Глава одиннадцатая
Скрещивание. Доминантные и рецессивные признаки
– Что получится, если скрестить свинью со змеей?
– Свинья, которая несколько раз в год станет сбрасывать сало.
Заслуга Грегора Менделя состоит в том, что он первым сорвал покров тайны с передачи признаков по наследству.
Беда Грегора Менделя состоит в том, что на его революционные и эпохальные (никакого пафоса, так оно и есть) открытия научное сообщество не обратило внимание. Никому не известный (в широком смысле) монах занимается на досуге опытами по скрещиванию садового гороха и публикует об этом статьи – что с того? Ничего особенного!
Сам Мендель понимал, что «что-то особенное» в его работах есть, и старался, чтобы итоги его семилетнего труда были замечены. Не удовлетворившись публикацией в сборнике местного Общества естествоиспытателей (дело было в Брюнне, который ныне называется Брно), Мендель напечатал свои «Опыты над растительными гибридами» в виде отдельной брошюры, которую разослал именитым ученым-ботаникам. Ни один «корифей» не откликнулся. Увы, так бывает, когда кто-то высказывает идеи, до которых общество еще не «доросло». Хотя, предпосылки к тому, чтобы дорасти, уже имелись. Мендель опубликовал результаты своих экспериментов в 1865 году, спустя шесть лет после выхода дарвиновского труда «Происхождение видов путем естественного отбора, или Сохранение благоприятствуемых пород в борьбе за жизнь». Предпосылки для правильного восприятия и правильной оценки того, что сделал Мендель, в 1865 году имелись. Но не сложилось, бывает. Слава пришла к Менделю посмертно в начале XX века, когда вдруг выяснилось, что он основал новую науку – генетику.
Если вникнуть в суть работ Менделя, то окажется, что ничего невероятного, в смысле – гениального, в них нет. Довольно простые эксперименты по скрещиванию, наглядные результаты, которые просто невозможно было истолковать как-то иначе… Но гениальность менделевских экспериментов заключалась именно в простоте, а также в методе.
Мендель исследовал только простые, наглядные, то есть хорошо определяемые признаки, такие как цвет семян или их форма. Это раз.
Мендель выбирал только альтернативные признаки, которые имели только два четко различающихся варианта и не имели промежуточных вариантов (семена гороха могли быть либо гладкими, либо морщинистыми, но не гладко-морщинистыми или же морщинистыми в различной степени). Это два.
Мендель проводил моногибридные (одногибридные) скрещивания, то есть скрещивал растения, отличающиеся друг от друга только по одному признаку. Это три.
Мендель наблюдал передачу признака на протяжении НЕСКОЛЬКИХ поколений. Это четыре.
Мендель фиксировал точное количество особей с определенным признаком и тщательно анализировал полученные данные. Это пять.
Просто, как закон всемирного тяготения, не правда ли?
Просто. И Платон, и Сократ, и Пифагор, и Аристотель, и Демокрит регулярно мылись в ваннах да бассейнах, а закон выталкивающей силы открыл только Архимед.
Закон единообразия гибридов первого поколения, или первый закон Менделя (в отечественном изложении) гласит, что при скрещивании двух гомозиготных организмов, относящихся к разным чистым линиям и отличающихся друг от друга по одной паре альтернативных проявлений признака, все первое поколение гибридов окажется единообразным и будет нести проявление признака одного из родителей.
Давайте вспомним, что гомозиготным называется организм (клетка), имеющий идентичные гены (аллели) в гомологичных хромосомах. Аллелями называются различные формы одного и того же гена, расположенные в одинаковых участках гомологичных хромосом. Организм, имеющий в гомологичных хромосомах различные аллели, называется гетерозиготным.
Гомозиготность группы особей по определенному признаку называется чистой линией, то есть линией, не содержащей «примесей» – других аллелей.
Аллели принято обозначать одной или несколькими латинскими буквами. Мы (как и Мендель) будем рассматривать гены, имеющие только два аллеля, поэтому для их обозначения будет достаточно одной буквы.
Мендель заметил, что при скрещивании чистых линий гороха с пурпурными цветками и гороха с белыми цветками первое поколение потомства имело только пурпурные цветки.
При скрещивании таких же чистых линий гороха с желтыми и зелеными семенами, у всех потомков в первом поколении семена были желтыми.
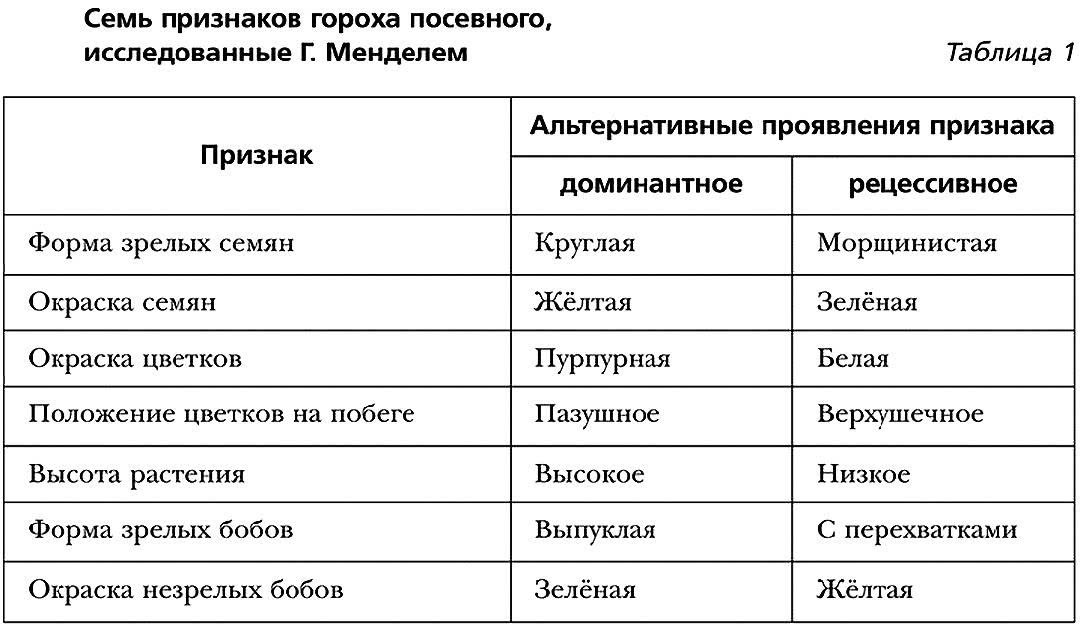
Семь признаков гороха посевного, ислледованного Г. Менделем
При скрещивании чистых линий гороха с гладкими и морщинистыми семенами у всех потомков в первом поколении были гладкие семена.
Гибриды первого поколения всегда единообразны по данному признаку и приобретают признак одного из родителей.
Более сильный признак, наследуемый первым поколением, Мендель назвал доминантным, а слабый, подавляемый, – рецессивным. Мы будем обозначать доминантные признаки заглавной буквой, а рецессивные – строчной.
Итак, гибриды первого поколения всегда приобретают доминантный признак.
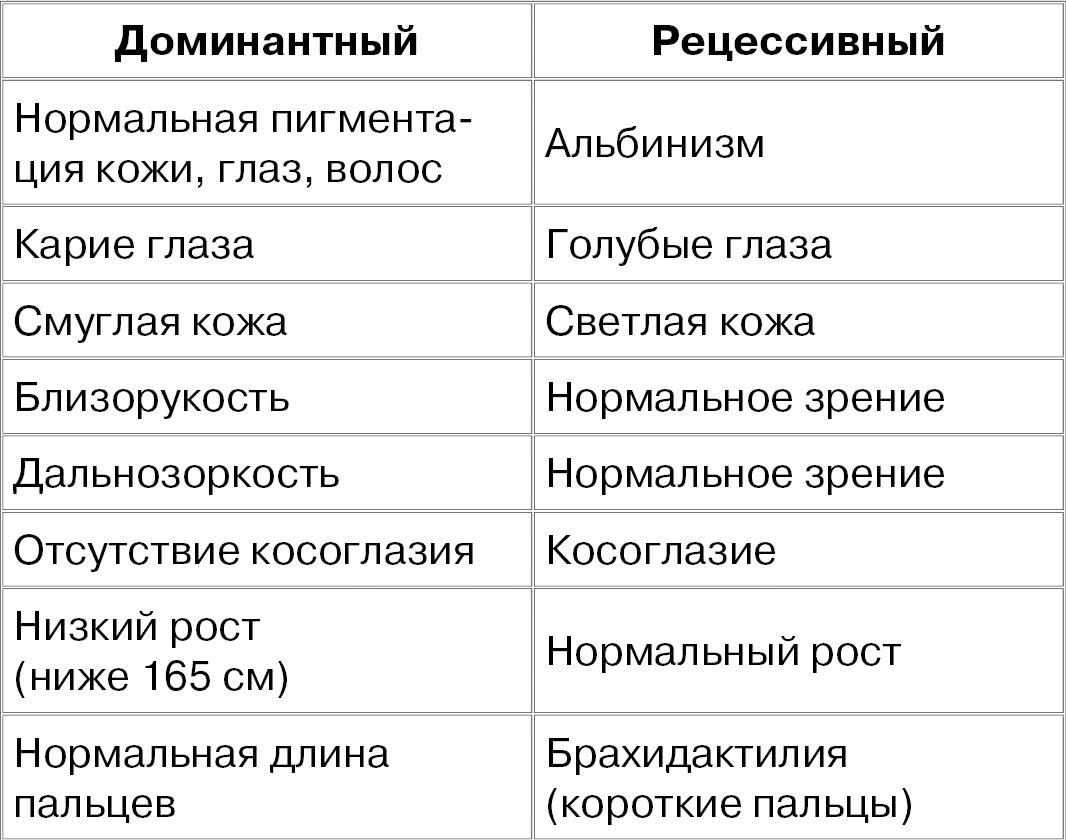
НЕКОТОРЫЕ ДОМИНАНТНЫЕ И РЕЦЕССИВНЫЕ ПРИЗНАКИ ЧЕЛОВЕКА
На первом потомстве Мендель не остановился. Он продолжал свои скрещивания по каждому признаку дальше и во втором поколении получил растения с признаками обоих родителей – и с пурпурными, и с белыми цветками. Три четверти от общего числа растений имели цветки с доминирующим признаком – пурпурной окраской, а одна четвертая часть растений имела цветки с рецессивным признаком – белой окраской.
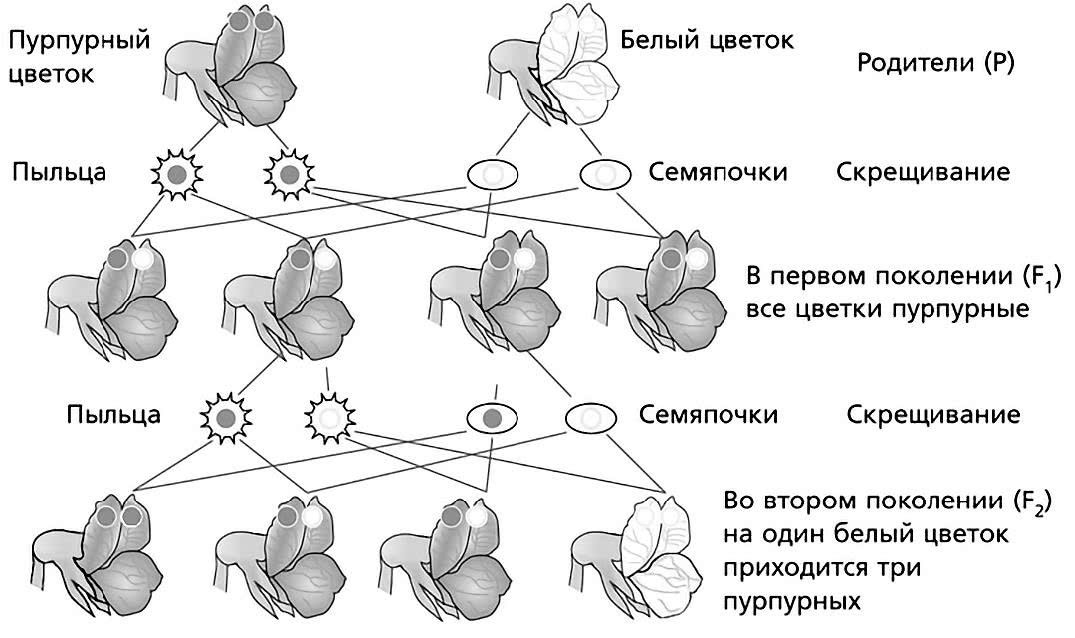
Схема опыления и результатов скрещивания гороха с пурпурными цветками и белыми цветками (поколения принято обозначать латинской буквой F и соответствующим номером).
История повторилась и с другими изучаемыми признаками. Во всех вторых поколениях обнаруживался рецессивный признак. И всюду – на одну четверть. Исходя из этого, Мендель сделал вывод о том, что рецессивный признак у гибридов первого поколения не исчезал совсем, а всего лишь был подавлен доминантным признаком.
Но каким образом рецессивный признак проявился во втором поколении?
Если мы обозначим ген (аллель), отвечающий за доминантный признак, заглавной буквой «А», а ген (аллель), отвечающий за рецессивный признак, строчной буквой «а», то схема первого скрещивания будет выглядеть следующим образом:
АА х аа = Аа + Аа
Гаметы, несущие ген А, соединяются с гаметами, несущими ген «а», и в результате образуются потомки с набором Аа, состоящим из сочетания доминантного и рецессивного гена. Доминантный ген подавляет рецессивный. Потомков с набором аа в первом поколении нет, таким образом рецессивный ген в первом поколении проявиться не может.
Но во втором поколении при скрещивании особей с генотипами Аа мы получим три вероятных сочетания: АА, Аа и аа. Схематически это можно выразить следующим образом:
Аа х Аа = АА + Аа + аа
Если мы уравняем эту схему-уравнение таким образом, чтобы слева и справа было бы одинаковое количество букв «А» и «а», то получим следующее:
2Аа х 2Аа = АА + 2Аа + аа
Таким образом, в трех четвертях потомства второго поколения проявится доминантный ген (одна часть особей с набором АА и две части с набором Аа), а в одной четверти – рецессивный (одна часть особей с набором аа).
Закономерность, характеризующуюся проявлением признаков обоих родителей во втором поколении гибридов, Мендель назвал расщеплением. Единый доминантный признак расщепляется на доминантный и рецессивный.
Закон расщепления, или второй закон Менделя (он же первый в зарубежной литературе) в современной трактовке гласит, что при скрещивании двух гетерозиготных потомков первого поколения между собой во втором поколении наблюдается расщепление в определенном числовом отношении: по фенотипу 3:1, по генотипу 1:2:1.
По фенотипу в трех четвертях потомства второго поколения проявится доминантный ген, а в одной четверти – рецессивный, поэтому соотношение будет 3:1.
По генотипу мы имеем три разных набора генов: АА, 2Аа и аа. Поэтому соотношение будет 1:2:1.
Проведя серию моногибридных скрещиваний, Мендель решил выяснять, каким образом проявляются при скрещивании парные альтернативные признаки гена. Для этого он провел серию опытов по дигибридному скрещиванию – скрещиванию организмов, различающихся по двум парам альтернативных признаков, например, по окраске цветков и форме семян.
Для этого эксперимента Мендель взял растения гороха с гладкими желтыми семенами и с зелеными морщинистыми. У первой разновидности гороха оба признака по генотипу являлись доминантными (AB), а у второй – рецессивными (ab).
Согласно правилу доминирования в первом поколении все семена оказались гладкими и желтыми. Во втором поколении произошло расщепление признаков, причем наряду с родительскими гладкими желтыми и морщинистыми зелеными, появились и совершенно новые семена – морщинистые желтые и гладкие зеленые.
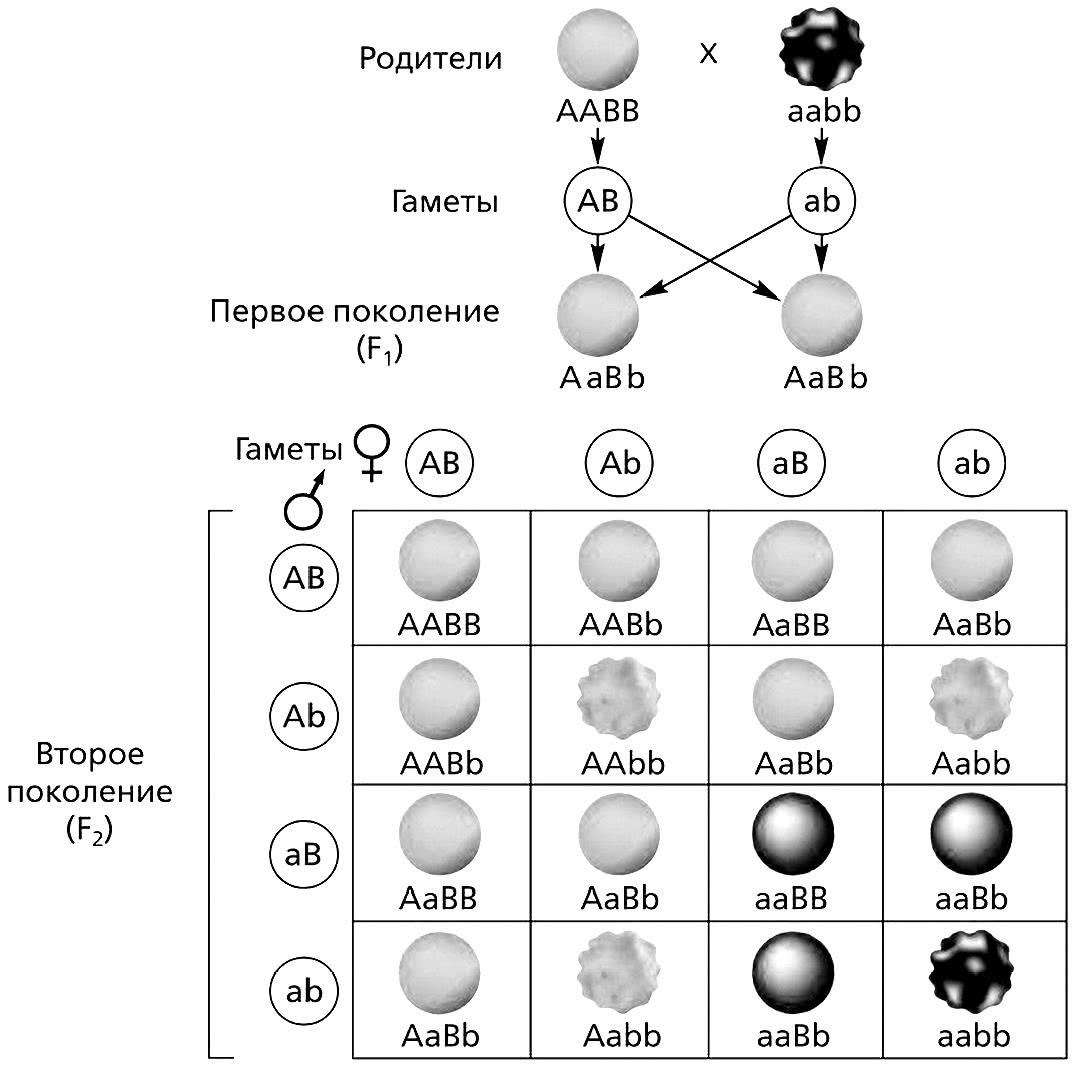
Определение расщепления признаков по генотипу во втором поколении дигибридного скрещивания (А – морщинистые семена, B – желтые семена, b – зеленые семена). Расщепления по фенотипу в соотношении 9:3:3:1 (гладкие желтые: морщинистые желтые: гладкие зеленые: морщинистые зеленые).
Таким образом Мендель установил, что гетерозиготы по двум парам аллельных генов образуют четыре типа гамет в равных количествах – AB, Ab, aB, ab. В двух случаях из четырех гены находятся в том же сочетании, что и у исходных родителей (AB и ab), а в двух других – в новых сочетаниях (Ab и aB, морщинистые желтые и гладкие зеленые). Иначе говоря, расщепление одной пары признаков (желтая и зеленая окраска семян) совершенно не связано с расщеплением другой пары (гладкая и морщинистая форма семян). Расщепление в обоих парах признаков происходит независимо друг от друга, и при этом возможно перераспределение (рекомбинация) признаков с созданием сочетаний, не встречающихся у родительской пары.
Согласно третьему закону Менделя при скрещивании двух особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.
В потомстве все признаки распределяются независимо друг от друга.
Аллели разных генов наследуются независимо.
Независимое распределение генов обусловливается поведением хромосом при мейозе. Давайте вспомним, что при мейозе пары гомологичных хромосом (а следовательно, и парные гены, расходятся в разные гаметы независимо друг от друга).
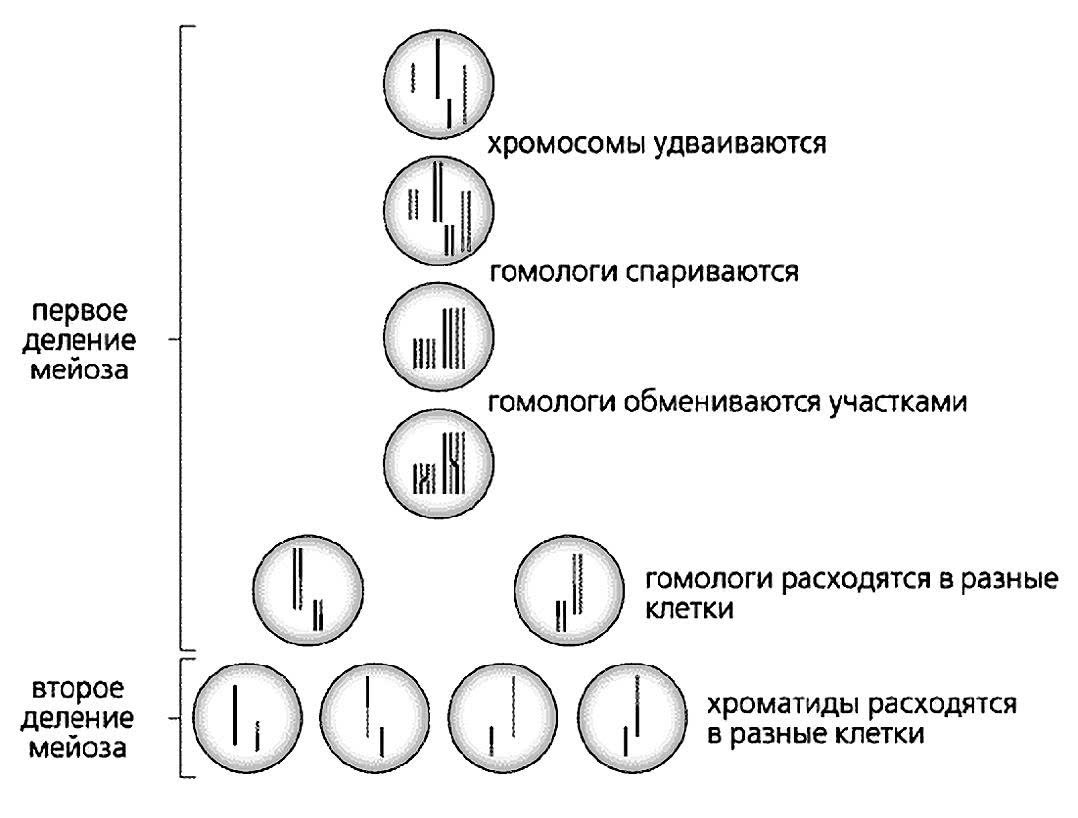
Мейоз
Уточнение, о котором Мендель так и не узнал: закон о независимом наследовании признаков при полигибридном скрещивании действует лишь в том случае, КОГДА РАЗНЫЕ ГЕНЫ РАСПОЛАГАЮТСЯ В РАЗНЫХ ПАРАХ ГОМОЛОГИЧНЫХ ХРОМОСОМ. Если изучаемые гены находятся в одной паре гомологичных хромосом, то расщепление в потомстве гибридов будет иным.
Так, например, если при дигибридном скрещивании оба гена входят в одну группу сцепления, то есть находятся в одной хромосоме и наследуются совместно, то вместо ожидаемого расщепления по фенотипу в соотношении 9:3:3:1, характерного для раздельного наследования, будет получено соотношение 3:1, как при моногибридном скрещивании. В примере с семенами гороха, который мы только что рассматривали, сцепление генов, определяющих цвет и гладкость семян, дало бы во втором поколении только гладкие желтые и морщинистые зеленые семена в соотношении 3:1. Семена были бы такими же, что и у родителей, без образования новых вариантов сочетаний.
Сцепленные гены, сколько бы их ни было в «связке», условно можно рассматривать как один ген и можно применять к ним все законы моногибридного скрещивания.
Закономерности сцепленного наследования были установлены уже известным нам Томасом Морганом. Морган исследовал дрозофилу, у которой гены, отвечающие за окраску тела и длину крыльев, находились в одной паре гомологичных хромосом и наследовались сцепленно.
Для определения фенотипов и генотипов потомства при дигибридном скрещивании английский генетик Реджинальд Пеннет предложил «решетку Пеннета» – удобный, наглядный графический метод. Решетка Пеннета представляет собой таблицу, по вертикальной оси которой размещены гаметы мужского родительского организма, а по горизонтальной – женского. В местах пересечения вертикалей и горизонталей записываются генотипы дочерних организмов. Таблица, изображенная на рисунке, изображающем расщепление при дигибридном скрещивании семян гороха – это и есть решетка Пеннета, очень удобная система, позволяющая избежать путаницы и ошибок. Если вы захотите исследовать наследование какого-то признака, то непременно ее используйте.
Все три закона Менделя универсальны для всех живых организмов, размножающихся половым путем, независимо от их видовой, родовой, классовой и прочей принадлежности.
Мендель также сформулировал гипотезу чистоты гамет, которая впоследствии была доказана и официально получила статус закона. Согласно этому закону в каждую гамету попадает только один аллель из пары аллелей данного гена родительской особи. Гаметы всегда остаются «чистыми» – они всегда несут только один ген, доминантный или рецессивный. Только один! Каким бы образом ни комбинировались аллели в потомстве, при образовании гамет они расходятся. Нормальная гамета всегда чиста от второго гена аллельной пары.
Почему о чистоте гамет сейчас было сказано несколько раз, на разные лады? Потому что это очень важный закон, определяющий все учение о наследственности. Минус на минус всегда дает плюс, а гаметы всегда чисты.
А теперь, опираясь на полученные знания, давайте рассмотрим наследование гемофилии, рецессивного аллеля, передающегося только с X-хромосомой и проявляющегося исключительно у мужчин (у женщин происходит подавление доминантным аллелем другой Х-хромосомы).
Вот вам задание. Миша и Маша – супружеская пара. Цвет глаз у обоих голубой, но у Машиной матери и у Мишиного деда по отцу глаза карие. Какой цвет глаз и в каком соотношении могут иметь дети Миши и Маши, если известно, что ген карих глаз является доминантным, а ген голубых – рецессивным? (Cм. таблицу «Некоторые доминантные и рецессивные признаки человека»).
Если вы всерьез собрались расписывать возможные комбинации, то вам, наверное, стоит еще раз перечитать эту главу с самого начала. Или же, как вариант, нужно лучше вникать в смысл заданий. У родителей с рецессивным признаком, вне зависимости от генотипов предков с обоих сторон, будут рождаться дети, имеющие рецессивный признак. Если генотип карих глаз может быть разным – ГГ или Гг, то генотип голубых глаз – только гг. Что тут рассуждать-высчитывать, если «гг» скрещивается с «гг»? Ничего, кроме «гг», в результате не получится.
Вот вам другое, на сей раз не шуточное задание. Миша и Маша – супружеская пара. Цвет глаз у обоих карий, но у Машиной матери и у Мишиного деда по отцу глаза голубые. Какой цвет глаз и в каком соотношении могут иметь дети Миши и Маши? Напишите все возможные комбинации для любых вариантов сочетания доминантных и рецессивных генов (карий цвет глаз может быть обусловлен как сочетанием АА, так и Аа). Тот, кто выполнит это задание, может считать себя продвинутым генетиком.
А теперь давайте ознакомимся с наследованием групп крови у человека. Это весьма интересный процесс, основанный на множественном аллелизме и таком явлении, как кодоминирование. При кодоминировании аллельные гены, оказавшиеся в одном генотипе, проявляются одновременно, без подавления рецессивного гена доминантным, поскольку оба они обладают одинаковой экспрессией, проще говоря – одинаковой силой.
Обратите внимание на то, что множественный аллелизм заключается в том, что в генофонде популяции существует более двух аллелей одного гена. Но в генотипе каждой особи присутствуют только два аллеля, поскольку хромосомы разбиты по парам, а не по тройкам или четверкам!
Множественность аллелей выражается в множестве их комбинаций. Если при двух аллелях может быть всего три комбинации (генотипа) – AA, Aa и aa, то при трех аллелях – A, a и a*) комбинаций будет уже шесть – AA, Aa, Aa*, aa, aa*, a*a*.
У гена, определяющего группу крови, существует три аллеля: IA, IB, I0, причем аллели IA и IB являются доминантными, а I0 – рецессивным (вообще-то аллелей этих на сегодняшний день известно более десятка, но мы будем рассматривать основные, а то запутаемся).
Гены IA и IB кодируют синтез белков-антигенов А или В, а ген I0 – никакого белка не кодирует. Белки-антигены находятся на поверхности эритроцитов. Эти белки способны связываться с антителами анти-А и анти-В, находящимися в плазме крови человека. Каждый антиген связывается со своим, одноименным антигеном, и в таком случае происходит распад эритроцитов. Стало быть, у одного человека не могут одновременно присутствовать антиген А и антитело анти-А или же антиген В и антитело анти-В. По этой же причине переливать можно только одногруппную кровь.
У человека с генотипом IAIB в крови будут присутствовать два антигена – А и В. Такую кровь называют кровью четвертой группы.
Люди с генотипом I0I0 не будут иметь в крови антигенов. Такую кровь называют кровью первой группы.
Вторую и третью группы крови определяют две комбинации генов – IAI0 и IAIA для второй, а для третьей – IBI0 и IBIB.
Соответственно:
– если оба родителя имеют первую группу крови, то у всех детей будет она же;
– если один из родителей имеет первую группу крови, а другой – вторую, то наследование группы крови детьми будет зависеть от генотипа второго родителя. Если он гомозиготный, то есть имеет комбинацию IAIA, то все дети будут иметь вторую группу – генотип IAI0. Если же второй родитель гетерозиготен (IAI0), то половина детей будет иметь первую группу крови (I0I0), а другая половина – вторую (IAI0).
– если один из родителей имеет первую группу крови, а другой – третью, расклад у детей будет аналогичным – либо все дети будут с третьей группой (IВI0), либо половина с третьей (IВI0), а половина с первой.
– если один из родителей имеет вторую, а другой – третью группы и при этом оба родителя гетерозиготны, то дети с равной вероятностью могут иметь любую группу крови.
Комбинации, возникающие при наследовании групп крови и их частоту удобнее и нагляднее изображать в виде таблицы. Генетики вообще очень любят таблицы.
Кстати говоря, мы с вами только что развенчали очень стойкий миф о том, что ребенок, имеющий группу крови, отличную от групп крови родителей, обязательно является неродным. У родителей с первой и четвертой группами крови неродной ребенок будет как раз иметь схожую группу, а не вторую или третью, как положено родному. Примерно половина добровольных обращений для установления отцовства по ДНК начинается со слов: «Непонятно почему у ребенка такая группа крови».
Как, по-вашему, почему при близкородственных браках значительно возрастает (по сравнению с неродственными) вероятность появления в потомстве различных отклонений от нормы – уродств и болезней?
Потому что родственники, имея общее происхождение, могут быть носителями одного и того же рецессивного патологического гена, следовательно, при близкородственных браках значительно возрастает риск рождения гомозигот по этому гену. Давайте «распишем» и сравним две комбинации – неродственную (АА х Аа) и родственную (Аа х Аа).
В первом случае расклад будет таким:
АА х Аа = АА + Аа
Пятьдесят процентов здоровых без носительства и пятьдесят процентов здоровых носителей рецессивного патологического гена а. Короче говоря, ни одного больного ребенка от этой пары родиться не может.
Во втором же случае расклад будет таким:
2Аа х 2Аа = АА + 2Аа + аа
То есть один из четырех детей родится больным.
Существует такой метод генетического анализа, как анализирующее скрещивание. Для того чтобы проверить, является ли данный организм гетеро- или гомозиготным по доминирующему аллелю, его скрещивают с анализатором – гомозиготой по рецессивным аллелям.
Проще говоря, если вы хотите узнать, с каким набором генов вы имеете дело в данном организме – АА или Аа, то вам надо скрестить этот организм с гомозиготой аа и оценить результат. Если рецессивный признак, кодируемый геном а, не проявится в первом поколении потомства, значит, вы имеете дело с гомозиготой по доминирующим аллелям – генотипом АА. Если же у половины потомства будет наблюдаться проявление рецессивного признака, значит вы имеете дело с гетерозиготой – генотипом Аа.
Вот уравнения для обоих случаев:
АА х аа = 2Аа
Аа х аа = Аа + аа
С аллельными генами мы более-менее разобрались, хоть и в самых общих чертах. Но генетикам известны случаи, когда подавление одного гена другим наблюдается между генами из разных аллелей. Да, представьте себе – из разных. Такое явление называется эпистазом, а ген-угнетатель, подавляющий фенотипические проявления другого неаллельного гена, – эпистатичным. Подавленный ген называется гипостатичным геном.
Как может происходить подавление в разных аллелях?
Прямым или косвенным путем. Белок или РНК, кодируемые эпистатичным геном могут препятствовать считыванию информации (то есть синтезу матричной РНК) с гипостатичного гена. Это прямой путь.
При косвенном продукт эпистатичного гена не дает проявиться соответствующему признаку в фенотипе. Так, например, у лошадей вороная масть определяется доминантным геном Р, а рыжая – рецессивным геном р. Но у них есть еще и ген раннего поседения С, который вызывает серую масть. При наличии в генотипе лошади доминантного аллеля С окраска всегда будет серой, вне зависимости от сочетания аллелей Р и р. Ген С не блокирует считывание информации с генов Р или р, но он вызывает раннее разрушение пигментов, кодируемых этими генами, то есть сводит на нет их проявление в фенотипе.
Интересная деталь – эпистатическое воздействие могут оказывать как доминантные, так и рецессивные эпистатичные гены. Говоря понятнее, эпистатичный ген не обязательно должен быть доминантным в своей аллели. Бывает и так, что доминантный ген не оказывает эпистатичного действия, а рецессивный – оказывает.
Примером доминантного эпистаза является наследование белой окраски у плодов тыквы. Ген В отвечает за желтую окраску плодов, ген b – за зеленую окраску, а доминирующий эпистатичный ген I подавляет гены В и b, вызывая белую окраску. Рецессивный ген i не оказывает влияния на окраску плодов. А вот у мышей белая окраска шерсти определяется по механизму рецессивного эпистаза – рецессивный эпистатичный ген i подавляет гены А и а, отвечающие соотвественно за серую и черную окраску шерсти. Доминантный ген I на окраску шерсти не влияет.
Явление эпистаза было открыто на основании множества «ошибок» в селекции лошадей. Селекционеры скрещивали лошадей так, как было нужно по науке, с учетом доминирования генов, для того чтобы получить в потомстве желаемую масть и ряд других признаков, но результат получался непредсказуемым. Одну-две ошибки можно списать на конюха – недоглядел, лентяй этакий, или спутал, короче говоря, подпустил к кобыле не того жеребца. Но если ошибок много, да вдобавок они укладываются в некую статистическую закономерность, то надо искать причину. Ученые поискали – и нашли.
Неаллельные гены могут не только враждовать, подавляя друг друга, но и сотрудничать, обеспечивая проявление одного признака в результате совместных усилий. Такое позитивное явление (а сотрудничество во имя благих целей может быть только позитивным), называют полимерией. Одним из самых распространенных примеров полимерии является наследование цвета кожи у людей. Ведущую роль в этом процессе играют четыре гена, эффект действия которых суммируется. Слово «ведущую» было употреблено не случайно, поскольку с точным количеством генов, определяющих цвет кожи, ученые пока еще не определились. Содружественные гены принято обозначать одной и той же буквой (одним и тем же кодом), несмотря на их принадлежность к разным аллелям – ведь определяют они один и тот же признак. Для отличия к буквенному коду добавляются цифры.
О том, что цвет кожи определяется целым набором генов, а не одной парой аллелей, ученые догадывались и до открытия явления полимерии. Уж больно много оттенков имеет человеческая кожа, не сравнить с цветом семян гороха, который использовал в своих экспериментах отец генетики Грегор Мендель.
Если ограничиться только четырьмя ведущими генами, то мы получим девять комбинаций, определяющих цвет кожи человека.
1. А1А1А2А2
2. А1а1А2А2
3. А1А1А2а2
4. А1а1А2а2
5. А1А1а2а2
6. а1а1А2А2
7. А1а1а2а2
8. а1а1А2а2
9. а1а1а2а2
Генотип А1А1А2А2 определяет наиболее темный цвет кожи, а генотип – самый светлый.
Вот вам задание, последнее в этой главе. Дайте аргументированный ответ на вопрос: может ли у пары мулатов с одинаковыми генотипами А1а1А2а2 родиться совершенно белокожий ребенок? При условии, что сцепленного наследования в этом случае нет.
ПОСТСКРИПТУМ. В генетике существует термин «возвратное скрещивание», который непосвященному человеку может показаться абсурдным.
В самом деле – как может скрещивание быть возвратным?
Оно к чему-то возвращается? Но как такое может быть? Ведь скрещивание – это движение вперед, к грядущему потомству…
Оно что-то возвращает? Но это тем более странно…
Возвратным скрещиванием, или беккроссом (от английского «backcross», означающего то же самое) генетики называют скрещивание гетерозиготных гибридов первого поколения (Аа), полученных от гомозиготных родителей (АА или аа) с родительскими особями или же с гомозиготными особями, сходными по генотипу с родительскими.
В чем смысл подобного скрещивания?
В том, чтобы дополнить (генетики говорят «насытить» генотип гибрида генами одного из родителей). Для насыщения генотипа нужно проводить многократные возвратные скрещивания с одной и той же гомозиготой. Потому и возвратное, что генотипы стараются вернуть к исходным вариантам. Условно говоря, сначала получили от АА и аа генотип Аа, а теперь хотим вернуть его к АА или аа. В принципе, можно было бы назвать возвратное скрещивание очищающим скрещиванием, поскольку в результате серии таких скрещиваний «грязные» гетерозиготные генотипы (Аа) очищаются от «примесей» – другого аллеля.
Разновидностью возвратного скрещивания можно считать анализирующее скрещивание, о котором было сказано в этой главе.

