Книга: Креативный мозг. Как рождаются идеи, меняющие мир
Назад: Как рождается новая идея?
Дальше: «Головоломка» оперативной памяти
Что стоит за метафорой – макроструктура
Конечно, лобные доли в качестве Мастера Лего и созидательный процесс инноваций как перегруппировка Лего – это просто метафоры. Они могут выражать общую идею о возникновении новых решений из старого опыта, но метафоры ничего не говорят нам о том, какие процессы на самом деле протекают в мозге. Готовы ли вы идти дальше, в глубь метафоры, и кинуть взгляд на нервный механизм инноваций? Мы готовы попробовать. Есть ли у нас всестороннее понимание механизмов? Нет, по крайней мере еще нет, но мы начинаем задавать наводящие вопросы. Их можно задавать на разных уровнях: на макроуровне целого мозга, всех структур и их взаимодействий и на микроуровне сложных нервных цепей.
«Канонические» сети
Как же префронтальная кора взаимодействует с другими структурами мозга в процессе выполнения сложного задания познавательного характера? По самой своей природе такое взаимодействие нельзя понять, если просто рассматривать отдельные структуры мозга; необходимо рассматривать сети. Переход от исследования функций отдельных областей мозга к изучению схем интерактивных мозговых структур (то есть сетей) был одним из главных достижений когнитивной нейробиологии последних десятилетий. Сегодня внимание исследователей чаще всего привлекают три сети: центральная управляющая сеть (ЦУС), сеть пассивного режима работы мозга (СПР) и сеть познавательных и эмоциональных функций (СПЭФ). Эти сети схематически изображены на рис. 4.3.
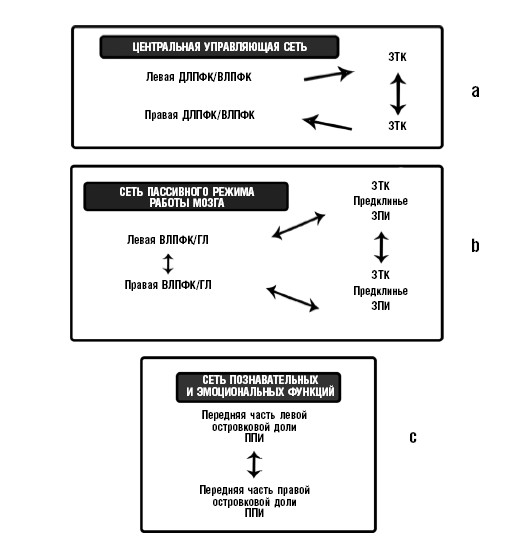
Рис. 4.3. Основные крупномасштабные сети. (а) Центральная управляющая сеть (ЦУС) включает дорсолатеральную и вентролатеральную части префронтальной коры (ДЛПФК и ВЛПФК) и заднюю теменную кору (ЗТК). (b) Сеть пассивного режима работы мозга (СПР) включает вентромедиальную (ВЛПФК) / орбитофронтальную (ОФ) часть коры, заднюю теменную кору (ЗТК), предклинье и заднюю поясную извилину (ЗПИ). (с) Сеть познавательных и эмоциональных функций (СПЭФ) включает переднюю островковую долю и переднюю поясную извилину (ППИ).
Центральная управляющая сеть и сеть пассивного режима изучаются особенно активно, и часто высказываются предположения о четкой границе между ними. Они «антикоррелируют»: когда одна активна, другая пассивна. Каждая сеть состоит из относительно небольшого количества макроскопических «хабов» – областей, известных своими двусторонними связями с особенно большим числом структур и областей мозга11.
Образованный человек, не cведущий в нейробиологии, инженер или математик, может быть обеспокоен склонностью современной когнитивной нейробиологии привлекать эти две сети (ЦУС и СПР) для объяснения слишком широкого круга познавательных процессов. Он также может задать вопрос о том, насколько обоснованно сведение всех возможных взаимодействий, которые происходят в богатой матрице миелиновых волокон головного мозга, всего лишь к нескольким сетям, а также предположение о том, что две крупные сети не могут работать параллельно. На самом деле было обнаружено некоторое количество дополнительных, часто перекрывающихся сетей; например, это сеть внимания, сеть языка и лобно-теменная сеть контроля (ЛТСК). Предполагается также, что две последние сети уникальны для человека, а ЛТСК, в частности, отражает закономерность увеличения коры головного мозга в процессе эволюции, особенно заметного в ее лобной и теменной частях12.
И не успели мы осознать, как старый соблазн вновь заявил о себе под новой личиной: стремление к ограниченному (и желательно не очень длинному) списку четко определенных модулей. Время от времени, в 1970-х и 1980-х годах, нейропсихологи сходили с ума от идеи о том, что кора представляет собой коллекцию «модулей», дискретных объектов с жесткими границами и фиксированными функциями. Это было последнее «усилие» своеобразного состояния дел, при котором наука о познании была в основном отделена от нейробиологии, и слияние этих двух наук произошло к концу двадцатого столетия (по большей части благодаря пришествию методов функциональной нейровизуализации). В результате понятие о «модульном составе коры» было заброшено ради более продвинутого понимания организации мозга. Сегодня, несмотря на то что место областей мозга заняли модули, называемые «сетями», распространение этой идеи поражает жутким сходством с расцветом концепции модулей старой школы, который наблюдался несколько десятилетий назад, и оно отражает ту же самую гносеологическую эстетику (которая никогда особо меня не привлекала). По мере увеличения количества предложенных сетей у них проявляется все большее сходство с классической функциональной системой, смонтированной специальным образом в ответ на существующие познавательные требования, заново открытой при помощи функциональной нейровизуализации13. Исходя из того, что в различных исследованиях нейроанатомии в каждой из этих сетей часто обнаруживается что-то иное, также подтверждает тот факт, что попытки свести все эти наблюдения к небольшому количеству «канонических» сетей являются сомнительным упражнением, результат которого нельзя принимать буквально. Вместо уникальных канонических сетей, возможно, более полезно с эвристической точки зрения думать в терминах «сетевых категорий», представляя, что существует, вероятно, столько сетей, сколько позволяет топология длинных нервных путей мозга. Было бы особенно удивительно, если бы господство этого нового бренда модульности следовало тем же курсом, что и старое понятие о «модульности» областей мозга. Тогда «модульность» сетей мозга ослабнет в свое время ради более тонкого понимания14.
Но, даже в самом грубом приближении, макротаксономия сети «достаточно верна» во многих аспектах в соответствии с состоянием дел в современной нейробиологии, и ее в некотором смысле можно использовать для объяснения. Имеет смысл рассматривать такие сети как крупные системы дорог, которые могут проводить самые разные потоки движения и включать специфические подсистемы. Чуть позже мы более подробно рассмотрим каждую из трех часто упоминаемых сетей.
Центральная управляющая сеть (ЦУС)
Эта сеть, также известная как сеть управления познанием, представляет собой сложное объединение областей мозга, которые совместно работают, когда мы напряженно размышляем, сознательно и целенаправленно, над решением сложной задачи познавательного характера. Вот поэтому сеть часто называют «положительно направленной на задачу». Точное описание компонентов ЦУС может различаться в разных опубликованных работах, что, возможно, объясняется различиями в использованных задачах познавательного характера. Однако латеральная префронтальная кора (дорсальная и вентральная части которой часто объединяются, и эта объединенная часть называется «дорсолатеральной префронтальной корой», или ДЛПФК) и задняя теменная кора (ЗТК) неизменно остаются ее главным компонентом.
Временная динамика в пределах ЦУС проливает свет на взаимоотношения префронтальной коры и задней ассоциативной коры, в которой сложные знания представлены в виде задач, требующих познавательного подхода. Когда ученые изучили порядок, в котором различные компоненты этой сети приходят в активное состояние, стало ясно, что активация внутри сети определяется префронтальной корой15. Сначала активизируется префронтальная кора, а потом другие компоненты ЦУС и участки, которые располагаются в теменных, иногда в височных долях.
Сеть ЦУС может и не быть отличительной особенностью человека. Использование некоторых методов спектроскопии в ближней инфракрасной области и поверхностных потенциалов поля позволило Хоакину Фастеру и его коллегам описать у макак-резусов сеть, анатомически очень близкую к ЦУС, в которой происходит обработка задач оперативной памяти. Для этой сети, объединяющей латеральные префронтальные и задние теменные области, были характерны сложные схемы синхронизации и десинхронизации по времени, частоте и пространству мозга. Это подчеркивает тот факт, что идентификация путей представляет собой только первый шаг к пониманию закономерностей межнейронного сообщения и за этим последует более подробный анализ этих закономерностей16.
Сеть пассивного режима (СПР)
Эта сеть активируется, когда ни одно внешнее задание не запускает познавательные процессы и мозг индивидуума предоставлен самому себе. Иногда такое состояние называется «отрицательно направленным на задачу», но этот термин неверный, поскольку мозг вовсе не бездействует: напротив, вместо того чтобы включаться под воздействием навязанной извне задачи, он занимается заданиями, выбранными внутри и направленными внутрь17.
Мы более подробно рассмотрим сеть СПР в Главе 5, но здесь мы просто сравним две сети – ЦУС и СПР – в отношении их нейроанатомических компонентов. Обе сети сконцентрированы вокруг двух макроскопических хабов: это префронтальная и задняя теменная кора. Их объединенное название – гетеромодальная ассоциативная кора, а их уникальная роль в более сложном познании, обширные связи и особенности работы во взаимодействии были установлены уже довольно давно18. Учитывая это, открытие двух крупномасштабных сетей методами функциональной нейровизуализации стало подтверждением и уточнением хорошо известного, характерного признака функциональной организации коры.
Но пока и ЦУС, и СПР объединяют префронтальную кору и заднюю гетеромодальную ассоциативную кору, их различные участки включены в эту сеть; вот именно это делает их открытие особенно интересным. Самая яркая разница между ЦУС и СПР наблюдается в префронтальной коре. Если в ЦУС активны дорсальная и вентральная части лобной доли, то в СПР активны орбитальная и вентромедиальная части.
Представляется, что задние части ЦУС и СПР частично совпадают, но, тем не менее, между ними есть разница. Обе сети включают гетеромодальную ассоциативную кору задней латеральной теменной доли, но эти участки более интенсивно включены в ЦУС, чем в СПР. И наоборот, СПР включает кору задней части поясной извилины и предклинье (область, которая находится в средней части задней теменной доли), не включенные в ЦУС.
Основываясь на наших знаниях об этих структурах мозга, создается впечатление, что ЦУС обрабатывает информацию о внешнем мире, а СПР – то, что идет изнутри. Обратите внимание, что латеральная поверхность префронтальной коры немного больше в правом, чем в левом полушарии; а орбитофронтальная кора и поясничная извилины (СПР) – наоборот19. Допустим, «чем больше, тем лучше» (грубое, но часто удивительно обоснованное выражение, когда речь заходит о взаимоотношениях структуры и функции в головном мозге); тогда эта асимметрия может служить намеком для выяснения относительной роли двух полушарий в ЦУС и СПР.
Сеть познавательных и эмоциональных функций
Эта сеть обеспечивает взаимодействие ЦУС и СПР. Она была описана в самое последнее время и намного менее изучена. Она состоит из передней части островка и передней части поясной извилины. Представляется, что эта сеть активизируется – особенно островка правого полушария, – когда происходит переключение между сетями СПР и ЦУС. В экспериментах, которые привели к открытию этой сети, переключение было однонаправленным, с СПР на ЦУС, но не обратно, и это переключение запускалось внезапным изменением в сенсорном входящем сигнале – внезапной переменой музыкального тона (темы) или столкновением с редким («оригинальным») стимулом. Поскольку переключение является однонаправленным, то можно утверждать, что оно действительно ускоряется при обнаружении новизны, а не «отличительной особенности», в смысле «значимости» (более подробно о различных толкованиях термина «значимость» и возможностях для его неверного использования в Главе 5). Как мы увидим в Главе 6, правое полушарие играет решающую роль в обработке новизны, что помогает понять асимметричность (правосторонний больше левостороннего) частей островка, участвующих в процессе переключения20.
Другим заслуживающим внимания отличительным признаком процесса переключения является то, что он вызывает не просто активацию ЦУС, но также и одновременную деактивацию СПР (помните, эти две сети «антикоррелируют» – когда одна активна, другая не активна). Это подразумевает, что взаимоотношения сетей носят характер торможения. Как такое происходит? Интересно, что две префронтальные области, которые управляют двумя сетями, проявляют противоположные функциональные свойства в самых разных ситуациях. Повреждение дорсолатеральной коры вызывает симптомы, похожие на депрессию, – именно по этой причине синдром поражения дорсолатеральной части префронтальной коры раньше был известен как «псевдодепрессия», такой термин можно встретить в старой литературе по неврологии. Напротив, повреждение орбитофронтальной области часто приводит к поверхностной эйфории, и этот симптом в шутку называют «мория». Подобным образом при депрессии дорсолатеральная кора часто гипоактивна, а орбитофронтальная – гиперактивна21. Если рассматривать эти данные вместе, то они подтверждают взаимно тормозящие взаимоотношения этих двух областей префронтальной коры.
Центральная нервная система добивается замечательных результатов, используя механизмы взаимного торможения. Может ли быть такое, что, наоборот, «антикоррелирующие» взаимоотношения СПР и ЦУС являются следствием взаимно тормозящих отношений между двумя областями префронтальной коры, которые «управляют» ими? На самом деле латеральная (вентральная и дорсальная) и медиальная части префронтальной коры работают в противоположном режиме: когда одна активна, другая – нет. С одной стороны, это доказывает, что эффективные, взаимно тормозящие, антикоррелирующие взаимоотношения двух крупномасштабных сетей не требуют наличия явно тормозящих, протяженных нервных путей и могут быть следствием динамики глобальной сети22. Этот вопрос остается невыясненным.
В любом случае сеть познавательных и эмоциональных функций может быть переключателем, который обеспечивает эффективное взаимное торможение. На самом деле передняя часть островка взаимосвязана с обширными областями коры, особенно с участками префронтальной коры, в том числе с орбитофронтальной и вентролатеральной областями. Это делает сеть познавательных и эмоциональных функций вероятным переключателем тормозящих взаимоотношений активаторов ЦУС и СПР – латеральной и орбитофронтальной частей префронтальной коры соответственно. Согласно этому предположению, сеть познавательных и эмоциональных функций является скорее «переключателем», чем «переключающей рукой»: как только правое полушарие обнаруживает новизну, экстравертная латеральная префронтальная кора подавляет интравертную орбитофронтальную кору. В результате сеть ЦУС активизируется, а СПР ослабляется.
Предположение об «антикоррелирующих», взаимно тормозящих взаимоотношениях латеральной и медиальной/орбитальной частей префронтальной коры приводит к другому интересному выводу: что каждая из этих двух областей префронтальной коры тяготеет к относительно экстремальным и противоположным уровням активации, избегая средних значений. Иными словами, в любой момент времени одна из этих областей пребывает в состоянии сильного возбуждения, а другая – слабого. Склонность пренебрегать уровнями нервной активации ради относительно экстремальных приводит к концепции «бистабильности» как главной характеристики деятельности латерального и медиального аспектов префронтальной коры. Более подробно об этом в Главе 7.
Назад: Как рождается новая идея?
Дальше: «Головоломка» оперативной памяти

