится в матричной РНК, да и вообще он с мРНК никак не взаимодействует. Возможно, конечно, древний рибозим, превратившийся впоследствии в пептидилтрансферазный центр, контролировал состав получающейся полипептидной цепочки каким-то иным способом, позже утраченным за ненадобностью. Но более вероятно, что он его никак не контролировал, а сшивал аминокислоты (правда, строго одной хиральности – “левые”) в том порядке, в каком они ему подворачивались.
Но какую пользу размножающимся и каталитически активным молекулам РНК мог бы принести белок случайного состава? Одна из гипотез связывает это с тем, что полипептидная цепочка случайного состава с высокой вероятностью закручивается в альфа-спираль – наиболее частый элемент вторичной структуры белка. В таком виде полипептид охотно связывается водородными связями с двухцепочечными участками РНК, прикрывая при этом ее “ахиллесову пяту” – гидроксильную группу у второго атома углерода в молекуле рибозы. Едва ли не любые химические агенты, способные гидролизовать РНК – от щелочи до специализированных ферментов-нуклеаз, – атакуют именно эту группу. (Недаром в дезоксирибозе, входящей в состав ДНК, именно этот гидроксил заменен атомом водорода – что придает ДНК гораздо бóльшую, чем у РНК, устойчивость к химическим повреждениям.) Нетрудно предположить, что когда самовоспроизводящиеся рибозимы “подъели” абиогенно синтезируемые нуклеотиды, и те стали дефицитным ресурсом, преимущество получили рибозимы, способные гидролизовать собратьев – другие молекулы РНК. А это, в свою очередь, породило отбор на способность защищаться – и эффективным средством такой защиты оказались полипептидные цепочки случайного (но хирально чистого) состава. Таким образом, первый шаг к кооперации полинуклеотидов с полипептидами тоже мог быть результатом естественного отбора.
На чем эту главу можно было бы и закончить. Но есть еще один сюжет, косвенно связанный с ранней эволюцией жизни и тоже окруженный странными недоразумениями. Правда, эти недоразумения не столь широко распространены в обществе, но зато осенены именами действительно выдающихся ученых нашего времени.

