Книга: Креативный мозг. Как рождаются идеи, меняющие мир
Назад: Значимость по умолчанию
Дальше: Разбавленная значимость
Значимость, дофамин и активация лобных долей – или отсутствие всего этого
Сигнальная система значимости
В механизме мозга, определяющем значимость, есть еще одна, биохимическая «шестеренка», и это дофамин. Это вещество обычно упоминается в научной литературе как DA и является нейромодулятором. В головном мозге действуют несколько биохимических систем, играющих важнейшую роль в сообщении между нейронами. Среди этих биохимических систем обычно выделяют быстродействующие нейромедиаторы и медленно действующие нейромодуляторы (хотя и те и другие часто называют «нейромедиаторами»). Дофамин принадлежит ко второй группе. Дофамин единственный из всех нейромедиаторов, который привлекает интерес широкой публики, поскольку он часто упоминается в бульварной прессе. Если есть такая вещь, как «модный» нейромедиатор, то это дофамин. Он причастен к образованию зависимости, развитию синдрома дефицита внимания и гиперактивности (СДВГ) и многих других состояний. Его точная функция (равно как и дисфункция) интересует как научную, так и широкую общественность. Дофамин называют «гормоном удовольствия» и «медиатором вознаграждения» – все это яркие обозначения, порождающие лишь иллюзию понимания, нежели истинное понимание.
Но дофамин – это просто химическое вещество, которое само по себе не управляет никакими процессами в нервной системе. Его функция заключается в облегчении процессов, протекающих в этих структурах. Чтобы разобраться в функции (а также дисфункции) дофамина, важно изучить анатомию его путей распространения и взаимоотношения тех структур, сообщение между которыми облегчает дофамин. Патриция Голдман-Ракич, исследования которой мы уже обсуждали в Главе 4, была среди тех, кто первым распознал роль дофаминергических путей в работоспособности префронтальной коры, а также ее дисфункцию в случае нарушения передачи дофамина.
Важное скопление дофамина находится в так называемой вентральной области покрышки (ВОП). Расположенные здесь нейроны посылают дофаминовые проекции в ряд структур мозга, в том числе в префронтальную кору, гиппокамп, миндалину и прилежащее ядро. Повреждение этих областей изменяет и процесс принятия решений (который обычно ассоциируется с префронтальной корой), и память. Эти проекции, исходящие из ВОП, привлекают внимание ученых, и я ранее изучал их роль в регуляторной функции и долгосрочной памяти13. Повреждение ВОП и ее двунаправленных проекций, к префронтальной коре и от нее, приводит к клиническому состоянию, практически не отличимому от повреждения лобных долей. Различие, вероятно, заключается в том, что малейшее повреждение этих путей, которое может быть почти неразличимо при помощи обычных методов нейровизуализации, приводит к неуловимым поведенческим и аффективным последствиям «легкой» черепно-мозговой травмы, которыми часто пренебрегают как «изменениями личности». Среди прочего, усовершенствования в выявлении такого повреждения помогут нам распознать различие между последствиями двух состояний, которые часто объединяются в клиническом диагнозе черепно-мозговой травмы (ЧМТ), при которой, вероятно, нарушаются эти пути, и синдрома посттравматического стресса (СПТС), с сохранением их целостности. Мои коллеги и я описали последствия структурного повреждения ВОП и ее проекций много лет назад, и мы назвали это состояние «синдромом ретикуло-фронтального разъединения»14. Истощение запасов дофамина в лобных долях проявляется так же, как «синдром ретикуло-фронтального разъединения», и тоже приводит к нарушению функций лобных долей15.
Но, чтобы разобраться в функциях дофамина, не менее важно рассмотреть проекции, которые приходят к ВОП. Некоторые такие проекции исходят из префронтальной коры и миндалины. Мы уже знаем, что префронтальная кора чрезвычайно важна для определения значимости, равно как и миндалина. Разница между этими двумя структурами состоит в том, что префронтальная кора определяет значимость на основе когнитивной обработки, а миндалина – эмоционального восприятия.
Давайте предположим, что префронтальная кора или миндалина распознают некий стимул или информацию как значимые, важные для преуспевания организма или предотвращения травмы. Они посылают сигнал в ВОП через нисходящие пути, и этот сигнал означает, что ВОП помещает стимул или информацию, так сказать, в «начало нейронной очереди». В ответ на это ВОП посылает сигнал через восходящие пути в неокортекс, прилежащее ядро, гиппокамп и другие структуры, отмечающие этот сигнал (или информацию) как особенно существенный, или «значимый», привлекая тем самым к нему внимание, «побуждая» организм сосредоточиться на нем, а также облегчая его перенаправление в долгосрочную память, для сохранения и закрепления. Таким образом, дофамин является переносчиком, нежели автором сообщения, исходящего из префронтальной коры или миндалины. Выброс дофамина из ВОП отмечает определенные события или частицы информации как «важные». Возможно, не будет слишком самонадеянно предположить, что студенты, которые в ранее приведенном примере слушали лекцию и в реальном времени выбирали ту информацию, которую стоило записать в конспект, действовали под влиянием выбросов дофамина. Распределение главных путей дофамина в мозге изображено на рис. 5.2.
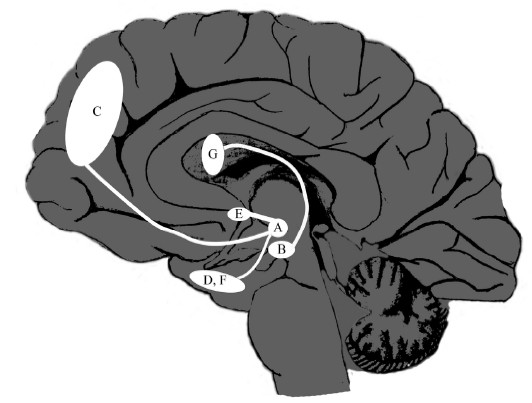
Рис. 5.2. Дофаминергические пути. (А) Вентральная область покрышки (ВОП); (В) Черная субстанция; (С) Префронтальная кора; (D) Миндалина; (E) Прилежащее ядро; (F) Гиппокамп и парагиппокампальные структуры; (G) Полосатое тело. Сеть связей между A, C, D и Е особенно важна для определения значимости
Различные компоненты дофаминовой системы играют разные роли в обучении, направляемом значимостью. При помощи методов позитронной эмиссионной томографии (ПЭТ) команда ученых из Университета Макгилла в Монреале пришла к выводу, что поведение приближения, основанное на положительном опыте, и поведение избегания, основанное на отрицательном, связаны с двумя разными рецепторами дофамина в кортикостриарной системе – соответственно D1 и D2. Важное значение этой работы заключается в том, что вариабельность связывания рецепторов D1 и D2 может объяснять индивидуальные различия в обучении в зависимости от стилей воприятия16.
Карл Дейссерот и его коллеги из Стэнфордского университета чрезвычайно элегантно продемонстрировали значение специфических дофаминовых рецепторов в процессе принятия решений, направляемом значимостью. В эксперименте крысы должны были выбирать между двумя рычагами: один был связан с доступом к кормушке, где всегда был корм, а другой – с доступом к кормушке, где корма обычно почти не было, но иногда появлялось очень много. Ученые обнаружили, что способность крыс учиться на собственном опыте и делать выбор направлялась сигналом, исходящим из нейронов прилежащего ядра с рецепторами D2. Это область мозга на границе вентрального полосатого тела, передней части поясной извилины и орбитофронтальной коры. Известно, что в этой области располагается центр системы вознаграждения мозга. Те крысы, у которых эти нейроны посылали сильный сигнал, учились на отрицательном опыте и использовали консервативную стратегию, чаще всего выбирая рычаг, который открывал кормушку, где всегда был корм. И наоборот, крысы, у которых нейроны с рецепторами D2 вырабатывали слабый сигнал, продолжали нажимать на рычаг, чтобы время от времени получить доступ к кормушке, где иногда появлялось много корма, хотя чаще всего его количество было очень ограниченным. Но эту упрямую забывчивость по отношению к отрицательному опыту можно было преодолеть у этих склонных к риску крыс, если стимулировать нейроны с рецепторами D2 в прилежащем ядре оптогенетическим методом. (Это относительно новый и многообещающий метод, когда нейроны подвергают генетической модификации, чтобы они приобрели чувствительность к свету и отвечали на стимуляцию светом17.)
Сигнальная система дофамина выполняет другую важную функцию: это часть системы модуляции возбуждения в мозге. «Значимый» сигнал, передаваемый дофамином, модулирует префронтальную кору, способствуя ее переходу в оптимальное возбужденное состояние, о котором мы говорили в Главе 4 и к обсуждению которого вернемся в Главе 7. Основываясь на ранних работах Патриции Голдман-Ракич, Эми Арнстен и ее коллеги показали, что вызванная дофамином и другим нейромедиатором, норадреналином, модуляция возбуждения играет важнейшую роль в функции префронтальной коры и что сбой этой модуляции приводит к самым различным когнитивным расстройствам18.
Я полагаю, что участие и дофамина, и норадреналина в возбуждении лобных долей отражает два дополняющих типа модуляции возбуждения: один основан на воспринимаемой важности стимула (дофамин) и опосредует использование хорошо установленных привычных когнитивных задач с ранее установленной значимостью. Другой основан на новизне стимула (норадреналин) и задании, которое требует умственной гибкости, а также отклонения от ранее установленной когнитивной привычки. Логическое обоснование этого предположения можно найти в моей предыдущей книге, «Новый управляющий мозг»19. Другим его подтверждением является дополняющее влияние норадреналиновой и дофаминовой модуляции на когнитивные функции. Фармакологическая модуляция норадреналиновой системы улучшала результат выполнения испытуемым ряда сложных, новаторских вербальных заданий, требующих гибкости мышления, таких как решение анаграмм и выполнение теста на беглость речи. И наоборот, введение бромокриптина (агониста дофамина) не вызывало такого эффекта20. С другой стороны, модуляция дофаминовой, но не норадреналиновой системы облегчает выполнение лексических задач, основанных на автоматической обработке и требующих хорошо установленных семантических отношений21.
Когда нейробиологи говорят о степени умственной гибкости, необходимой для выполнения задания, они делают различие в переключении между однородными признаками стимула и разнородными и обработкой данных. Похоже, что дофаминергическая система играет более важную роль в относительно ограниченном переключении между однородными признаками, которое открывает переход между стимулами одного рода, тогда как норадренергическая система связана с переключением между различными признаками, с более свободным переходом между разнородными стимулами22.
Чуть позже мы обсудим динамическое взаимодействие различных состояний активации лобных долей, то есть гипер- и гипофронтальности, которые являются неотъемлемой частью творческого процесса и способности к созданию новых идей. Емкость мозга для достижения и поддержания широкого диапазона префронтальной активации и деактивации представляется чрезвычайно желательной характеристикой и важным ингредиентом комплексного познания. В той степени, в которой этот диапазон может варьировать у разных людей, индивидуальные различия могут проявляться как различия в свойствах вентральных ядер ствола головного мозга и их проекций, выходящих из голубого пятна, которое участвует в синтезе норадреналина, и ВОП, где происходит синтез дофамина. В Главе 7 я докажу, что чем шире диапазон модуляции, тем это лучше для инноваций и креативности.
И снова о дофамине
Значение взаимодействия префронтальной коры и вентрального ствола головного мозга для обеспечения оптимального процесса познания стало понятным благодаря работам Такеуши и его коллег из Университета Тохоку в Японии. При помощи обычного метода измерения величины различных структур мозга – МРТ воксельной морфометрии – ученые показали, что индивидуальные различия в выполнении тестов на «креативность» (так называемые тесты на «дивергентное мышление») были связаны с размером правой дорсолатеральной префронтальной коры и ВОП. То есть лучшие результаты соответствовали более крупному размеру этих структур23. Опять – «чем больше, тем лучше»!
Между уровнями дофамина и психологическими характеристиками разных людей была обнаружена взаимосвязь, но ее природа пока остается предметом обсуждения. Предполагается, что высокие уровни дофамина связаны с более выраженной психической ригидностью и ограниченной гибкостью мышления. Наоборот, низкие уровни дофамина соответствуют ослабленной ригидности и повышенной гибкости, а также стремлению к новизне24. Результаты нескольких исследований с участием людей и животных поддержали эту идею: повышение уровня дофамина в результате фармакологических вмешательств вызвало повторяющееся стереотипное поведение25. В ранее описанной работе Карла Дайссерота и его коллег у крыс наблюдались индивидуальные различия в сигналах, исходящих из нейронов D2 прилежащего ядра. Различия в силе сигналов соответствовали различиям между консервативным и рискованным стилями принятия решений26.
И все же научные публикации противоречат друг другу. Другие исследования демонстрируют связь между дофаминергической системой и поиском новизны27. Подобные противоречия могут отражать различную степень участия многих структур мозга, которые испытывают влияние манипуляций с уровнями дофамина. Есть и другое мнение, это может показывать, что такие компоненты поведения, как поиск новизны и направляемый значимостью поиск вознаграждения, могут быть очень тесно переплетены. Тогда невозможно различить, на какой из этих двух компонентов больше влияет фармакологическое воздействие. В экспериментах на животных, поведение которых по определению направляется вознаграждением, особенно сложно распознать это различие.
Более глубокое представление о роли дофамина в поддержании адаптивного баланса между стабильностью и гибкостью психики отмечается между тонической и фазической активацией в дофаминергической системе. Тоническая активация обеспечивает стабильность, а фазическая – пластичность. Различные рецепторы дофамина могут участвовать в проявлении тонических и фазических эффектов (это рецепторы D1 и D2 соответственно). Также возможно, что различные аллели (варианты) фермента КОМТ (ингибитор катехол-О-метилтрансферазы), играющего некоторую роль в распределении дофамина и норадреналина, могут по-разному влиять на этот баланс28.
Оказывается, что левое полушарие более богато дофаминовыми путями, чем правое (см. рис. 5.2), и эта асимметрия наблюдается у нескольких видов млекопитающих, включая наш собственный вид29. Похоже, что несколько важных элементов механизмов, которые мозг использует для определения значимости, более выражены в левом, чем в правом полушарии. Эти важные элементы включают орбитофронтальную кору и переднюю поясную кору (см. рис. 5.1), а также дофаминовые пути, особенно идущие к лобным долям. Эти данные подтверждают, что левая префронтальная система и ее пути особенно тесно вовлечены в познание, направляемое внутренней значимостью.
Назад: Значимость по умолчанию
Дальше: Разбавленная значимость

