Магия творчества (факты, соображения, гипотезы)
Современная физиология мозга человека – результат интенсивного развития инвазивных и неинвазивных технологий второй половины и главным образом последних десятилетий XX века. Взаимообогащение и взаимопроникновение идей и достижений физиологии мозга человека и психологии определили реальный базис для попыток проникновения в психофизиологию творчества, его мозговую организацию и мозговые механизмы.
Вклад Института мозга человека РАН в формирование и становление современной физиологии мозга человека вполне существен. По чисто паранаучным причинам он более значителен в первом прорыве XX столетия, когда оказалось возможным с максимальной полнотой судить о событиях, происходящих в микромирах мозга (Бехтерева, 1997), однако и во втором, «технологическом», прорыве наше участие обозначено пионерскими работами именно в области психофизиологии творчества. Подойти к этой сложной, захватывающей, но и одновременно коварной проблеме нам позволил долгий путь изучения мозговой топографической организации и, что очень важно, мозговых механизмов мышления (Бехтерева, 1965–1999). Картирование и микрокартирование мозга при реализации различных функций лишь на первых порах оказывалось основной задачей, сменяясь сверхзадачей изучения механизмов управления мозговой функциональной организацией высших функций. Такому подходу субъективно, по-видимому, способствовали почти случайное открытие нами в 1968 году и последующее осмысление одного из важнейших мозговых механизмов и особенно в том, что касается высших функций, – детекции ошибок (Bechtereva, Gretchin, 1968).
Изучение мозговой организации и мозговых механизмов творчества реально стало осуществимым в настоящую, «технологическую» эру, когда в мозговой физиологии оказалось возможным уйти от поистине сизифова труда примитивных форм анализа данных. Реорганизация физиологических процессов мозга при индуцированном развитии творческого процесса исследуется нами на основе анализа данных локального мозгового кровотока с помощью позитронно-эмиссионной томографии (ПЭТ) и количественной электроэнцефалографии – локальной и пространственной синхронизации. Уникальным изучением творчества делает эти исследования психологический аспект работы, создающий модельную ситуацию вербального творчества и позволяющий далее с помощью адекватных контролей «изымать» из физиологических данных шаг за шагом максимально все, не являющееся собственно творчеством. Конструирование психологических тестов при изучении творчества постоянно совершенствуется и в соответствии с конкретной «подзадачей» и стремлением к «идеалу». Вполне понятно, что выделение физиологических механизмов собственно творчества – задача, на пути решения которой исследователя ожидают далеко не одни удачи.
Это они создавали волшебство. Жанна Нагорнева, Сергей Данько, Маша Старченко, Наташа Шемякина
Тесты на вербальную креативность предъявлялись в наших исследованиях волонтерам на экране монитора в виде черно-белых матриц с наборами различных текстов и с соответствующими словесными заданиями к ним. Опыт предварительных психологических исследований (Старченко, 2000) показал, что испытуемые использовали разную стратегию в решении предъявляемых им психологических заданий. Соответственно, далее использовались две разные (основные) конструкции тестов. Естественно, не исключено, что этими двумя вариантами возможные различия в способах решения вербальных задач не исчерпываются. Далее тесты реконструировались в соответствии с конкретными задачами.
В отличие от дальнейшего изложения материала, описание первых двух типов тестов приводится здесь достаточно подробно.
Первый тест состоял из 4 заданий. В каждом задании испытуемому предъявлялись в течение 90 секунд черными буквами на белом фоне в виде матрицы наборы из 16 слов (8 инфинитивов и 8 существительных единственного числа именительного падежа). Слова состояли из 2–3 слогов и не повторялись в разных заданиях.
В первом задании D (наиболее сложный вариант) испытуемые должны были составить в уме рассказ, используя как можно больше слов из предъявленного списка. Слова в нем принадлежали разным семантическим полям (сложный вариант). Второе задание Е было аналогично первому, однако предъявляемые слова принадлежали одному семантическому полю (более простой вариант). В этих заданиях разрешалось использовать дополнительно свои слова и менять грамматические формы слов. Третье задание R состояло в восстановлении связного текста из слов без изменения порядка их следования, но при изменении словоформ и добавлении служебных слов. Четвертое задание W (запоминание слов) требовало механического запоминания слов в заданном порядке, сначала начинающихся на одну, затем на другие буквы, и соответствующего их последующего воспроизведения. Таким образом, три типа контрольных заданий позволили максимально приблизиться к получению в результате ряда контрастов данных о мозговой организации собственно творческого процесса.
Второй тест состоял из 3 заданий. В каждом задании испытуемому предъявлялись в течение 90 секунд черными буквами на белом фоне в виде матрицы наборы из 12 слов – существительные в именительном падеже. Все слова состояли из 1–4 слогов и не повторялись в разных заданиях. В первом задании D испытуемому нужно было посредством произнесения вслух слов-существительных, ассоциативно не связанных друг с другом, переходить по цепочке от одного слова к другому. Например, цепочкой между словами «стекло, река» могли являться слова «отражение, вода», и таким образом одна из цепочек получалась как «стекло, отражение, вода, река». Во втором задании Е испытуемому нужно было к каждому слову из матрицы, представляющему какую-то категорию, назвать 5 слов, относящихся к данной категории. Например, «одежда» – «брюки, юбка, носки, рубашка, галстук» и т. д. Третье задание R представляло собой чтение вслух слов, представленных на матрице (Данько и др., 2003; Bechtereva et al., 2004).
Данные электроэнцефалографии теоретически позволяли проводить обсуждение полученных результатов с использованием мирового опыта, так как принципиально подобного рода работы проводились с середины 90-х годов (Petsche, 1966, 1997; Molle, 1996, 1999; Разумникова, 2000; Jausovec, Jausove, 2000). Однако различия в методических подходах и, соответственно, в результатах определяли сложности в сопоставлении данных. Что касается так называемых новых технологий, то работы по изучению мозговой организации и механизмов творчества на их основе начались лишь в ближайшие (последние) годы (Бехтерева и соавт., 2000, 2001; Carlsson, 2000; Bechtereva et al., 2004).
Методические аспекты исследований подробно приведены в соответствующих статьях (Бехтерева и др., 2000; Bechtereva et al., 2005). Как и можно было предположить, данные ЭЭГ позволили обнаружить прежде всего общую реакцию активации, наиболее выраженную при выполнении наиболее сложного, творческого задания, что проявилось в соответствующих контрастах D – Е, D – R, D – W и др.). Однако на основе данных локальной синхронизации выявлены также местные перестройки преимущественно в височных областях, причем в контрасте D – E различия локальной синхронизации в передневисочных зонах были достоверны слева.
Что касается пространственной синхронизации, то наибольшее количество достоверных различий обнаруживалось в контрастах D – R и D – W. Достоверное увеличение контраста пространственной синхронизации в D – E обнаруживалось не во всех группах обследованных лиц.
Полученные данные оказались принципиально воспроизводимыми, однако, как правило, последующие исследования приводили и к получению дополнительных материалов (рис. 1, 2). Идеальной воспроизводимости данных на разных группах испытуемых может препятствовать ряд факторов, причем индивидуальный опыт испытуемых играет здесь не последнюю роль.
Почти тривиальным может считаться здесь пожелание о «проведении дополнительных исследований», а также – о проведении сопоставимых и желательно аналогичных исследований в различных лабораториях, на разных группах лиц.
Следует, однако, подчеркнуть, что и в этом случае не исключено, что никакие высокие технологии сегодняшнего и завтрашнего дня не спасут от некоторого разнообразия в результатах в связи с индивидуальными вариациями стратегий и тактик мозга в «свободном полете» творчества, при решении задач, регламентированных (если!) только желанным результатом.
Проведение исследований мозговой организации вербального творчества на основе тех же психологических тестов, но с регистрацией нейродинамики по данным локального мозгового кровотока (ПЭТ) также подтвердило возможность получения принципиально воспроизводимых результатов, причем проведение дальнейших исследований позволило получать все более полные и надежные данные, рисуя все более убедительную картину о главных мозговых зонах, наиболее значимых для вербального творчества.
В 2000 году мы писали о трех зонах активации в мозгу в контрасте D – E, четырех зонах активации в контрасте D – R, шести зонах активации в контрасте D – W и т. д. Исследования, проведенные в последующие годы (Старченко, 2002, 2003; Bechtereva, 2004), позволили обнаружить более полную картину, выявить дополнительно ряд зон активации мозгового кровотока в исследуемых условиях (рис. 3, 4, табл. 1, 2).
Таблица 1. Результаты исследования при использовании первого теста
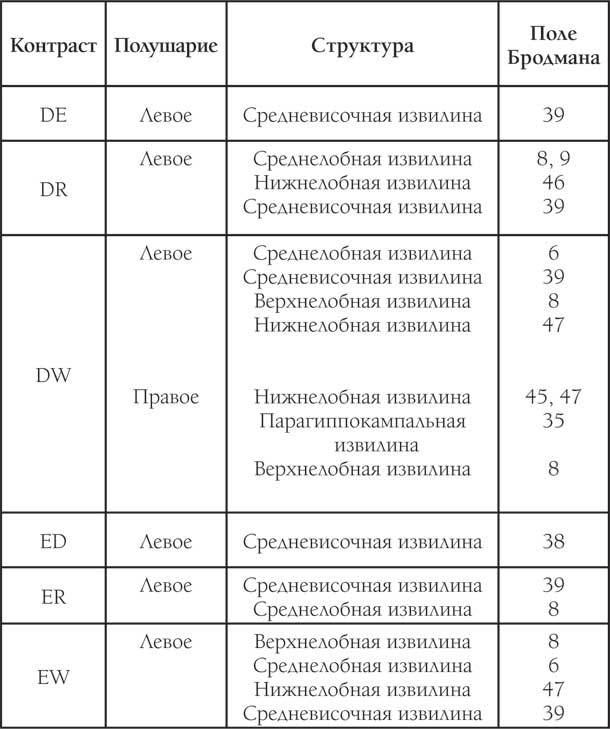
Окончание таблицы 1
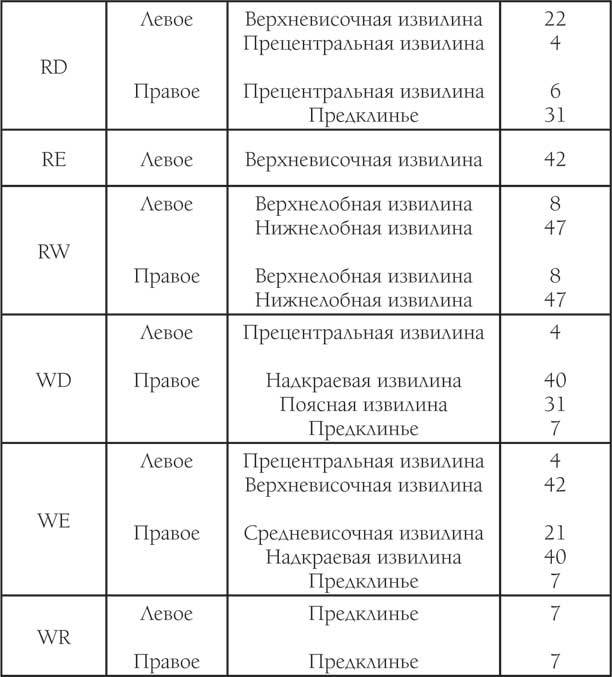
Таблица 2. Результаты исследования при использовании второго теста

Окончание таблицы 2
Вполне понятно, что обнаружение множества зон активации в мозгу, зависимых от типа контраста, является важным для расшифровки структурно-функциональной организации мозга применительно к основным составляющим творческого процесса. Так, предполагается, что в контрасте D – E обнаруживаются мозговые корреляты гибкости творческого процесса и т. д.
Для иллюстрации (наглядности) результатов о «главных» зонах мозга, обеспечивающих вербальный творческий процесс в зависимости от стратегии решения задач, а также от других важнейших составляющих заданий (и прежде всего – фактора сложности), были использованы возможности F-статистики (выявление зон интереса, в которых кровоток меняется в зависимости от условий исследования).
Анализ результатов с помощью F-статистики в условиях применения первого типа задания выявил 8 областей интереса. Графики, иллюстрирующие относительный уровень активации этих областей мозга в различных условиях, могли быть разделены на три паттерна по их виду. В связи с нелинейностью первого паттерна (отсутствием зависимости между уровнем кровотока и последовательности заданий), отвечающего предположению о различиях творческого компонента – в заданиях D, Е, R и W, данный паттерн активации рассматривается как характеризующий собственно творческий процесс.
Второй паттерн активации имеет вид линейной зависимости (уровень активации пропорционально снижался от задания D к W). Предполагалось в связи с характером соотношения заданий по сложности, что данный линейный паттерн отражает, скорее всего, именно степень сложности заданий. Третий паттерн активации, где задания D, Е и R не различаются по уровню активации, отличаясь в то же время от задания W отражает, по-видимому, общность заданий D, E и R в построениях синтаксических (грамматических) конструкций (рис. 5).
Анализ областей интереса по результатам теста второго типа выявил также несколько областей интереса. По той же логике анализа были выделены три группы паттернов, первый из которых предположительно был наиболее тесно связан именно с творческим компонентом заданий, второй – с их сложностью, а третий, скорее всего, отражал слуховые составляющие задач (рис. 6).
Данные исследования областей интереса иллюстрируют связь с творческим компонентом заданий в первом случае средневисочной извилины (ПБ 39). По литературным данным, эта структура связана с оперативной рабочей памятью (Collette et al., 2001; Zurowski et al., 2002) с переключением (Sohn et al., 2000), может являться звеном системы, участвующим в образовании параноидных галлюцинаций. Все это не противоречит обеспечению данной зоной гибкости мышления и также важному для творчества подключению фантазии и воображения. Другие зоны существенной активации (например, ПБ 8) непротиворечиво литературным данным могут быть связаны с активацией соответствующего массива памяти (Павлова, 1988; Petsche, 1996, 1997, и из более близких по времени и методике работ – данными Carlsson, 2000).
Что касается результатов применения F-статистики к данным ЛМК при реализации теста второго типа, они иллюстрируют данные второй таблицы о связи с творческим процессом левой надкраевой извилины (ПБ 40) и поясной извилины (ПБ 32). Сопоставление с данными соответствующей литературы (Booth et al., 2002; Sohn et al., 2000; Knauff et al., 2000, 2002; Finchan et al., 2002) дает основание полагать, что в данном случае именно 40-е поле наиболее тесно связано с творческим процессом и в максимальной степени отвечает за фактор гибкости мышления. ПБ 32, судя по сущности контраста D – R и данным литературы, скорее всего, связано с требующимися для успешного решения творческой задачи процессами селекции (Petersen et al., 1988; Pardo et al., 1991; Taylor et al., 1994). Возможно, эта зона отвечает за «собственную» для творческого процесса эмоциональную составляющую. Вероятность этих предположений дополняется рядом литературных данных о важности теменных регионов для творческих процессов (Petsche, 1966, 1997; Molle, 1999; Jausovec, Jausovec, 2000; Rasumnikova, 2000).
Таким образом, по данным ПЭТ, в известной связи с характером используемой волонтерами стратегии наиболее значимыми зонами для творческого процесса оказались ПБ 39 и 40, причем во втором случае существенно важна и область ПБ-32 (поясная извилина). Как указывалось, в исследованиях наблюдалась существенная активация и в других зонах (ПБ 8, 45, 46, 47) (Bechtereva et al., 2005).
Сейчас появляются исследования мозговой организации творчества, сопоставимые по психологическим и физиологическим подходам (Howard-Jones P. A. et al., 2005). В зависимости от контраста (творческое – нетворческое задание и т. д.) обнаружены активации в различных зонах мозга (ВА 10, 9, 9/32, 17, 18, 40, 32, 24, 18, 9/10). Наиболее значимыми для творческого процесса авторы считают премоторную зону правой лобной области. Сходные данные приводились и в некоторых наших предыдущих работах (Behtereva N. P., 2004). В работе авторов среди обнаруженных достоверных перестроек в некоторых условиях эксперимента проявляются также и ВА40 и ВА32.
Таким образом, «в первом приближении» получены данные о перспективности изучения мозговой организации творчества с помощью количественной ЭЭГ, мозгового кровотока (ПЭТ) и fMRI. И в том и в другом случае удалось обнаружить преобладание значимых перестроек нейродинамики в ряде областей мозга. Дополнительное применение в ПЭТ-исследованиях приема F-статистики (определение зон интереса, зон активации, которые связаны с определенными условиями задания) позволило наглядно представить зоны, наиболее тесно связанные именно с творческой деятельностью, причем несколько различающиеся топографически в зависимости от стратегий выполнения заданий, использованных волонтерами. Оно показало зависимость мозговой нейродинамики от сложности заданий и некоторых других факторов.
Межлабораторный проект с обязательной унификацией по крайней мере максимального количества аспектов исследования – несомненно, идеал в такой сложной задаче, как получение надежных данных об основных мозговых коррелятах собственно творческого процесса в картах его мозговой организации. Удастся ли приближение к этому идеалу, по другому поводу реализованного в 50-х годах французским ученым Гасто (Gastaut, 1954) в нашем объединенном Интернетом и все же таком разобщенном мире, – вопрос прежде всего лидерства и, конечно, финансирования в науке.
Известна дискуссия о кризисной ситуации в оценке результатов картирования высших функций мозга в целом (Fox et al., 1998; Cabeza Nyberg, 2000; Grezes, Duety, 2001; Chein et al., 2002; Phan et al., 2002; Jobard et al., 2003 и др.). Несмотря на это, данная конкретная линия исследований мозговой организации творчества как будто бы «держится на плаву», прежде всего, по-видимому, за счет рациональной конструкции тестов и адекватных приемов анализа. Физиологические результаты, как видно из данных статистики, достаточно надежны.
И в то же время, как видно именно из приведенных выше данных 2000–2005 годов, топически результаты ЭЭГ и ЛМК, и прежде всего по мозговым зонам максимальной значимости для вербального творческого процесса, не совпадают. Накопление материала увеличило этот разрыв. Такого рода разночтения, по-видимому, не только наша локальная проблема. Отсюда на основе первых наших работ, и именно тех, которые были проведены на сходной или одной и той же группе волонтеров (2000–2001), была предпринята попытка сопоставить данные полиметодических исследований о мозговой организации вербального творческого процесса по зонам максимальной значимости мозговых перестроек и в зависимости от конкретных контрастов.
Таблица 3. Соответствие проекций стандартных ЭЭГ-отведений системы 10–20 и полей Бродмана
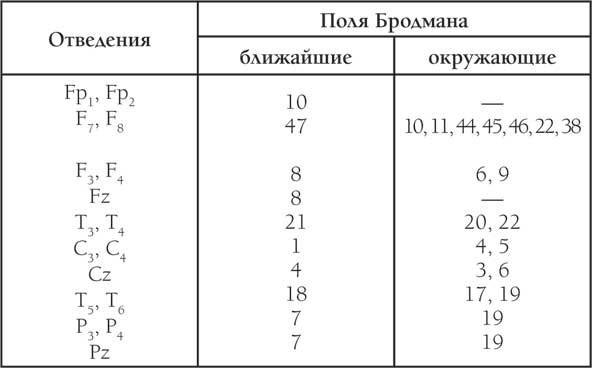
Для топографического сопоставления результатов мы предприняли попытку использовать единый анатомический (топографический) язык полей Бродмана. ЭЭГ-данные рассматривались через таблицу соответствия стандартных ЭЭГ-отведений (системы 10–20) с полями Бродмана, этот же язык (поля Бродмана) использовался нами в описании топографии результатов ЛМК(табл. 3).
Зоны наибольших изменений ЛМК и локальной синхронизации могли анатомически (топографически) совпадать и могли, наоборот, так же существенно анатомически различаться. Приведем некоторые конкретные данные такого сопоставления.
В исследованиях, взятых в основу сопоставления, в наиболее значимом контрасте D – Е (оба задания творческие) локальные изменения на ПЭТ четко прослеживались в правой лобной доле (ПБ 10, 11, 44, 45, 46, 47). Изменения ЭЭГ в данной области в тех же условиях выявлены не были. Можно медитировать на тему о том, что в этом случае изменения импульсной активности (ЛМК) проявлялись без местной и без дистантной синхронизации. В контрасте D – R обнаружена активация ЛМК в левой лобной доле (ПБ 6/8, 8, 46) и теменно-затылочной области (ПБ 37, 7, 19) слева. На ЭЭГ изменения ПС обнаруживаются в обеих лобных долях и также превалировали слева (ПБ 45, 8, 9, 10), топографически, не идеально, но все же как бы подтверждая данные ЛМК, дополняя их раскрытием физиологической сущности изменений, – большие области мозга объединялись видимым образом в творческой работе для реализации задачи D. Как указывалось выше, разные контроли, разные контрасты позволяют выявить мозговую организацию различных сторон творческого процесса.
Несомненно, проведение ПЭТ- и ЭЭГ-исследований у сопоставимых (сходных) групп испытуемых при использовании одних и тех же психологических тестов позволило получить большой массив новой информации. Однако именно она при всей актуальности идеи призыва к полиметодичности убедительно показывает, что даже в тех случаях, когда значимые изменения мозговой нейродинамики, получаемые с помощью различных методик, топографически сходны, они, описывая разные стороны жизнедеятельности каждой зоны мозга, всегда могли и должны были расцениваться лишь как взаимодополняющие данные, представляя все более полную картину развивающихся в мозгу событий, не отрицая, а лишь дополняя данные, полученные с помощью различных подходов.
В ходе дальнейшего изучения вопроса были получены убедительные примеры взаимных дополнений, расшифровки локальных событий в мозгу. Так, в контрасте «творческое, эмоционально-нейтральное задание – контроль» на рис. 7 выявлена локальная десинхронизация в ЭЭГ в области левой лобной доли, и одновременно в той же области зарегистрирована активация ЛМК (Шемякина, Данько, Медведев, 2005). Отсюда можно себе представить, что «активация» ЛМК здесь действительно представляет активацию нейронной активности без ее структурирования. В этом частном случае данные пространственной синхронизации подтверждают эти соображения. Можно привести и другие примеры соотношений данных ЭЭГ и ЛМК, однако следует помнить, что это всегда будет один из возможных вариантов их взаимодополняющих свойств.
Допустимо полагать, что использованный нами топографический подход может рассматриваться как своего рода предыстория новой методологии, где комплексное многоуровневое описание физиологических процессов в каждой зоне мозга явится основой новых, далее действительно содержательных карт нейродинамических событий в мозгу.
Используя высочайшие современные технологии исследования мозга, мы все, как хорошо известно, пережили эйфорию картирования самых различных мозговых функций. Выпущено бесчисленное множество статей в профильных и не очень профильных изданиях, созданы обобщающие труды (Gusnard and Raichle, 2001). Исследование мозговой организации различных видов психической деятельности и состояний привело, однако, с накоплением материала к тому, что при успехах в почти каждой из отдельных работ, как указывалось выше, создалось впечатление о том, что физиологические корреляты самых разных видов психической активности могут быть обнаружены почти в каждой точке мозга. С другой стороны, как видно из приведенных выше данных наших наблюдений и новейших данных литературы, сложнейшие процессы высшей нервной деятельности «задействуют» большое количество областей мозга.
При всем накопленном «многознании» о мозге человека «кризисный аспект» сегодняшней ситуации невольно напоминает давние споры, ставшие анахронизмом уже в середине XX столетия, об эквипотенциальности мозга (Lashley) и локализационизме, представлений о мозге как лоскутном одеяле, сотканном из самых различных центров, в том числе и «очень» высших функций. Как известно, восторжествовал «третий», системный подход, в который «улеглись» и прошлые споры и укладываются новые данные. Сегодняшнее положение в картировании высших функций, кстати, дополнительно подтверждает представления об обеспечении этих функций мозговой структурно-функциональной организацией с жесткими и гибкими звеньями (Бехтерева Н. П., 1966).
Можно понять результаты картирования или, точнее, подлежащих им событий в мозгу с позиций огромного востребования именно гибких звеньев системы обеспечения психических процессов в оптимальных условиях жизни и психологического эксперимента, однако при поиске жестких (основных) звеньев системы заставляет обращаться к дополнительным приемам анализа (см. выше) и, может быть, не сразу, обязательно к данным более тривиальных методик (имеются в виду, в частности, данные электростимуляции мозга, обобщенные Валленштейном (Vallenstein, 1973) и у нас в лаборатории В. М. Смирновым (1976). Сегодня надо отдать должное методам анализа, используемым в науке о мозге, – из сложностей топографического сходства карт мозга при самых различных процессах высшей нервной деятельности и нередком множестве достоверно активированных зон в каждом данном исследовании нужные решения находятся с помощью дополнительных приемов анализа данных. Было бы очень заманчиво найти пути увеличения надежности методов, подобных уже упоминавшейся F-статистике. Подчеркнем, однако, уже сегодняшний анализ позволяет без обращения к данным инвазивных методов находить «жесткие» звенья систем, в большинстве случаев зоны мозга, наиглавнейшие для какой-то данной деятельности, и обеспечивать контроль валидности данных. На этом можно было бы и закончить обсуждение данной позиции, хотя здесь возможны и дальнейшие прорывы. Но у нас уже и сейчас есть в активе «кое-что», что определяет наш оптимизм в будущем изучаемой проблемы. Это – воспроизводимость результатов, о которой мы упомянем и далее в тексте при описании развития работ. Для меня как физиолога воспроизводимость результатов – наиболее надежный критерий их валидности.
Несмотря на проблемы в продвижении в познании организации и механизмов мозга за счет полиметодических исследований, именно эта позиция, по-видимому, все же наиболее перспективна. Некоторые трудности, неизбежно ждущие исследователей при реализации этого рода работ, иллюстрированы нами выше.
Работая всю долгую жизнь в коварной проблеме «Мозг человека», видишь не только угнетающий, но и мобилизующий эффект казалось бы иногда тупиковых ситуаций, преодоление которых подтверждает известный тезис развития знаний по спирали. В XX столетии произошли два уже упоминавшихся выше основных методологических прорыва в изучении мозга человека (Бехтерева, 1997; первый, в котором исследователь обрел полноту возможных тогда знаний о точке мозга, – инвазивные исследования, и второй – технологическим прогрессом обусловленный – получения монометодических знаний о всем мозге). Полагаем, что получение с помощью современных методик полноценных знаний о точках мозга и объединение данных в целую картину явится следующим уровнем этой спирали и поможет еще дальше продвинуться по бесконечной дороге познания мозга человека. Перспективность этого пути несомненна. Он на сегодня иллюстрируется пилотной работой С. В. Медведева и А. М. Иваницкого (2003) и других.
То, о чем говорилось выше, относилось прежде всего к усовершенствованию и физиологической расшифровке данных картирования мозга. И в то же время, веря в значение совершенствования этого, первого, базисного этапа в изучении мозговой организации высших функций мозга, и в частности творчества, уже сейчас важно и можно исследовать мозговые механизмы реализации творчества, его облегчающие и отягчающие факторы. Понятно, что даже будущая полноценная карта мозга приобретает особый смысл при раскрытии управляющих механизмов мозга.
С целью подхода к этой проблеме использовалось изучение влияния индуцирования эмоционального фактора и детекции ошибок на нейродинамику мозга при вербальном творчестве, отражающуюся в картах, приведенных в ходе изучения собственно мозговой организации данного процесса или специально дополнительно сконструированных по тому же основному принципу. В обоих случаях, также дополнительно к приведенным выше, использовались и еще некоторые другие психологические «субтесты» (Шемякина, Данько, 2004).
Так, например, в основном задании испытуемые должны были предлагать как можно больше оригинальных определений эмоционально положительных, отрицательных и нейтральных понятий через понятия из других семантических полей. В контрольном задании применялась выработка определений к эмоционально нейтральным понятиям через слово из того же семантического поля. Субъективное изменение креативности при введении эмоционального фактора характеризовалось некоторым увеличением беглости, однако при уменьшении оригинальности ответов.
Анализ показал более выраженный эффект эмоциональной индукции на процессы в коре головного мозга по сравнению с эффектом, вызванным заданием творческого процесса, а также влияние характера эмоций на изменения мозговой активности. Без индукции эмоций наибольшие изменения локальной синхронизации, как приведено выше, при выполнении творческих заданий наблюдались в левой лобной зоне (уменьшение локальной синхронизации). Пространственная синхронизация при этом уменьшалась главным образом в межполушарных парах с фокусами сходимости в переднелобных, передне-, средневисочных зонах левого полушария и средне-, задневисочных зонах правого полушария (а также в зонах отведений – F4, 02). Индукция положительных эмоций вызывала существенное увеличение локальной и пространственной синхронизации с участием большинства зон коры. Индукция отрицательных эмоций была связана с топографически более диффузными и разнонаправленными по знаку (и увеличение, и уменьшение) изменениями параметров ЭЭГ. Положительные эмоции в данном эксперименте вызывали больший эффект, чем отрицательные. На основе данных количественной ЭЭГ показано, что эффекты «творчества» и вызванных положительных эмоций могли быть противоположными по знаку. Особенно отчетливо это проявлялось в бета-2 (18,5–30 Гц) диапазоне ЭЭГ (см. рис. 7).
Исследование с индукцией эмоций как одного из возможных управляющих механизмов было предпринято нами на основе хорошо известного положения, что творчество, как правило, происходит при изменении эмоционального фона индивидуума. Однако на основе физиологических данных создалось впечатление, что вызванные в нашем эксперименте эмоции были чужеродными творческому процессу. Из литературы и жизненной практики известно, что соотношения эмоционального и творческого факторов очень неоднозначны, что в известной мере сопоставимо с тем, что мы видим в мозговой реорганизации при данной пилотной модельной ситуации с попыткой оценки влияния эмоционального фактора на вербальное творчество. При творческом процессе развивается свое, особое эмоциональное состояние, в данный момент и именно этому процессу присущее. Мы полагаем, что изучение соотношений эмоций и творческого процесса должно быть продолжено и, в частности, с помощью дополнительного конструирования психологических методик (тестов).
Одним из открытых нами ранее управляющих механизмов мозга является детектор ошибок. Огромное количество статей, опубликованных в последние годы по поводу детекции ошибок, полностью подтвердило наличие в мозгу такого механизма. Показана (Bechtereva et al., 2005) его обязательная «задействованность» в определенных условиях, в связи с чем именно при исследовании проблемы творчества мы позволили себе использовать специальный для данного случая методологический прием. Сущность этого приема состояла в проведении исследований в условиях, в которых детектор ошибок обязательно должен был активироваться, что, позволяя обходиться без одновременной регистрации активности самого детектора ошибок, расширяло возможности собственно психофизиологического изучения творчества.
Как известно и как неоднократно подчеркивалось здесь, одним из обязательных определений творчества считается отход от стереотипа, предложение новых оригинальных, продуктивных решений. В то же время по ходу всей формирующейся (формируемой) индивидуальной жизни развитие индивидуума всегда представляет собой процесс развития и определения границ дозволенного. Творчество, как хорошо известно, всегда развивается в рамках какой-то деятельности и одновременно происходит с выходом за ее рамки.
Проблемные ситуации, неизбежные в изучении мозговой организации творчества, не только породили накопление ценных фактических данных, но и вызвали размышления и осмысление возможных идейно-методических перспектив проблемы.
В изучении вопроса о роли детектора ошибок в процессе творчества мы ориентируемся на хорошо известную многоплановость, поливалентность мозговых механизмов. Отсюда известная ограничительная роль детектора ошибок может проявиться и негативом, и позитивом – речь идет не только о «запретах», но и о возможности поддержания с помощью детектора именно нетривиальности и продуктивности творческого процесса. В исследовании и первого и второго аспектов исключительно важна конструкция психологических тестов.
Прямых данных о возможных отношениях детектора ошибок и творчества сейчас практически нет. Однако кое-что об отношениях детектора ошибок и мыслительных процессов можно извлечь из экспериментальных данных.
Мировой экспериментальный материал по изучению детектора ошибок в мозгу в психологическом плане, как правило, базируется на реализации испытуемыми не только простых, но и более или менее сложных задач высшей нервной деятельности. В том же плане могут быть рассмотрены и наши наблюдения при прямой регистрации реакций детекции ошибок в области передних отделов поясной извилины в тесте с прослушиванием правильных (корректных) и ошибочно построенных (некорректных) фраз (Bechtereva et al., 2005, рис. 8). И наконец, уже более прямо свидетельствует о возможной роли детекции ошибок собственно в творческом процессе топографическое единство мозговых зон детекции ошибок и «главных» творческих зон, в частности ПБ 40, 8. В литературе имеются указания на то, что ПБ 40 в процессе принятия решений более активна при низкой частоте появления ошибок (Paulus et al., 2002). Так как в творчестве теоретически не может быть ошибок и предварительного ответа на творческую задачу не существует, этот анатомо-функциональный факт дает дополнительные основания полагать, что связь может осуществляться не только по одной из возможных схем отношений творчества и детекции ошибок, но проявляться и «негативом», и «позитивом».
Специально для попыток проникновения в эти соотношения были разработаны и использованы дополнительные психологические тесты. Испытуемым-волонтерам предъявлялись две пары заданий. Предъявлялись хорошо известные пословицы и поговорки, в которых последнее слово отсутствовало. Волонтеры должны были предложить свой вариант окончания, который бы полностью изменил суть пословицы или поговорки (задание С – creative). В контрольном задании предлагалось просто вспомнить недостающее слово-окончание (R – remember). Вторая пара заданий отличалась от первой тем, что в текстах заданий С и R присутствовали ошибки в форме перестановки букв или их замен без изменения длины слов. Предполагалось, что такого рода задание будет активировать детектор ошибок (соответственно задания CD и RD). Испытуемым предлагалось: 1) не только выполнить основную задачу, но и определить количество ошибок в текстах («осознанный вариант»); 2) не обращать внимания на ошибки и решать творческую задачу («неосознанный вариант»). Исследовалась локальная синхронизация в различных ЭЭГ-диапазонах. Существенная разница была обнаружена не только в контрасте C – R, но и в контрасте CD – C также. Результат проявлялся главным образом в полосе гамма-частоты. Добавление детекции ошибок (в «осознанном варианте») к основной задаче уменьшало мощность г-ритма (CD – C) и таким образом уменьшало различия в контрасте CD – R (рис. 9). Похожий, но меньший эффект наблюдался в полосе в2-ритма (методика и результаты этой части работы подробнее представлены в статьях Bechtereva et al., 2005). Введение в тест ошибок в тексте без активации внимания к нему («неосознанный вариант») вызвало существенно меньшие изменения ЭЭГ, по тенденции схожие с «осознанным вариантом». В связи с конструкцией теста сделана логичная попытка оценки влияния на ЭЭГ активации внимания. Это осуществлялось в модифицированных условиях принципиально того же эксперимента. В предъявляемых текстах не было ошибок, а были звездочки, отношение к которым регламентировалось такой же, как указано выше, инструкцией. Активация внимания в нашем варианте теста вызывала относительно небольшие изменения ЭЭГ, из которых наиболее отчетливыми были угнетения медленноволновой (D) активности. Все варианты включений (дополнений) в основной текст оказывали большее влияние на мозговые корреляты основного задания и, соответственно, меньшее – на контрольное. Проведение исследований в данном варианте теста подтвердило связь динамики наиболее частных компонентов ЭЭГ с креативностью, хотя изменения в g-диапазоне не всегда достигали уровня достоверности. Воспроизводимость указанных результатов, однако, была очевидной (рис. 10).
Гамма-активность, считающаяся характерной для мыслительных процессов (Fitzgibbon S. P. et al., 2004; Данько и соавт., 2005; Thilo Womelsdorf, 2006), оказалась в условиях данного теста также тонким критерием подавления потенциала мозга в условиях активации детектора ошибок.
Исследование мозговых механизмов творчества, по существу, только начинается. В то же время и потому, что становится все очевиднее, что детектор ошибок – важнейший управляющий механизм мозга на разных уровнях сознания, в разных процессах высшей нервной деятельности, мы представляем схему о возможной роли детектора ошибок в здоровом и больном мозге, где только часть схемы-гипотезы может считаться условно подтвержденной, а ее основная часть сейчас дается нами как гипотеза, руководство к действию, как идея, и, в частности, в изучении механизмов творчества.
Как в случае обсуждавшейся нами ранее специально (Bechtereva, 1984) поливалентности других механизмов мозга, и в том числе их различных проявлений в здоровье и болезни, мы допускаем возможность такой же схемы в роли детекции ошибок в зависимости от здоровья, болезни и творчества человека.
Итак, положение первое. Детектор ошибок в здоровом, скажем так, обычном среднестатистическом мозге – это структура системы, сформировавшаяся главным образом в ходе индивидуального развития. Она существенно оберегает человека от раздумий в стереотипных, тривиальных ситуациях в ходе обычной жизни, контролирует оптимальную реализацию процессов высшей нервной деятельности. При любом обучении наряду с позитивом развития в мозге неизбежно формируются и необходимые, и излишние ограничения (вплоть до табу). Они работают с помощью детекции ошибок, через формирование детекторов ошибок. В формировании табу в более масштабных проблемах ярким примером такого рода процесса являлось многовековое церковное воспитание (не убий, не укради), что осложняло посягательство на границы законов.
Положение второе. Детектор ошибок разрушается под влиянием самых различных причин, или его активность может становиться чрезмерной. В обоих случаях развиваются нарушения психической деятельности. В случае чрезмерной активности детектор ошибок может превратиться в детерминатор ошибок. По-видимому, во втором случае именно превращением детектора ошибок в детерминатор ошибок могут определяться некоторые формы психической зависимости с проявлениями типа устойчивых наркоманий (Медведев С. В., Аничков А. Д., Поляков Ю. И., 2003) и некоторых форм маниакального поведения. Детектор ошибок в этом случае из полезного слуги превращается в злого хозяина.

Мы вместе много десятков лет. Раиса Вольская и Татьяна Аверьянова
И наконец, положение третье, имеющее прямое отношение к задачам изучения физиологии творчества. Рождается человек с большой (огромной) творческой потенцией, не теряющейся по пути детских инфекций и прочих жизненных сложностей. Естественно, что сила ограничений воспитанием, обучением различна у разных индивидуумов, а иногда такого рода «нажим» может вызывать и обратную реакцию, хотя в целом, статистически, в государстве это, как известно, работало.
Детектор ошибок должен и может теоретически препятствовать выходу в новизну, прорыву через известные догмы и законы, преодолению сформированных ограничений, полученных в том числе и в обучении. Однако ограничения именно в мозгу любознательного и дерзкого творца либо исходно преодолеваются, либо формируются сознательно или бессознательно по принципу «не мешать», либо при осознании стремления к прорыву в неизвестное переформировываются, «перевоспитываются» из ограничителей – в помощников. Возможность функциональных перестроек детектора ошибок прямо показана нами при регистрации детекции ошибок в передних отделах поясной извилины в ходе диагностических и лечебных электрических стимуляций (см. рис. 8) (Bechtereva, 2005). В мыслительном (и любом другом) творчестве детекция ошибок начинает оберегать от тривиальности (от «изобретения велосипеда») и от бессознательного плагиата.
Реализующийся творец начинает определять более или менее частную или глобальную историю мира.
Многолетний опыт нашей работы в области физиологии здорового и больного мозга человека неизбежно заставлял нас обращаться к многоликости управляющих механизмов мозга. Нами делается попытка рассмотрения этого вопроса, в том числе в физиологической природе творчества, его особого положения в мыслительных процессах, его магии. Магия творчества в обществе – прорыв к новой истине сквозь устоявшиеся знания. Физиологически – это процесс, развивающийся при преодолении и переориентации детектора ошибок (из противника в помощника), создание и воссоздание «своего» эмоционального фона. У творчества свое, только ему присущее состояние, «свои» эмоции, «своя» детекция ошибок. В свободном полете творческой мысли индивидууму-творцу надежно служат различные, в том числе и преобразованные, механизмы мозга. Творчество, преобразуя мир, своим базисом имеет творчески преобразованный мозг человека.
Будущее исследователей творчества – в руках психолога, который поможет с помощью рационального эксперимента расшифровать то, что сейчас может обозначаться лишь как физиологические механизмы магии творчества. Естественно, с помощью физиологов.

