Книга: Восхождение на гору Невероятности
Назад: Глава 6 Музей Всех Возможных Раковин
Дальше: Глава 8 Пыльца и самоуправляемые ракеты
Глава 7
Эмбриональный калейдоскоп
Тела формируются в процессе развития и роста – как правило, в эмбриональном периоде, – поэтому мутация, из‐за которой меняется форма тела, обычно корректирует процесс развития эмбриона. Например, мутация может ускорить рост какого‐то участка ткани в голове зародыша, а потом у взрослого человека будет длинное лицо с впалыми щеками. Сбой в раннем развитии может иметь шокирующие последствия – две головы или лишнюю пару крыльев. По причинам, которые мы обсуждали в третьей главе, естественный отбор вряд ли поддержит такие чудовищные мутации. Эта глава посвящена другой теме. Мы поговорим о зависимости типа мутаций, с которыми готов работать естественный отбор, от характера эмбрионального развития вида. Эмбрионы млекопитающих и насекомых развиваются по‐разному. Существуют и аналогичные, правда не настолько выраженные, отличия в эмбриональном развитии разных отрядов млекопитающих. Вы увидите, что можно выделить типы эмбрионального развития, в определенных отношениях более “способные” к эволюции. Это не значит, что для них более вероятны мутации, которые сами по себе тоже важны. Я хочу сказать, что те или иные варианты изменчивости, которые формируются в ходе некоторых типов эмбрионального развития, могут оказаться более перспективными для эволюции, чем варианты, продуцируемые другими типами развития. Более того, в результате отбора на более высоком уровне (эволюции способности к эволюции, как я назвал это ранее), мир может быть населен существами, которые обладают высокой способностью к эволюции благодаря особенностям своей эмбриологии.
В устах такого закоренелого дарвиниста, как я, это может показаться варварской ересью. В кругах строгих неодарвинистов считается, что естественный отбор не выбирает между крупными группами. И разве еще в третьей главе мы не пришли к выводу, что естественному отбору больше нравится нулевая частота мутаций – к счастью для будущего нашей планеты, недостижимая? Можно ли теперь утверждать, что эмбриональное развитие того или иного типа удобно для мутаций? Пожалуй, можно – и вот в каком смысле. Определенные типы развития имеют тенденцию варьировать определенным образом (отличным от других типов развития). И некоторые варианты могут быть плодотворнее других в плане эволюции – возможно, они с большей вероятностью продуцируют широкий спектр новых форм, как произошло с млекопитающими после исчезновения динозавров. Именно такой смысл я вкладывал в свой спорный тезис о “более способных” к эволюции типах эмбрионального развития.
Лучше всего сравнить это с калейдоскопом, разве что в калейдоскопе нас привлекает не практичный дизайн, а красочные, замысловатые картинки. В калейдоскопе цветные осколки собираются в кучки как попало. Но благодаря составленной с тонким расчетом системе зеркал мы любуемся геометрическими узорами, похожими на изящные снежинки. Осколки налетают на хаотически расположенные выступы на стенках трубы и перемешиваются. Мы же, глядя в окуляр, видим симметричные перемещения фрагментов на лучах снежинок. Мы крутим трубу, и она словно переносит нас в сокровищницу Аладдина.
Действие калейдоскопа основано на повторах событий в пространстве. Непредсказуемые изменения копируются на все четыре стороны. Но, в зависимости от количества зеркал, осей может быть не четыре, а сколько угодно. Эффект мутации тоже может повторяться в разных отделах организма, хотя сама мутация – это единичное изменение. Считайте это еще одним примером закономерности мутаций, помимо тех, о которых шла речь в главе 3. Сколько будет повторов – зависит от особенностей эмбрионального развития. Мы рассмотрим различные калейдоскопические картины в эмбриологии. Мой интерес к калейдоскопической эмбриологии подогрели опыты с селекцией биоморфов и особенно результаты внедрения “зеркал” в программу “Слепой часовщик” (см. ниже). Не красоты ради я привожу здесь столько иллюстраций с биоморфами и прочими компьютерными тварями.
Начнем с симметрии, а точнее – с ее отсутствия. Мы сами, как и большинство знакомых нам животных, в целом симметричны, пусть и с некоторыми изъянами, поэтому мы нередко забываем о том, что далеко не всем живым созданиям присуща симметричность. Асимметричны, в частности, отдельные группы Protozoa (одноклеточных организмов) – как их ни разрежь, не получишь двух одинаковых или зеркально отображающих друг друга частей. Как скажется мутация на идеально симметричном животном? Чтобы это понять, лучше всего обратиться к компьютерным биоморфам.
Четыре биоморфа на рис. 7.1а – это мутанты одного и того же вида, полученные в процессе эмбрионального развития, на которое не наложено ограничение со стороны симметрии. Симметричные формы не запрещены, просто нет цели создать именно их. Мутация просто меняет форму биоморфов, эффекта калейдоскопа и зеркальных отображений не наблюдается. Но взгляните на другие биоморфы (рис. 7.1.b). Здесь тоже показаны варианты мутаций одного вида, однако в эмбриональное развитие включено правило симметрии – в программу добавлено требование зеркального отражения относительно средней оси тела. Мутации могут затронуть что угодно, в том числе асимметричные биоморфы, но любые изменения в левой половине зеркально воспроизведутся справа. Такие формы выглядят более “жизненными”, чем асимметричные картинки на предыдущем рисунке.

Рис. 7.1. Количество “калейдоскопических зеркал” накладывает ограничения на вид и симметрию биоморфов.
Казалось бы, требование симметрии накладывает серьезные ограничения на “творческие” возможности эмбрионального развития, это своего рода барьер. Вообще‐то так и есть – раз без каких‐либо запретов зарождаются как симметричные, так и асимметричные формы, то в сумме форм получится больше. Но в этой главе мы увидим, что ограничение по симметрии дает противоположные результаты и на самом деле обогащает коллекцию эмбрионов. Проблема в том, что при эмбриональном развитии, на которое не наложены ограничения, надо перебрать несметное множество форм, прежде чем найдется одна симметричная. Но и тогда долгожданной симметрии в будущем угрожают мутации. При обязательном стремлении к симметрии, независимо от прочих возможных изменений, эмбриональное развитие, на которое действует такое ограничение, будет более продуктивным, да и глазу приятнее. Вам не придется тратить уйму времени на сортировку бесполезных несимметричных форм, что было бы неизбежно при ничем не ограниченном эмбриональном процессе.
По правде говоря, большинство живых существ, включая и нас с вами, не отличается идеальной симметричностью левой и правой сторон. Сама по себе красота – не главное, поэтому возникает резонный вопрос: чем так уж хороша зеркальная симметрия c практической точки зрения? Среди зоологов есть приверженцы выдвинутой в восемнадцатом веке теории “плана строения” (Bauplan), согласно которому развиваются анатомические особенности животных, в том числе симметричность тела. Bauplan – это чертеж или план по‐немецки. Считается, что в иностранной речи больше значительности – как язвительно заметил сэр Питер Медавар, “словно из глубин Рейна раздаются трубные звуки”. Но, строго между нами, в самом слове “план” скрыта насмешка, ибо оно подразумевает как раз упрощенный, механический перенос, взаимно-однозначное соответствие строения проекту, что, с точки зрения генетики, оскорбляет тех самых фанатов термина Bauplan в лучших чувствах. Мне гораздо симпатичнее англо-саксонская простота моего коллеги, доктора Генри Беннета-Кларка, который сказал, когда мы обсуждали эту тему: “На все вопросы ответ один – естественный отбор, хотя и не всегда это что‐то проясняет”. Разумеется, каждый вид выигрывает от зеркальной симметрии по‐своему, но доктор Беннет-Кларк формулирует базовый принцип, который касается того, о чем сейчас пойдет речь.
Как правило, животные либо сами червеобразны, либо происходят от червеобразных предков. Вдумайтесь, что за существо такое – червь; на одном конце (с которого поступает пища) у него рот, а на другом – задний проход, откуда должны исторгаться отходы пищеварения, иначе пришлось бы их съедать. Так определяются передняя и задняя стороны. Далее, верх и низ обычно также различаются в немалой степени, хотя бы из‐за действия земного притяжения. В частности, многие животные передвигаются по твердой поверхности – по суше или морскому дну. Как ни крути, разумно было бы, чтобы ближайшая к земле часть тела животного отличалась от той, что обращена к небу. Стало быть, дорсальный (спинной) и вентральный (брюшной) отделы неодинаковы и, раз уже известно, где перед и зад (голова и хвост), можно определить, где левая сторона, а где правая. Но почему левая и правая половины должны быть зеркальным отображением друг друга? А почему бы и нет? Понятно, почему несимметричны зад с передом и верх с низом, но нет ни одной веской причины полагать, что самая подходящая форма для левой стороны не подойдет наилучшим образом и для правой. Напротив, если для левой половины тела можно выбрать оптимальную форму, то логично предположить, что и для правой она окажется оптимальной. Точнее, любое существенное отклонение от зеркальной симметрии может привести к тому, что животное, вместо того чтобы идти из одной точки в другую кратчайшим путем, начнет нарезать круги.
Поскольку, неважно почему, параллельное – синхронное и симметричное – развитие левой и правой сторон предпочтительно, более выгодным становится “калейдоскопический” эмбриональный процесс с единственным “зеркалом” посередине. Сколько‐нибудь полезные новые мутации будут автоматически воспроизводиться слева и справа. Какова же альтернатива развитию по калейдоскопическому механизму? Поначалу в генетической линии может обнаружиться выгодное изменение – скажем, слева. До следующей такой же мутации сменится еще много асимметричных поколений. Очевидно, калейдоскопический процесс имеет преимущества. Видимо, калейдоскопический процесс эмбрионального развития с устойчивыми – и в то же время продуктивными – лимитирующими факторами благоприятен для естественного отбора определенного типа.
Это вовсе не означает, что не встречается асимметрия левой и правой сторон. Бывает, что мутации сильнее проявляются с одной стороны. Иногда по ряду причин асимметричные мутации даже желательны – скажем, брюшко рака-отшельника должно быть приспособлено к форме витой раковины, – поэтому естественный отбор к ним благосклонен. В 4 главе мы говорили о камбаловых рыбах (рис. 4.7), к ним относятся собственно камбала, морской язык и палтус. Некогда левый бок камбалы оказался у нее внизу, и левый глаз переместился на прежний правый бок, превратившийся в верхнюю часть туловища. Такой же перестройке подверглись и морские языки, только некоторые их виды лежат на правом боку, что указывает на вероятность, хотя и не стопроцентную, независимого формирования этой анатомической особенности. Изначально левый бок камбалы выполнял функциональную роль нижней, придонной стороны – как и следовало ожидать, он стал более плоским и обрел серебристую окраску. Бывший правый бок – на практике верх туловища – обращен к небу, поэтому он выпуклый и пестрый, под цвет дна. Унаследованные от предков дорсальная (спинная) и вентральная (брюшная) зоны поменяли свою функциональную роль и стали левым и правым боками. Соответствующие этим сторонам спинной и анальный плавники стали почти одинаковыми, просто левым и правым, хотя у других рыб они, как правило, выглядят совершенно по‐разному. Заново обнаруженная зеркальная (право-левая) симметрия у камбаловых рыб подчеркивает могущество естественного отбора в противовес универсальным и нерушимым анатомическим планам. Любопытно было бы взглянуть – и это осуществимо, – воспроизводятся ли автоматически проявления мутаций у камбалы слева и справа (спереди и сзади, если по‐старому). Или они все еще воспроизводятся одинаково на прежних левой и правой сторонах, то есть сверху и снизу? Почему столь различны оказались серебристая и пестрая стороны туловища камбалы – вопреки неблагоприятному старому эмбриональному калейдоскопу или при содействии нового, благоприятного? В любом случае ответ на этот вопрос подтверждает, что можно говорить о “благоприятном” и “неблагоприятном” для эволюции эмбриональном развитии. И еще раз спрошу: можно ли предположить, что существует естественный отбор более высокого уровня, который делает эмбриональное развитие еще “более дружественным” для определенных направлений эволюции?
В контексте этой главы на первый план выходит одна особенность зеркальной (право-левой, или билатеральной) симметрии – мутация сказывается не в одной зоне организма животного, а сразу в двух одновременно. Это я и называю калейдоскопической эмбриологией – мутации будто отражаются в зеркалах. Но билатеральная симметрия – не единственный вариант. Мутационные зеркала могут располагаться и в других плоскостях. Биоморфы на рис. 7.1с симметричны как по ширине, так и по длине. Словно бы установили перпендикулярно два зеркала. Реальные создания с “двухзеркальной” эмбриологией встречаются в природе гораздо реже, чем симметричные относительно одной плоскости. Таков, например, венерин пояс, дивный мираж, лентовидный планктонный организм, который принадлежит к типу гребневиков, или ктенофор (тип Ctenophora). Более распространены эмбриологические калейдоскопы с четырьмя плоскостями симметрии по четырем направлениям, как на рис. 7.1d. Такая геометрия присуща многим медузам, которые относятся к типу книдарий (Cnidaria). Представители этого типа либо плавают в море (как сами медузы), либо, как актинии, цепляются за морское дно, поэтому не испытывают сжатия по оси, характерного для ползающих существ – например, червей. У них были все основания сформировать различающиеся верхнюю и нижнюю стороны тела, но по направлениям от переднего конца к заднему и справа налево никакие силы не действуют. Поэтому, если смотреть сверху, непонятно, с чего бы какой‐нибудь одной стороне тела отличаться от любой другой – и они действительно обладают радиальной симметрией. Изображенные на рис. 7.2 медузы обладают четырехлучевой радиальной симметрией, но, как мы еще увидим, осей может быть больше. Автор этого и многих других рисунков в этой главе – знаменитый немецкий зоолог и талантливый художник-иллюстратор Эрнст Геккель.
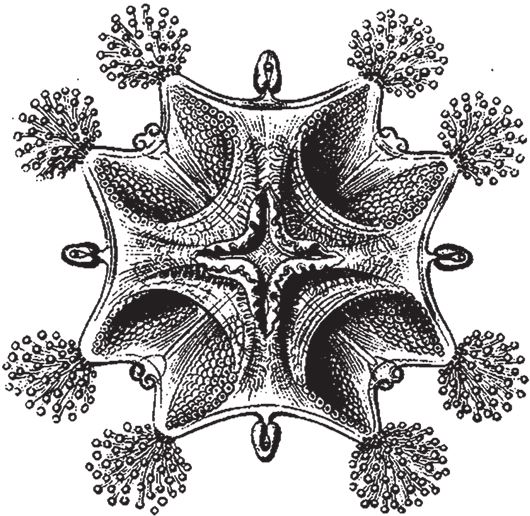
Рис. 7.2. Ставромедуза, животное с четырехлучевой симметрией. Обратите внимание на зеркальную симметрию всех лучей, таким образом, максимальное число отражений – восемь.
Симметрия такого типа приводит к бесконечному разнообразию форм, но с одним ограничением, которое, впрочем, как и “калейдоскоп”, может оказаться полезным. Случайные изменения будут затрагивать одновременно все четыре сектора. Поскольку четырежды воспроизведенные элементы часто еще и зеркально отражаются, эффект каждой мутации повторяется восемь раз. Это хорошо видно на примере ставромедузы (рис. 7.2): на концах ее “рук” имеются щупальца, собранные в кисточки, по две кисточки на каждой руке, всего восемь. Можно предположить, что эффект мутации, влияющей на форму кисточки, воспроизвелся бы восемь раз. По геометрии биоморфов, показанных на рис. 7.3, можно судить о том, как выглядела бы фигура с радиальной симметрией того же типа без дополнительного удвоения. Вряд ли вы найдете животное с симметрией такого типа, столь похожее на свастику или на остров Мэн, однако на рис. 7.3 изображено нечто в этом роде. Это сперматозоид речного рака.
Большинство животных с лучевой симметрией обладает еще и зеркальной симметрией относительно лучей, сколько бы их ни было. Для нас имеет значение кратность воспроизведения эффекта одной мутации, поэтому мы должны посчитать лучи и умножить это число на два. Можно предположить, что у типичной морской звезды с пятью зеркально-симметричными концами (“руками”) каждая мутация “отражается” десять раз.
Геккель особенно увлекался рисованием одноклеточных организмов, таких как диатомеи (рис. 7.4). Перед нами калейдоскопические узоры, полученные “с помощью” двух, трех, четырех, пяти и более “зеркал” в сочетании с отражением левой и правой сторон каждой “руки”. При всех типах симметрии для эмбрионального развития характерна повторяемость эффектов мутаций в нескольких определенных зонах. Например, пятилучевая звезда в верхней части рисунка 7.4 могла мутировать так, что ее концы стали острее. Тогда все пять лучей должны заостриться синхронно. Нам не нужно было бы сидеть и ждать, когда произойдут пять независимых мутаций. Возможно, изменение количества зеркал – это тоже мутация, хотя гораздо более редкая. Может, когда‐то в результате случайной мутации трехконечная звезда превратилась в пятиконечную.
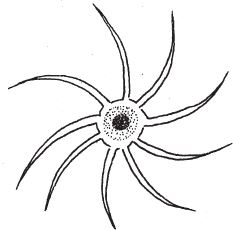
Рис. 7.3. Аксиальная симметрия: сперматозоид речного рака.

Рис. 7.4. Диатомовые водоросли, микроскопические одноклеточные растения, примеры калейдоскопических отражений в одной группе организмов с разным количеством зеркал.
На мой взгляд, в мельчайшей категории калейдоскопов лидирует радиолярия, член другой планктонной команды, которой Геккель уделил особое внимание (рис. 7.5). В этой группе мы тоже видим симметрию разного порядка – подобные картинки можно было бы получить с двумя зеркалами, тремя, четырьмя, пятью, шестью и даже больше. Микроскопические меловые скелеты радиолярий так изысканны и гармоничны, что невольно вспоминаешь о калейдоскопической эмбриологии.
Калейдоскопический шедевр с рис. 7.6 достоин карандаша Бакминстера Фуллера, архитектора и мечтателя; однажды, когда ему уже перевалило за девяносто, мне посчастливилось попасть на его лекцию – это было трехчасовое завораживающее шоу без единой паузы. Прочность изображенной здесь конструкции, как и геодезических куполов Фуллера, обеспечивается строгой геометрией ее треугольных компонентов. Это, несомненно, продукт калейдоскопической эмбриологии высшего порядка. Эффект любой отдельно взятой мутации отразится многократно – сколько раз, по картинке понять нельзя. Другие радиолярии Геккеля химики используют в кристаллографии как примеры известных с древних времен правильных геометрических тел – октаэдра (включает 8 треугольных граней), додекаэдра (12 треугольников) и икосаэдра (20 треугольников). Даже Дарси Томпсон (мы познакомились с ним, когда обсуждали раковины улиток) доказывал, что в развитии безупречных форм радиолярий больше от процесса роста кристаллов, нежели от эмбриогенеза в общепринятом смысле слова.

Рис. 7.5. Радиолярия. Еще примеры калейдоскопических отражений с разным количеством зеркал в группе микроскопических одноклеточных организмов.

Рис. 7.6. Изысканный скелет крупной радиолярии.
Так или иначе, многоклеточные организмы и одноклеточные – такие как диатомеи и радиолярии – в эмбриональном периоде развиваются совершенно по‐разному, и если между этими калейдоскопическими процессами есть какое‐то сходство, то по чистой случайности. Мы рассматривали пример многоклеточного животного, медузы, с четырехлучевой симметрией. Число лучей четыре и кратное четырем свойственно медузам, это легко реализуется за счет дупликации некоторых процессов в ходе эмбрионального развития. Трахимедузы из класса гидроидных (класс Hydrozoa) обладают шестилучевой симметрией (рис. 7.7).
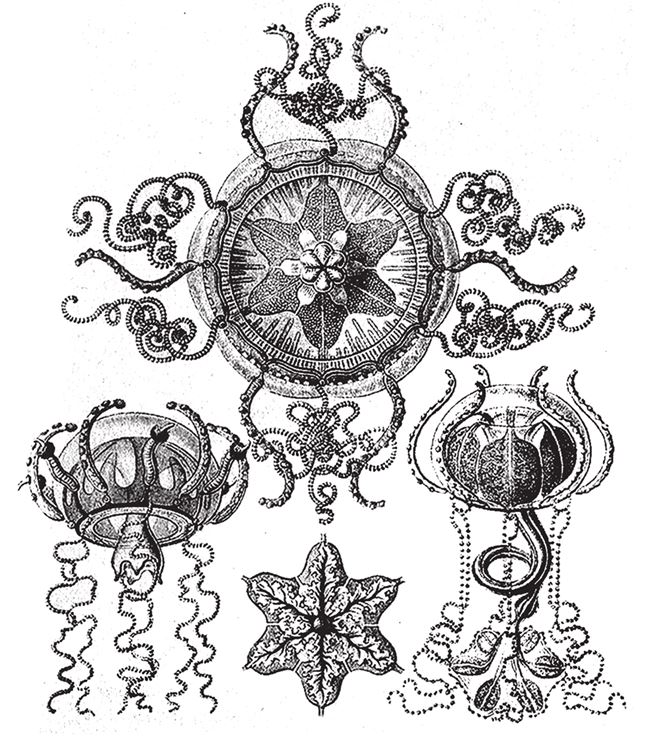
Рис. 7.7. Медузы с шестилучевой симметрией.
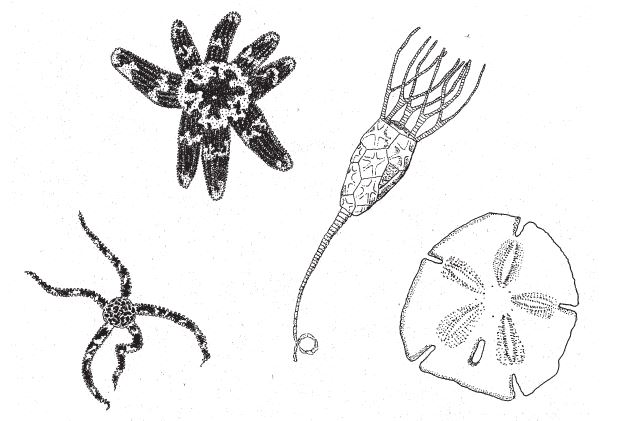
Рис. 7.8. Иглокожие из разных групп; слева направо: офиура, морская звезда (с неодинаковыми лучами – возможно, из-за травмы и последующего восстановления), морская лилия, плоский морской ёж.
Иглокожие – самые типичные образчики пятилучевой симметрии; к этому обширному типу (тип Echinodermata) колючих обитателей морей относятся морские звезды, морские ежи, офиуры, голотурии (морские огурцы) и морские лилии (рис. 7.8). Есть мнение, что их древними предками были животные с трехлучевой симметрией, но вот уже свыше полумиллиарда лет они живут с пятью лучами, и логично предположить, что пятилучевая симметрия составляет основу одной из наиболее консервативных схем строения тела, столь милых глобальному мышлению некоторых зоологов. Однако в природе немало морских звезд, у которых количество рук отлично от пяти; более того, даже среди добропорядочных пятируких видов попадаются отдельные особи с трех-, четырех- и шестилучевой симметрией, что противоречит такой идеалистической точке зрения.
С другой стороны, лучевая симметрия характерна даже для ползающих иглокожих, хотя от животного, которое всю жизнь копошится на морском дне, этого совсем не ждешь. И видимо, лучевая симметрия – вовсе не пустяк для них: все их руки одинаковы, и им все равно, куда идти. В каждый момент времени одна из рук морской звезды играет роль ведущей, но потом другая рука становится главнее. За долгие годы эволюции некоторые иглокожие “заново открыли” для себя билатеральную симметрию. У сердцевидных и плоских морских ежей, которые зарываются в песок и, очевидно, в борьбе с его массой должны были обрести обтекаемые контуры тела (в целом явно те же, что у морских ежей с пятилучевой симметрией), сформировалась передне-задняя асимметрия и, сверх того, лево-правая асимметрия.
Пытаясь “вывести” с помощью “Слепого часовщика” правдоподобные биоморфы, я хотел сделать их похожими на настоящих, фантастически красивых иглокожих. Однако все мои попытки воссоздать пятилучевую симметрию провалились. Эмбриологический калейдоскоп “Слепого часовщика” оказался не тем, что нужно. Ему недоставало необходимого количества “зеркал”. Но, как мы знаем, среди иглокожих встречаются “белые вороны”, уклонившиеся от обязательной пятилучевой симметрии, и я немного смухлевал – нарисовал морских звезд, офиур и морских ежей с четным числом лучей (рис. 7.9).
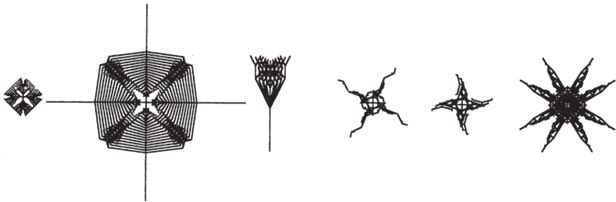
Рис. 7.9. Компьютерные биоморфы – почти такие же, как настоящие иглокожие, только вот пятилучевую симметрию воссоздать не удается. Для этого пришлось бы переписать всю программу.
Ничего не поделаешь – существующая версия программы “Слепой часовщик” не позволяет разводить биоморфы с пятилучевой симметрией (этот факт подтверждает главную идею данной главы). Чтобы исправить недочет, мне пришлось бы переписать программу (не просто изменить число мутаций для имеющегося в наличии гена, а добавить еще одно “зеркало”) и ввести новый тип калейдоскопических мутаций. Я уверен, что при этом можно было бы добиться гораздо большего сходства с большинством основных групп иглокожих за счет обычных случайных мутаций и отбора, пусть и длительного. С помощью описанной в “Слепом часовщике” первой версии программы удается воспроизвести лишь те мутации, что зеркально отражаются слева направо. Я решил переделать ее – запрограммировать серию “зеркал” с генетическим контролем, – и сейчас с помощью этого открытого для пользователей ресурса можно создавать биоморфы с четырехлучевой симметрией, в виде свастики и трехконечной “вертушки”.
Я рассказал вам о разных типах симметрии, чтобы объяснить суть калейдоскопической эмбриологии. В живой природе есть еще один не менее важный, хотя не такой зрелищный, феномен – сегментация (метамерия). Сегментация – это сериальное повторение элементов строения организма вдоль всего тела, и чаще она наблюдается у животных с длинным, зеркально симметричным телом. Первыми вспоминаются аннелиды (кольчатые черви, сухопутные земляные черви, а также морские кольчатые черви – пескожилы, нереиды и другие) и членистоногие (насекомые, ракообразные, многоножки, трилобиты и прочие), но и позвоночным вроде нас тоже присуща метамерия, хотя несколько иного рода. Тело членистоногого представляет собой последовательный ряд сегментов с теми или иными особенностями – вроде железнодорожного состава, в котором все вагоны примерно одинаковые, но с небольшими отличиями. Многоножка подобна товарному поезду с похожими как две капли воды вагонами. Других членистоногих можно считать улучшенными моделями многоножек: в их поездах каждая открытая платформа и каждый вагон имеет свое назначение (рис. 7.10).
Строение тела многоножки примитивно и однообразно. По всей длине “состава” повторяются объемные, зеркально симметричные фрагменты. Но если отойти дальше от многоножек и их разновидностей, станет заметна устойчивая эволюционная тенденция к уменьшению сходства между сегментами – не все эффекты мутаций будут просто повторяться в каждом сегменте. Насекомые напоминают многоножек, у которых ножки остались только в трех сегментах – седьмом, восьмом и девятом, считая от головы. У пауков конечности сохранились в четырех сегментах. На самом деле и пауки, и насекомые помимо своих собственно ножек частично сохранили и первичные конечности. Просто они используют их иначе – например, как антенны или челюсти. Омары и, скорее даже, крабы продвинулись еще дальше по пути дифференциации сегментов.
У гусениц ближе к голове есть три пары “правильных” ножек, какие и подобает иметь насекомому, а вот задние пары также подверглись реформам. Эти измененные ножки мягче состоящих из сочлененных фрагментов типичных крепких ног, расположенных в трех грудных сегментах, да и по другим признакам они тоже отличаются от настоящих. В седьмом и восьмом сегментах у насекомых, как правило, имеются крылья. Некоторые насекомые бескрылые, и их предки никогда не имели крыльев. Другие – например, блохи и рабочие муравьи – за время долгой эволюции утратили крылья, хотя их предки были крылатыми. У муравьев есть генетический аппарат для формирования крыльев – любая самка, если растить ее должным образом, может стать маткой (царицей), а у матки крылья есть. Любопытно, что муравьиная матка в течение жизни теряет крылья и даже обламывает их сама по окончании брачного полета, прежде чем спрятаться в муравейнике. Под землей крылья мешают, как и среди лесной растительности или в шерсти животных и перьях птиц, где обычно находят прибежище блохи.
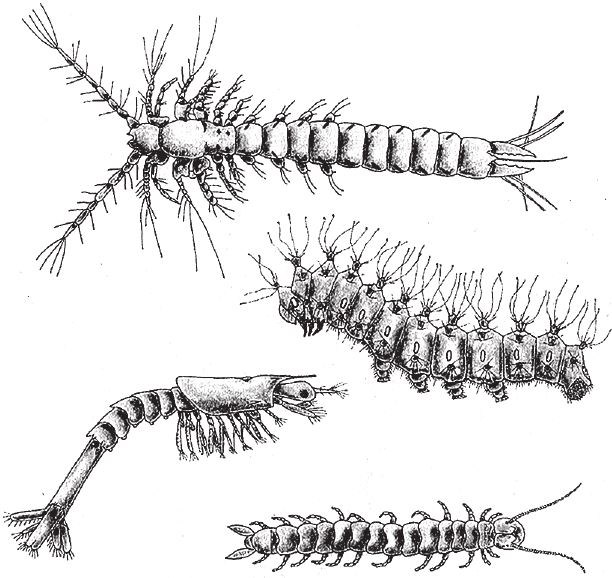
Рис. 7.10. У членистоногих сегменты повторяются от головы к хвосту, зачастую с некоторыми вариациями: ракообразное подкласса мистакокарид, Derocheilocaris; гусеница большого павлиньего глаза, Saturnia Pyri; десятиногий рак, Penaeus; многоножка из группы симфила, Scutigerella.
Блохи избавились от обеих пар крыльев, а вот мухи, несметное множество видов которых, включая комаров, составляет колоссальное семейство Diptera, лишились только одной пары, а вторую сохранили. Первая осталась в сильно редуцированном виде – это жужжальца, похожие на крошечные молоточки, которые торчат позади основных крыльев (рис. 7.11). Жужжальца не способны выполнять функции крыльев, это ясно и без технического образования. Но вот зачем все же нужны жужжальца, без хорошего технического образования не понять. По-видимому, они служат миниатюрным стабилизирующим устройством и играют ту же роль для насекомого, что и гироскоп для самолета и ракеты. Жужжальца вибрируют с той же частотой, с какой насекомое машет крылышками. Малюсенькие датчики у основания жужжалец реагируют на вращающие силы, направленные по трем осям, – как говорят в авиации, на крен по продольной и поперечной оси и на рысканье. Все в духе эволюции – приспособиться и пользоваться тем, что уже есть. Авиаконструктор проектирует стабилизирующее устройство, вычерчивая его на кульмане с нуля. Эволюция модифицирует то, что уже имеется, в данном случае крыло, и достигает того же результата.
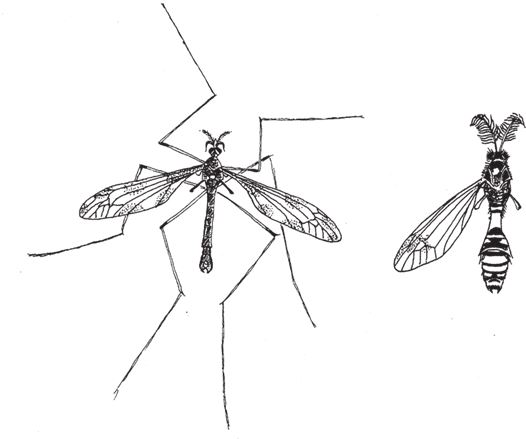
Рис. 7.11. У всех членов семейства Diptera (комары, мухи) вторую пару крылышек заменяют жужжальца. У комара-долгоножки они видны особенно отчетливо. Слева Tipula maxima, справа Ctenophora ornate (ножки и правое крыло не показаны).
В простом видоизменении сегментов ничего нет от калейдоскопа – как раз наоборот. Но к калейдоскопическим (в менее прямолинейном смысле, чем мы имели в виду до сих пор) можно отнести изменения иного рода. Строение тела членистоногого можно сравнить с предложением, в котором есть круглые скобки (если в предложении открываются скобки <вроде этих>, то их обязательно следует закрыть). Фразу в скобках можно сократить или дополнить, но независимо от ее сложности каждой открывающей скобке – знаку ‘(’ – должна отвечать закрывающая – знак ‘)’. Точно так же надо расставлять кавычки, по правилам грамматики. Что еще интереснее, этот закон распространяется и на выделяемые конструкции внутри предложения. Начало фразы “учитель, севший на кнопку…” аналогично открывающей скобке и требует сказуемого. Можно сказать: “Учитель подскочил” или “учитель, севший на кнопку, подскочил”, но нельзя оборвать фразу на полуслове – если только это не ответ на вопрос и не название произведения живописи, когда понятно, к чему вы клоните. В грамматике предложения должны быть правильно построены. По той же причине ближе к голове креветки, омара и краба всегда расположены шесть первых сегментов, а сзади всегда есть тельсон – последний особый сегмент. Все, что в середине, может варьироваться.
Мы изучили один вид калейдоскопических эффектов мутаций – зеркально отображенных относительно разных плоскостей симметрии. “Грамматические” эффекты мутации роднит с калейдоскопом другое свойство. Опять‐таки, не все вариации разрешены, но в данном случае лимитирует не симметрия, а принцип, который можно сформулировать так: при любом количестве суставов на конце ноги должна быть клешня. Мы с Тедом Келером, сотрудником корпорации Apple, разработали программу с учетом правил такого рода. Она напоминает “Слепого часовщика”, но производит “животных”, которых мы назвали артроморфами и в эмбриональном развитии которых действуют правила, отсутствующие в программе для биоморфов. “Тела” компьютерных артроморфов, как и живых членистоногих, представляют собой цепочку сегментов. Каждый сегмент – это округлая часть “тела”, а его форма и размер определяются такими же “генами”, как и у биоморфов. По бокам каждого сегмента могут торчать (а могут и не торчать) ножки, состоящие из члеников. За это – а также за толщину ног, количество, длину и углы соединения члеников – тоже отвечают гены. От генов же зависит, будет ли на конце ножки клешня, а если будет – то какой формы.
Если бы эмбриональное развитие артроморфов шло так же, как у биоморфов, то количество сегментов должен был бы регулировать некий ген – Nseg. Это было бы просто число, и оно могло бы меняться. При Nseg = 11 у животного будет 11 сегментов. Должен быть и другой ген – Njoint, – определяющий количество суставов конечности. При всем внешнем разнообразии биоморфов, показанных на рис. 1.16 (я чрезвычайно рад, что они получились такие разные, и горжусь этим), у всех ровно по шестнадцать генов. Первые биоморфы, созданные в программе “Слепой часовщик”, имели всего по девять генов. У цветных биоморфов генов больше – тридцать шесть, – и под них пришлось переписать всю программу. В итоге получились три разные программы. Артроморфы устроены иначе. У них нет фиксированного набора генов. Их генетическая система более лабильна (вряд ли кому‐нибудь, кроме увлеченных программистов, интересно, что информация о генах артроморфов хранится в виде связного списка с указателями, а о генах биоморфов – как запись на Паскале с фиксированным форматом). В ходе эволюции артроморфов путем дупликации старых генов могут спонтанно появляться новые. Иногда гены дуплицируются по одному. Иногда – в иерархически структурированных кластерах. Это означает, что теоретически у ребенка-мутанта может оказаться вдвое больше генов, чем у его родителя. Если в результате дупликации появляется новый ген или группа генов, их начальные численные значения равны тем, что были до дупликации. Возможна мутация другого типа – делеция (выпадение), – поэтому генов может стать как больше, так и меньше. Дупликации и делеции проявляются в виде изменения формы тела, а стало быть, поддаются отбору – визуальной селекции, как и в случае с биоморфами. Изменение количества генов часто выражается в изменении количества сегментов (рис. 7.12). Кроме того, может измениться число суставов в конечности. В обоих случаях идея та же, что и в грамматике – можно прицеплять или отцеплять вагоны в середине, в то время как голова и хвост состава остаются неизменными.
Дупликация и делеция сегментов могут происходить не только у концов, но и в средних отделах животного. Точно так же дупликация и делеция суставов конечностей возможны в середине и на периферии. Можно убирать и вставлять сегменты, словно придаточные предложения в середине длинной фразы, то есть говорить о “грамматике” эмбриологии артроморфов. Помимо того, что артроморфы подчиняются правилам “грамматических” вставок, к ним применимы и принципы калейдоскопической эмбриологии. На каждый количественный параметр тела артроморфа (например, на угол той или иной клешни или ширину того или иного сегмента) влияют три гена, численные значения которых увеличиваются – как именно, я сейчас объясню. Один ген влияет на данный сегмент, второй – на все животное в целом, третий – на подотделы сегментов, так называемую тагму. Этот термин взят из настоящей биологии. Грудь и брюшко настоящих насекомых – тоже тагмы.
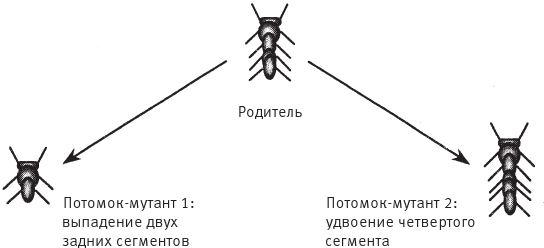
Рис. 7.12. Артроморфы с разным количеством сегментов. Родитель (вверху) дал потомков с двумя различными мутациями.
На любой отдельно взятый орган – скажем, клешню – оказывают влияние все три гена в совокупности. Во-первых, каждый сегмент имеет свой индивидуальный ген. Тут нет калейдоскопического эффекта, так как мутация этого гена меняет только данный сегмент. На рис. 7.13а показан артроморф, каждому сегменту которого отвечают разные значения гена для угла клешни в этом сегменте. Поэтому углы клешни во всех сегментах разные. Кстати сказать, все артроморфы обладают билатеральной симметрией.
Второй из трех генов влияет на все сегменты животного в целом, в том числе и на угол клешни. При его мутации одновременно меняются все клешни во всех сегментах по всей длине животного. Артроморф, показанный на рис. 7.13b, отличается от своего предшественника с рис. 7.13а только слегка недоразвитыми, укороченными клешнями. Ген, который влияет на размер клешни на уровне всего животного, мутировал (его значение стало меньше). Из-за этого уменьшилась каждая клешня по отдельности, но во всем прочем в каждом сегменте клешни не изменились относительно друг друга. Как я уже говорил, математически это выражается умножением численного значения гена для угла клешни в данном сегменте на численное значение гена для угла клешни животного в целом. Конечно, угол клешни – лишь один из многих количественных параметров, которые синхронно меняются во всем “поезде” в зависимости от совокупного эффекта разных факторов. Так, гены, влияющие на длину ног на уровне всего тела, перемножаются с генами, влияющими на длину ног в данном сегменте. На рис. 7.13c и 7.13d показаны артроморфы, у которых отсутствуют вариации сегментов, но отличаются гены угла клешни на уровне целого организма.
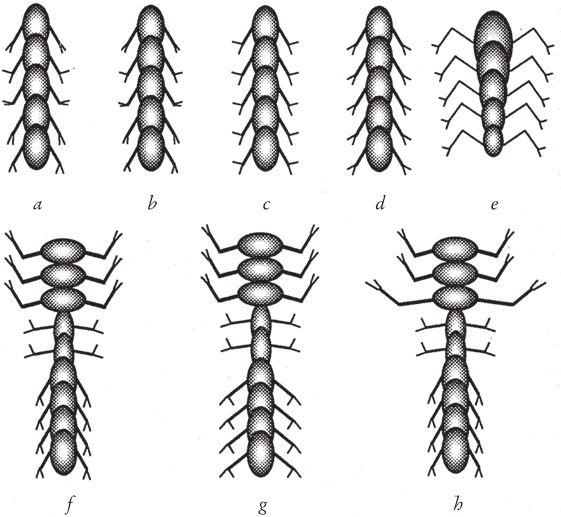
Рис. 7.13. Артроморфы, отобранные для иллюстрации влияния генов: (а) артроморф, в каждом сегменте которого проявились разные варианты гена угла клешни; (b) мутация гена угла клешни для всего тела; (с) артроморф с абсолютно одинаковыми сегментами; (d) единичная мутация повлияла на ген угла клешни для всего тела артроморфа (с); (е) размер туловища меняется от головы к хвосту, но конечности не затронуты мутацией; (f) артроморф с тремя отличающимися тагмами, но в пределах одной тагмы все элементы одинаковы; (g) тот же артроморф с измененными конечностями в третьей тагме; (h) артроморф (f) с измененными конечностями в одном сегменте.
Третий класс генов влияет на отдельные участки тела, тагмы, например, на грудной отдел членистоногого. У насекомых три тагмы, но артроморфы могут иметь их сколько угодно, причем в каждой тагме может быть сколько угодно сегментов – по законам нашей “грамматики” мутациям подвержены как количество сегментов в целом, так и их количество в тагме. Каждой тагме соответствует набор генов, определяющих форму тела, конечностей и клешней в ее пределах. На рис. 7.13f изображен артроморф с тремя тагмами. Как правило, различия между тагмами более заметны, чем различия внутри одной тагмы. Как и в рассмотренном уже случае воздействия генов всего организма, эффект дает перемножение величин генов.
Итак, каждая характеристика – например, угол клешни – в количественном отношении определяется численными значениями трех генов, а именно: сегментного гена угла клешни, гена угла клешни для данной тагмы и гена угла клешни для всего организма. Умножение на ноль дает тот же ноль, отсюда – если в данной тагме ген размера конечности равен нулю, то в ее сегментах вовсе не будет конечностей (как на брюшке осы) независимо от величин двух других генов. На рис. 7.13g вы видите дочку артроморфа с рис. 7.13f, чей ген размера конечностей для третьей тагмы изменен. На рис. 7.13h – еще одна дочка того же артроморфа с рис. 7.13f, но у нее изменился только ген конечностей для одного сегмента.
Таким образом, эмбриональное развитие артроморфов представляет собой как бы трехуровневый калейдоскопический процесс. Возможны изменения в пределах одного сегмента – тогда эти изменения отображаются на другой стороне тела один раз. Возможно калейдоскопическое отображение на уровне всего организма, как бы в масштабе многоножки, – при этом эффекты мутации повторяются во всех сегментах, а заодно отражаются зеркально на противоположной стороне. И бывают промежуточные калейдоскопические “мутации насекомого” (на уровне тагмы), которые влияют на сегменты в локальной группе и не влияют на остальные сегменты. Думаю, если бы артроморфам пришлось бороться за жизнь в реальном мире, то, по тем же соображениям экономики эволюции, которые мы обсуждали в связи с зеркальной симметрией, такой трехуровневый калейдоскоп дал бы им преимущества. Допустим, если конечности средней тагмы выполняют функции ходильных ног, а задней – жабр, то усовершенствования, приобретаемые в ходе эволюции, сериально повторялись в пределах сегментов одной тагмы, не затрагивая другую. Едва ли дыхательные конечности выиграют от тех же усовершенствований, что и ходильные конечности. Стало быть, такие мутации, которые при первом же проявлении отражаются во всех сегментах тагмы, могут быть полезны. С другой стороны, можно извлечь дополнительную выгоду от мелких изменений конечностей в отдельных сегментах – в таком случае предпочтителен эмбриологический процесс с дополнительной тенденцией к выбросу одних только билатерально симметричных мутаций. И наконец, могут оказаться полезными мутации, эффект которых проявляется во всех сегментах тела одновременно, которые не отвергают уже существующие в сегментах и тагмах вариации, а усиливают их – например, перемножаясь.
Размышления о биологии подсказали нам с Тедом идею добавить в уже готовую программу артроморфов “переменный” ген. Благодаря ему то или иное свойство артроморфа – предположим, угол клешни – планомерно увеличивается или уменьшается по мере продвижения от головы к хвосту. У артроморфа с рис. 7.13е сегменты практически одинаковы, за исключением того, что их размер меняется с отрицательным градиентом. Тело сужается от головы к хвосту.
Размножение и селекция артроморфов идут по той же схеме, что и в программе для биоморфов. В центре экрана сидит артроморф-родитель, окруженный потомством со случайными мутациями. Как и с биоморфами, селекционер (человек) видит не гены, а только результат их влияния – форму тела, – и из разных форм отбирает пригодные для разведения (опять‐таки неполового). Выбранный артроморф перемещается в середину и окружает себя выводком собственных деток-мутантов. От поколения к поколению из‐за непредсказуемых мутаций меняются за кадром количество генов и их численные значения. Селекционер видит только ряд постепенно эволюционирующих артроморфов. Можно сказать, все артроморфы произошли от  , как и все биоморфы – от
, как и все биоморфы – от  . Сегменты тел аккуратно закрашены исключительно для красоты – в программе этой функции нет, хотя в дальнейшем, при трехуровневом генетическом контроле, ее нетрудно будет добавить. На рис. 7.14 показана коллекция артроморфов, подобная парку биоморфов (рис. 1.16); я периодически выводил “новые виды”, стараясь выдерживать стиль биологического реализма.
. Сегменты тел аккуратно закрашены исключительно для красоты – в программе этой функции нет, хотя в дальнейшем, при трехуровневом генетическом контроле, ее нетрудно будет добавить. На рис. 7.14 показана коллекция артроморфов, подобная парку биоморфов (рис. 1.16); я периодически выводил “новые виды”, стараясь выдерживать стиль биологического реализма.
, как и все биоморфы – от . Сегменты тел аккуратно закрашены исключительно для красоты – в программе этой функции нет, хотя в дальнейшем, при трехуровневом генетическом контроле, ее нетрудно будет добавить. На рис. 7.14 показана коллекция артроморфов, подобная парку биоморфов (рис. 1.16); я периодически выводил “новые виды”, стараясь выдерживать стиль биологического реализма.У представленных в моем зоопарке форм изменчивости подвержено все, что допускает калейдоскопическая эмбриология. Можно выделить группу, в которой наличие хотя бы одного переменного гена вызывает сужение тела. Можно различить четкое деление тел на тагмы – отделы последовательно расположенных сегментов, сходных между собой и явно отличающихся от других. Но даже в пределах одной тагмы можно различить вариации. В природе насекомые, ракообразные и пауки способны к таким же калейдоскопическим мутациям. Самый наглядный пример – гомеозис у членистоногих, когда в результате мутаций какой‐нибудь сегмент начинает развиваться по сценарию другого сегмента.

Рис. 7.14. Зоопарк артроморфов. Коллекция артроморфов, выведенных методами селекции и отобранных на глазок за внешнее сходство с природными членистоногими.
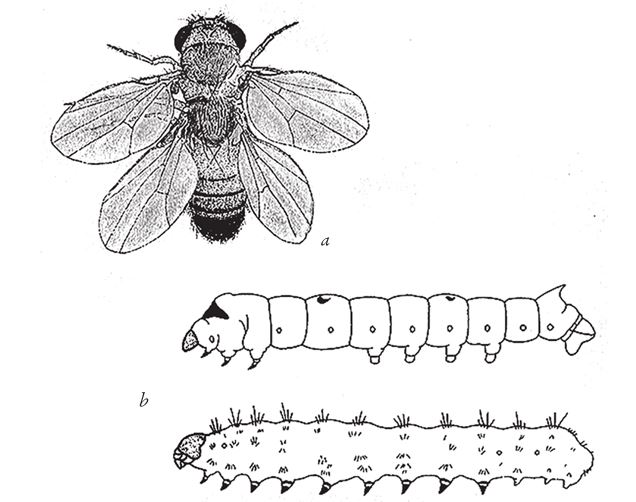
Рис. 7.15. Гомеозис: (а) дрозофила с четырьмя крылышками; в норме у дрозофилы вторую пару крыльев заменяют жужжальца, как на рис. 7.11; (b) типичная гусеница тутового шелкопряда (вверху) и гусеница, несущая мутацию (внизу). У нормальной гусеницы настоящие членистые конечности расположены только в трех грудных сегментах, у мутанта – девять “грудных” сегментов.
На рис. 7.15 показаны примеры гомеозиса у дрозофилы и гусеницы тутового шелкопряда. У нормальной дрозофилы, как и у всех мух, одна пара крылышек. Вместо второй – жужжальца (мы о них говорили). У изображенной на рисунке дрозофилы-мутанта вместо жужжалец выросла вторая пара крыльев, да вдобавок вместо третьего грудного сегмента дуплицировался второй грудной сегмент. Для артроморфов подобного эффекта можно добиться “грамматической” дупликацией с последующей делецией. На рис. 7.15b вы видите мутанта гусеницы шелкопряда. У всех нормальных гусениц, и шелкопряд не исключение, имеются три пары “настоящих”, составленных из члеников ног, хотя, как я уже говорил, в задних сегментах ноги мягкие, так сказать, “преобразованные”. Однако гусеница-мутант (рис. 7.15 внизу) крепко стоит на девяти настоящих ногах – как и правый артроморф с рис. 7.12. Самый известный случай гомеозиса – мутация antennapedia – был выявлен у дрозофилы. У мухи с мутацией гена antennapedia там, где должны были бы быть антенны, образовались обычные лапки. Механизм развития ног включился не в том сегменте, в каком положено.
Подобные мутанты выглядят форменными уродцами и вряд ли выживают в природе, что лишний раз подтверждает неприятие гомеозиса эволюцией. Именно поэтому я, проходя мимо стола с закусками на одном банкете в Австралии, невольно загляделся на блюдо с горой голенастых морепродуктов, хотя моя привередливость не позволила мне задержаться рядом с ним надолго. Мое внимание привлекла тварь, известная австралийским гурманам как глубоководный жук. Это разновидность омаров из группы, которую называют по‐разному – испанские омары, “тапочки” или лопатоносые лангусты. На рис. 7.16 изображен типичный представитель рода Scyllarus, экспонат Оксфордского университетского музея естественной истории. Меня поразило то, что у этого существа было явно два задних конца. Обман зрения был вызван тем, что передние антенны (строго говоря, вторая пара антенн) выглядели в точности как уроподы – конечности, характерные для заднего конца тела омаров. Почему антенны приобрели такую форму, я не знаю. Возможно, они служат лопатами или призваны обманывать хищников, как обманули меня. У омаров чрезвычайно развит быстрый сгибательный рефлекс, для этой цели предназначен гигантский нейрон. При малейшей угрозе они отскакивают назад как ошпаренные. Хищник мог бы предвидеть эту неприятность и подкрасться сзади. С обычным омаром военная хитрость удалась бы, но то, что вроде бы находится сзади по отношению к Scyllarus, запросто может оказаться перед ним, и тогда предусмотрительный хищник выберет самую неподходящую позицию для атаки. Не знаю, насколько справедливо такое предположение, но, по‐видимому, омарам их нетипичные усики приносят какую‐то пользу. Омары пускай устраиваются в жизни как хотят, я же, глядя на них, выдвину более смелую гипотезу. Мое мнение таково, что мы имеем дело с явлением гомеозиса в природе, аналогичным мутации типа antennapedia у лабораторных мушек-дрозофил. Однако здесь мутация была включена в естественный эволюционный процесс. Предположим, в древности сциллариды подверглись гомеозисной мутации, и подпрограмма развития уропод сместилась в те сегменты, где должны были бы сформироваться усики – и новая форма оказалась выгодной. Если я прав, тогда это подтверждает теорию “счастливых уродов”, о которой мы говорили в 3 главе, и мы наблюдаем тот самый редкий случай, когда естественный отбор “одобрил” макромутацию.
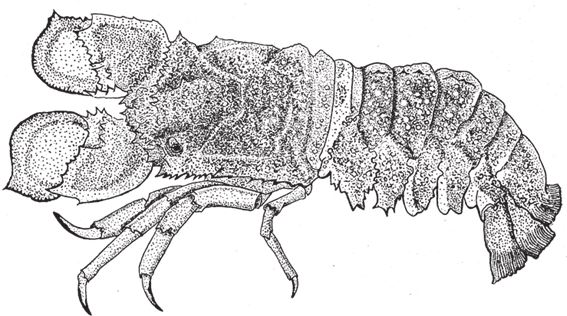
Рис. 7.16. Может, это удачный пример гомеозиса, реализовавшегося в ходе эволюции? Scyllarus, лопатоносый лангуст.
Однако это все спекулятивные рассуждения. Эмбриологи изучают гомеозисные мутации в лабораторных условиях и реконструируют детальную картину механизмов, обеспечивающих формирование сегментированного тела членистоногих в процессе эмбрионального развития. Их подробные отчеты чрезвычайно интересны, но мы не будем сейчас на них отвлекаться. В заключение я бы предложил читателю сопоставить некоторых настоящих членистоногих с компьютерными артроморфами и их калейдоскопическими мутациями.
Попробуйте представить себе, как членистоногие, изображенные на рис. 7.17, видоизменились бы под влиянием калейдоскопических “генов” артроморфов. Может быть, появятся особи с сужающимся телом, как на рис. 7.13е? Взгляните еще раз на подлинных членистоногих и вообразите какие‐то мелкие мутационные изменения, влияющие, например, на крайние членики конечностей или форму сегментированного туловища. Пусть для начала воображаемая мутация будет влиять на отдельно взятый сегмент. Готов поспорить, вы сразу подумали, что она должна зеркально отразиться с другой стороны, но это вовсе не обязательно. Теперь представьте себе мутацию, влияющую на кончики ножек, но в последовательно расположенных сегментах. На рис. 7.17 приведены примеры насекомых, у которых последовательные сегменты очень похожи. Третий вариант – мутация, влияющая на крайние членики конечностей всех сегментов – точнее, тех сегментов, где вообще есть конечности. Что касается меня, то, занимаясь артроморфами и их трехуровневой калейдоскопической эмбриологией, я стал по‐новому смотреть на живых членистоногих – таких, как на рис. 7.17. Более того, как и в случае с отражающими мутации зеркалами, легко догадаться, что эволюционный потенциал эмбрионального развития артроморфов с калейдоскопическими “ограничениями” на поверку может оказаться куда выше, чем при хаотическом, ничем не ограниченном процессе. В этом ключе особи, показанные на рис. 7.17, и все прочие не нарисованные здесь членистоногие обретают для меня особое значение.
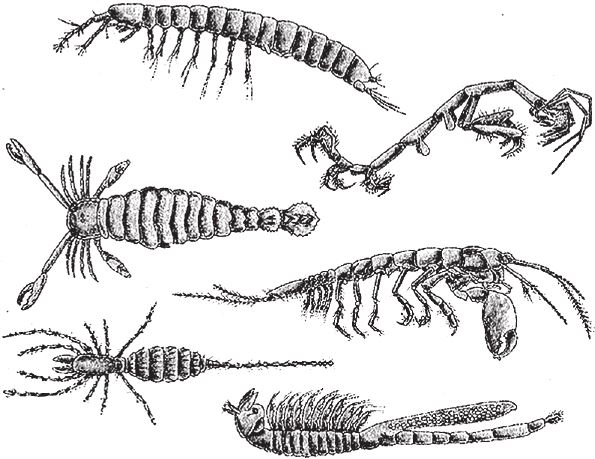
Рис. 7.17. Вот что может происходить с сегментами: представители членистоногих. По часовой стрелке, начиная сверху слева: четыре ракообразных, пальпиград (щупальцеходное, дальний родственник пауков и скорпионов) и ракоскорпион (эвриптерид, вымерший гигантский “морской скорпион”, его длина достигала почти трех метров).
В этой главе я хотел прежде всего донести до вашего сознания, что эмбриологический калейдоскоп парадоксальным образом служит как ограничивающим, так и стимулирующим фактором эволюции независимо от области его применения – в сегментах и линейных сегментных кластерах, выстроенных от головы до хвоста насекомого, или при зеркальном отображении у моллюсков. Ход эволюции ограничивается в том смысле, что калейдоскоп сужает спектр вариаций, с которыми мог бы работать естественный отбор. Эволюция стимулируется, так как естественный отбор (позволим себе несколько его очеловечить) не тратит времени на сканирование заведомо бесперспективных зон. Каждой из основных групп животных, которые населяют мир, – членистоногим, моллюскам, иглокожим, позвоночным – присущ свой, плодотворный с точки зрения эволюции, тип эмбрионального развития с калейдоскопическими ограничениями. У калейдоскопической эмбриологии есть все для того, чтобы унаследовать землю. Всякий раз, когда калейдоскопический режим, или “зеркало”, становился причиной удачного всплеска форм в эволюции, все генеалогические линии в новом пуле видов наследовали этот режим и это зеркало. Это уже не обычный дарвиновский естественный отбор, а его разновидность нового, более высокого уровня. И вывод о том, что вследствие такого отбора произошла эволюция с повышением способности эволюционировать, уже не кажется игрой разгоряченного ума.

