Книга: Мозг. Такой ли он особенный?
Назад: Что означает разница в правилах нейронного шкалирования
Дальше: Что значит быть приматом
Преимущество приматов
В мозговой коре животных, не принадлежащих к отряду приматов, по мере роста числа нейронов плотность равномерно уменьшается, и это уменьшение можно описать степенной функцией от числа нейронов с аллометрическим показателем функции –0,6 (рис. 4.13). Поскольку средний размер нейрона изменяется обратно пропорционально плотности нейронов, этот показатель степени подразумевает, что средняя масса корковых нейронов возрастает с увеличением числа нейронов, возведенного в степень +0,6. Это означает, что если кора приобретает в десять раз больше нейронов, то размер каждого нейрона в среднем увеличивается в четыре раза, а масса всей коры – в сорок раз. Если же кора приобретает в сто раз больше нейронов, то нейроны становятся в 16 раз больше, а масса коры – в 1600 раз; при приобретении 1000 нейронов они становятся в 64 раза больше, а масса коры увеличивается в 64 тысячи раз. Напротив, у приматов, по мере увеличения числа корковых нейронов, плотность их упаковки в коре практически не изменяется (рис. 4.13). Это означает, что по мере развития коры приматов она начинает приобретать дополнительные нейроны без существенного увеличения их среднего размера. Это не означает, что нейроны приматов становятся мельче, но лишь то, что они перестают расти: размер клеток в коре приматов остается таким же, как размер нейронов в коре крыс или кроликов, и значительно мельче, чем у агути. Вследствие этого, когда кора приматов приобретает в десять раз больше нейронов, ее масса становится всего в десять раз больше, а если она приобретает в 100 раз больше нейронов, то и масса ее увеличивается тоже в 100 раз.
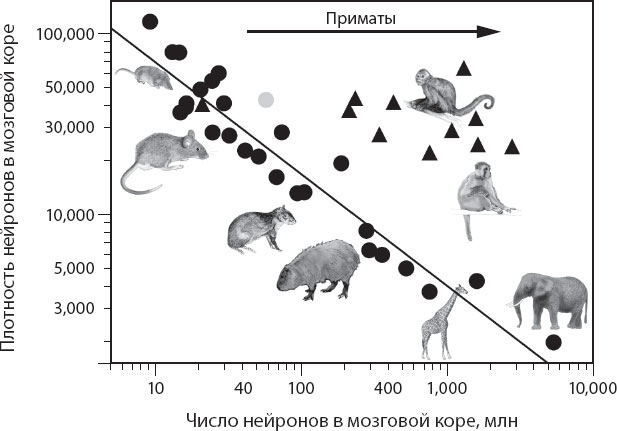
Рис. 4.13. По мере того как кора головного мозга животных, не принадлежащих к отряду приматов (кружки), приобретает нейроны, плотность их (в нейронах на 1 мг мозговой коры) уменьшается пропорционально увеличению числа нейронов, возведенного в степень –0,6, что означает следующее: масса нейронов возрастает в зависимости от увеличения числа нейронов, возведенного в степень +0,6. Напротив, по мере того как кора приматов (треугольники) приобретает нейроны, плотность нейронов снижается очень незначительно. Учитывая, что эволюция млекопитающих началась с животных малого размера с маленькими мозгами и крошечной корой, можно считать, что приматы отделились от общего предка с изменением, которое устранило связь между добавлением новых нейронов и увеличением средних размеров нейронов в коре головного мозга
Отчетливая тесная корреляция между плотностью упаковки нейронов (а следовательно, средним их размером) и числом нейронов у неприматов позволяет предположить, что должен существовать механизм, благодаря которому в эволюции этих млекопитающих произошло сопряжение увеличения числа нейронов в мозговой коре с увеличением среднего размера этих нейронов. Каждый раз, когда пока не открытый механизм увеличивал число нейронов, образующих мозговую кору нового биологического вида, этот механизм приводил и к увеличению среднего размера нейронов в фиксированном отношении. Более того, этот механизм был (и продолжает оставаться) весьма устойчивым, ибо он работает уже более 90 миллионов лет, то есть до расхождения африканских зверей, грызунов, насекомоядных и парнокопытных, и, на самом деле, до сих пор присутствует в современных видах каждой из этих таксономических групп. Когда приматы, в свою очередь, отделились от своего общего с неприматами предка, случилась поломка: упомянутый механизм перестал работать и произошло разобщение увеличения числа корковых нейронов и увеличения их среднего размера. То есть, на сколько бы ни увеличивалось число корковых нейронов при переходе от одного примата к следующему, не происходило одновременного увеличения среднего их размера. Таким образом, правила нейронного шкалирования у приматов стали другими, специфичными для этого отряда животных.
Поскольку первые, самые ранние виды приматов были мельче нынешних, этот «разрыв» в происхождении приматов проявился у животных с малым числом нейронов в их мозговой коре, и это число подчинялось правилам нейронного шкалирования, характерным как для приматов, так и для неприматов, то есть этот разрыв был локализован на пересечении распределения приматов и неприматов, как показано на рис. 4.8 и 4.13. Ископаемые остатки родоначальника приматов Ignacius graybullianus позволяют оценить его внутричерепной объем в 2,14 см3, а массу тела в 231 г. Если этот вид действительно был близок к точке ответвления приматов и если древние правила нейронного шкалирования для предковых форм млекопитающих были еще применимы, то мы можем считать массу мозговой коры равной 0,96 г, что дает число нейронов, равное 42,4 миллиона, что близко к числу нейронов, определенному для современного мышевидного лемура. Между тем мышевидный лемур находится на эволюционной лестнице там, где можно ожидать появления предкового примата на наших графиках, – на пересечении правил нейронного шкалирования мозговой коры приматов и неприматов. Применив правила нейронного шкалирования, которые, как мы считаем, были характерны для предкового млекопитающего, мы получили промежуточный результат – 33,9 миллиона нейронов в коре головного мозга. Близость двух оценок – это как раз то, чего следовало ожидать на основании предполагаемого сценария ответвления приматов от общего предка, то есть свойство, носителем которого был этот предок, общее с современными неприматами, но которое все более модифицировалось с изменением правил нейронного шкалирования, каковое становилось все более заметным по мере эволюционного увеличения числа нейронов в мозговой коре приматов.
Что может означать тот факт, что кора головного мозга приматов приобретает нейроны, которые, как правило, не становятся крупнее, но становятся более многочисленными в противоположность другим млекопитающим, для которых из увеличения числа нейронов обязательно следует увеличение массы отдельно взятого нейрона? Первое и самое главное – это значит, что природа не ограничена одним-единственным способом построения коры мозга. Но, во-вторых, в этом заключается важное преимущество приматов, состоящее в том, что нейроны не становятся крупнее по мере роста их числа: кора увеличивается в объеме лишь настолько, насколько больше она приобретает нейронов, и не более того. Что касается объема, добавление нейронов, которые лишь очень незначительно увеличиваются в размерах, является весьма экономичным способом добавления нейронов к коре в сравнении с альтернативой, имеющей место у неприматов, кора которых намного больше при равном числе нейронов. Это важно, потому что объем играет очень большую, если не главную роль: например, чем больше кора, тем больше времени требуется для того, чтобы потенциал действия распространился по волокнам коры, что замедляет обработку и интеграцию информации, поступившей в кору. В самом деле, по нашим оценкам, среднее время распространения сигнала в белом веществе коры быстро увеличивается у грызунов, по закону степенной функции от числа корковых нейронов, возведенного в степень +0,466, в то время как у приматов время распространения сигнала нарастает куда медленнее, так как кора приматов приобретает новые нейроны при аллометрическом показателе степени +0,165. Характерный для приматов способ нейронной организации коры дает им явное преимущество, так как укорачивает время распространения сигнала по мере приобретения корой новых нейронов (а следовательно, увеличения объема коры) в сравнении с неприматами.
Здесь, однако, возникает сложная проблема: даже если кора приматов становится лишь прямо пропорционально больше по мере приобретения нейронов, то расстояние между двумя точками коры тоже неизбежно увеличивается, а, значит, эти точки должны быть соединены между собой более крупными или более длинными нейронами, тем не менее нейроны сохраняют свой средний размер. Как это может быть верным одновременно? Ответ заключается в ключевой концепции среднего размера нейронов: это означает, что некоторые нейроны могут быть крупнее (в данном случае – длиннее), но если это происходит, то за счет того, что другие становятся короче. В отношении коры и связанного с ней белого вещества это означает, что если некоторые корковые нейроны в более крупном мозге становятся крупнее, обладая более длинными волокнами, проходящими через ткань белого вещества, то в тем большей степени остальные нейроны ограничивают местоположение своих волокон серым веществом. Если средний размер нейронов не изменяется или изменяется очень незначительно среди видов приматов, то нейроны первого типа (образующие дальние связи с подкорковыми структурами через белое вещество) должны увеличиваться в числе медленнее, чем нейроны второго типа (образующие ближние связи внутри серого вещества, не выходя за его пределы в ткань белого вещества). В самом деле, проведенное нами отдельное изучение того, как подкорковое белое вещество увеличивается в размерах по мере приобретения корой дополнительных нейронов, показало, что корковые пути в белое вещество, то есть доля корковых нейронов, связанных с длинными волокнами, уходящими в белое вещество, на самом деле уменьшаются по мере увеличения числа нейронов, как это и предсказано согласно приведенному выше сценарию. Именно так укрупняются локальные сети: не путем роста всех связей, а добавлением множества новых местных связей и лишь небольшого числа ключевых длинных связей. Напротив, связи коры с белым веществом у грызунов остаются стабильными по мере того, как кора этих животных приобретает новые нейроны, и эти нейроны становятся в среднем крупнее.
Что касается мозжечка, то здесь мы видим, что плотность упаковки нейронов у неприматов и у животных, не принадлежащих также к отряду насекомоядных, уменьшается по мере того, как мозжечок приобретает новые нейроны, как это показано на рис. 4.14, что указывает на то, что средний размер нейронов увеличивается совместно с увеличением числа мозжечковых нейронов у этих видов, в соответствии с законом степенной зависимости с показателем функции +0,3. Это означает, что десятикратное увеличение числа нейронов, образующих мозжечок животных, не принадлежащих к отрядам приматов и насекомоядных, сопровождается двукратным увеличением среднего размера нейронов; тысячекратное увеличение числа этих нейронов сопровождается восьмикратным увеличением среднего их размера. В результате мозжечок приматов, независимо от размера, имеет плотность упаковки нейронов – а следовательно, средний размер нейронов, – сравнимую с этим показателем в мозжечке крысы.
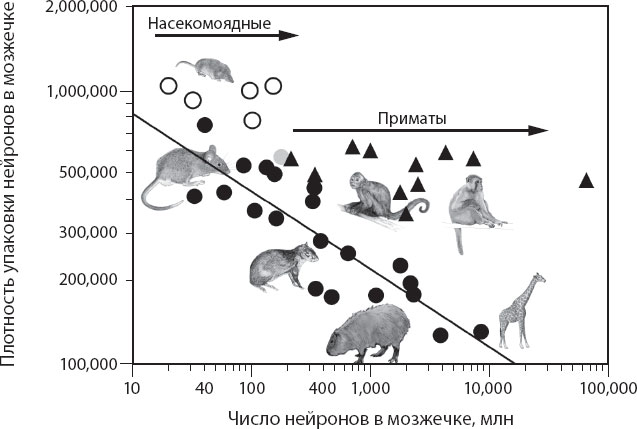
Рис. 4.14. По мере того как мозжечок животных, не принадлежащих к отрядам приматов и насекомоядных (черные кружки), получает дополнительные нейроны, плотность их упаковки падает пропорционально числу нейронов, возведенному в степень –0,3, а это означает, что средняя масса нейронов возрастает пропорционально числу нейронов, возведенному в степень +0,3. Напротив, по мере того как мозжечок получает новые нейроны у приматов (треугольники) и у насекомоядных (белые кружки), не наблюдается отчетливого уменьшения плотности упаковки нейронов. Учитывая, что эволюция млекопитающих началась с животных, обладавших малым мозгом и, соответственно, малой корой, можно считать, что приматы и насекомоядные ответвились от соответствующих им предков с изменением, которое устранило сопряжение между увеличением числа мозжечковых нейронов и увеличением их среднего размера
Примечательно, что увеличение среднего размера нейронов происходит в мозжечке значительно медленнее, чем в коре головного мозга, при равном приобретении новых нейронов этими структурами у животных, не принадлежащих к отряду приматов. Это несовпадение приобретает смысл, если сравнить анатомию этих двух структур. Замедление увеличения среднего размера нейронов в мозжечке по сравнению с корой, вероятно, обусловлено разницей строения длинных связей между этими двумя структурами. В то время как мозговая кора обладает большим числом длинных реципрокных связей между участками коры, проходящих через подкорковое белое вещество, белое вещество мозжечка состоит исключительно из входных и выходных связей коры мозжечка, но отсутствуют связи, соединяющие разные участки этой коры. В мозжечке нет ничего похожего на мозолистое тело, соединяющее два полушария. Самые длинные волокна нейронов мозжечка достигают в длину всего нескольких миллиметров (в противоположность сантиметрам в коре большого мозга), и эти волокна проходят в верхнем слое коры самого мозжечка параллельно листкам мозжечка, где эти волокна переносят информацию между клетками Пуркинье. Организация волокон в мозжечке, таким образом, геометрически более экономична, чем в коре большого мозга, и именно это является основой многих различий в локальных функциях этих двух структур. В то время как длинные волокна, проходящие через белое вещество полушарий большого мозга, позволяют колонкам нейронов коры мозга обрабатывать информацию, приходящую из участков мозга, отстоящих от колонок на расстояние в несколько сантиметров, и направлять ее дальше в расходящихся, сходящихся или реципрокных направлениях, более многочисленные параллельные волокна верхнего слоя мозжечковой коры (одно волокно приходится на каждую зернистую клетку) передают информацию в латеральном направлении на расстояние всего в несколько миллиметров до ближайших клеток Пуркинье, где происходит обработка и анализ информации.
Насекомоядные и приматы, как представляется, независимо друг от друга отошли от соответствующих предковых правил нейронного шкалирования и приобрели изменение, согласно которому произошло разобщение увеличения числа нейронов и увеличения среднего их размера, что наглядно показано на рис. 4.14. Это изменение подразумевает, что в ходе эволюции насекомоядных и приматов их мозговые структуры стали приобретать дополнительные нейроны без дальнейшего значимого увеличения их среднего размера. Отсутствие увеличения среднего размера нейронов в мозжечке насекомоядных и приматов позволяет предположить, что параллельные волокна в верхнем слое коры мозжечка передавали сигналы латерально на те же самые или меньшие расстояния у крупных и мелких приматов и насекомоядных, если клетки Пуркинье в более крупном мозжечке не становились меньше, в каковом случае параллельные волокна начали бы передавать сигналы на более длинные расстояния. Это последнее едва ли правдоподобно, однако; у некоторых видов млекопитающих, у которых были определены размеры клеток Пуркинье, происходит их увеличение по мере возрастания объема головного мозга. Тем не менее самая важная находка состоит в том, что насекомоядные и приматы получают нейроны в мозжечок без увеличения среднего размера нейронов, а это указывает на то, что не существует единого пути построения более крупного мозжечка, как не существует и единственного пути построения более крупной коры.

