Исследование № 5
Происхождение митоза, мейоза и полового размножения
До сих пор мы говорили об эволюции бесполых популяций, в которых нет горизонтального обмена генетической информацией между организмами. В природе, однако, такой обмен в той или иной форме распространен очень широко, а самых впечатляющих успехов добились организмы, практикующие на удивление сложный (и порой весьма затратный) его вариант, известный под названием “половое размножение”. Прежде чем обсуждать, зачем оно нужно (один из любимых вопросов эволюционистов-теоретиков, который им никогда не надоедает), хорошо бы понять, как вообще мог возникнуть данный тип межорганизменной рекомбинации (перемешивания генетического материала разных особей). Познакомимся с одной из гипотез. Она связывает происхождение полового размножения с полиплоидными прокариотами (археями), обитавшими в мелководных микробных сообществах в начале протерозойского эона. В то время уровень свободного кислорода уже начал расти, но озонового экрана еще не образовалось, так что темп мутагенеза должен был резко повыситься. Моделирование показывает, что в таких условиях полиплоидность у прокариот, не имеющих митоза, дает кратковременное эволюционное преимущество, но в долгосрочной перспективе повышает риск вымирания из-за накопления рецессивных вредных мутаций. Полиплоидные микробы могли справляться с этой проблемой несколькими способами, причем все они подозрительно напоминают те или иные аспекты или этапы полового размножения. Их постепенное совершенствование и комбинирование логически приводит к появлению сначала митоза, а затем мейоза и полового размножения. Гипотеза объясняет, как и почему вместе с половым размножением должны были возникнуть и некоторые другие специфические особенности эукариот: множественные линейные хромосомы, высокий уровень генетической избыточности и быстрое появление новых генных семейств на заре эволюции эукариот. К числу фактов, согласующихся с гипотезой, относится и недавно обнаруженная корреляция между полиплоидностью и наличием гистонов у архей.


Происхождение эукариот и половое размножение. Появление эукариот – второе по значимости событие в истории земной жизни (первое – само возникновение жизни). Эукариотическая клетка устроена гораздо сложнее, чем прокариотическая, а промежуточные формы между ними, существовавшие когда-то, вымерли. Поэтому вопрос о происхождении эукариот остается одним из самых сложных и спорных в эволюционной биологии. Правда, недавно ситуация резко изменилась к лучшему благодаря открытию асгардархей – неизвестной ранее большой группы архей, которая по многим признакам является промежуточной между обычными археями и эукариотами. Но даже с учетом асгардархей получается, что эволюционный путь от последнего общего предка эукариот и их ближайших архейных родственников до последнего общего предка всех современных эукариот (LECA, last eukaryotic common ancestor) был долгим и трудным. На этом пути предки эукариот приобрели целый ряд признаков, которых нет ни у кого из прокариот, даже у асгардархей. Одним из последних шагов на этом пути стало, по-видимому, приобретение внутриклеточных симбионтов – альфа-протеобактерий, давших начало митохондриям.
К числу эукариотических инноваций, наиболее трудных для объяснения, относится эукариотический секс (называемый также амфимиксисом, или попросту половым размножением). Это специфический и очень эффективный способ перемешивания генетического материала разных особей, включающий сингамию (слияние гаплоидных клеток или ядер, приводящее к удвоению хромосомного набора) и мейоз (особый вариант клеточного деления, приводящий к сокращению числа хромосом вдвое). Мейоз сопровождается кроссинговером, в ходе которого гомологичные хромосомы обмениваются гомологичными участками.
Прокариотический секс (горизонтальный перенос генов, ГПГ) устроен гораздо проще. Можно назвать три основных отличия эукариотического секса (полового размножения) от прокариотического (ГПГ).
Во-первых, у эукариот обмен генами взаимный, двусторонний: каждый участник является и донором, и реципиентом. У прокариот передача генетического материала строго однонаправленная: один участник – донор ДНК, другой – реципиент.
Во-вторых, для эукариот характерна полногеномная рекомбинация: в обмене участвуют два полных генома. У прокариот в рекомбинации участвует один полный геном и один фрагмент.
В-третьих, у эукариот все гены, участвующие в рекомбинации, сохраняют шанс перейти к потомкам. У прокариот фрагмент ДНК донора либо встраивается в геном реципиента вдобавок к уже имеющимся там генам (но этот вариант не может использоваться регулярно, например в каждом поколении, ведь это привело бы к безудержному разрастанию генома), либо замещает собой гомологичный фрагмент генома реципиента. При этом замещенные аллели реципиента уничтожаются и теряют шанс перейти к потомкам (рис. 5.1).
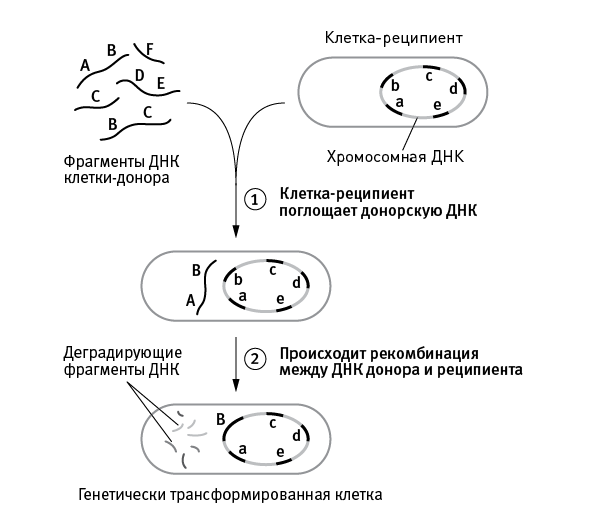
рис. 5.1. Один из способов горизонтального переноса генов у прокариот (естественная трансформация, сопряженная с гомологичной рекомбинацией), наиболее близкий по своим эволюционным последствиям к эукариотическому сексу. Бактерия поглощает фрагмент чужой ДНК из окружающей среды (это могут быть, например, фрагменты геномов погибших бактерий того же вида), а затем встраивает его в свою кольцевую хромосому вместо собственного гомологичного фрагмента. В результате аллели бактерии-реципиента уничтожаются, замещаясь чужими аллелями. В данном случае аллель B встроился в хромосому и будет передан потомкам, а аллель b погиб.
Раньше многие специалисты предполагали, что первые эукариоты были бесполыми (размножались клонально – простым делением – и не имели полового процесса), а потом у кого-то из них появилось половое размножение. В таком случае проблема возникновения эукариотического секса не связана напрямую с проблемой происхождения эукариот. Однако по мере накопления геномных данных стало ясно, что эукариотический секс является одной из фундаментальных особенностей эукариот, которая почти наверняка имелась уже у LECA. В частности, появляется все больше аргументов в пользу того, что все современные бесполые группы эукариот произошли от предков, имевших мейоз (в их геномах сохранились остатки необходимых для мейоза генов), а значит, и половой процесс. Стало быть, происхождение эукариот и происхождение полового размножения – проблемы взаимосвязанные.
Половой процесс солелюбивой археи Haloferax. Если эукариотический секс – исходный признак эукариот, то имеет смысл поискать среди современных архей варианты генетического обмена, переходные между типичным прокариотическим и эукариотическим сексом. Крайне интересно было бы выяснить, как меняются генами асгардархеи (переходная группа между обычными археями и эукариотами), но об этом пока ничего не известно. На сегодняшний день подробно изучена система генетического обмена только у одной археи – солелюбивого (галофильного) микроба Haloferax volcanii, обитателя Мертвого моря. У Haloferax обнаружена система передачи генетического материала, действительно похожая на что-то промежуточное между обычным прокариотическим ГПГ (например, конъюгацией бактерий) и эукариотическим сексом.
Клетки Haloferax спариваются, соединяясь цитоплазматическими мостиками, по которым геномная ДНК может передаваться в обе стороны, то есть обе клетки могут быть как донорами, так и реципиентами генетического материала. При этом могут передаваться и крупные фрагменты хромосомы (длиной до 13 % генома как минимум). В лаборатории можно добиться даже слияния клеток: для этого понижают концентрацию ионов Mg2+ в среде, что приводит к растворению клеточных стенок и дестабилизации цитоплазматических мостиков. Предполагается, что такое слияние иногда происходит и в природе.
В 2010 году Гросс и Бхаттачарья предложили красивую гипотезу, связывающую воедино происхождение эукариот и полового размножения и опирающуюся в том числе на данные по половому процессу Haloferax (Gross, Bhattacharya, 2010). В ней обосновываются три положения:
1) Первые эукариоты появились в раннепротерозойских мелководных местообитаниях, когда концентрация кислорода начала расти, но озонового экрана еще не было. Под действием ультрафиолета на мелководьях должна была повыситься концентрация активных форм кислорода – сильных мутагенов. Появление эукариот стало закономерным итогом попыток архей защититься от этой напасти.
2) Архейные предки эукариот обменивались генами примерно так же, как современные представители рода Haloferax. Гросс и Бхаттачарья выстроили гипотетический сценарий постепенного эволюционного перехода от полового процесса Haloferax к полноценному эукариотическому сексу.
3) Совершенствование системы генетического обмена было необходимо предкам эукариот, поскольку они использовали чужую ДНК в качестве матрицы для починки разрывов двойной спирали своей ДНК при помощи гомологичной рекомбинации (примерно так, как это делает микроб Deinococcus radiodurans, способный выдерживать колоссальные дозы радиации).
При всех достоинствах этой гипотезы она не учитывает одну важную деталь, которая не противоречит первым двум пунктам, но сильно бьет по третьему. Дело в том, что Haloferax, как и многие другие археи, – полиплоиды, то есть имеют в каждой клетке множество (в среднем 17) копий генома – кольцевых хромосом.
Полиплоидность архей и ее эволюционные следствия. Все привыкли думать, что у прокариот в клетке только одна кольцевая хромосома. У многих бактерий и архей это действительно так. По умолчанию считалось, что Haloferax тоже моноплоиды. Однако исследования последних лет показали, что полиплоидность широко распространена у бактерий и архей. В частности, полиплоидами являются, по-видимому, все галоархеи (группа солелюбивых архей, к которым относится род Haloferax), метаногены и термоплазмы. Полиплоидной клетке не нужна чужая ДНК, чтобы чинить разрывы при помощи гомологичной рекомбинации: у нее для этого достаточно собственных хромосом.
Полиплоидность может быть полезна прокариотам по целому ряду причин, включая упомянутую репарацию разрывов ДНК, отсроченное фенотипическое проявление вредных рецессивных мутаций и повышенную надежность синтеза белка в неблагоприятных условиях. Помимо прочего, “лишняя” геномная ДНК служит запасом пищи на черный день (полиплоидные археи попросту съедают часть своих хромосом, когда им не хватает фосфора).
Полиплоидность Haloferax заставляет по-новому взглянуть на природу факторов отбора, которые привели к появлению эукариотического секса. Причем все становится даже интереснее, чем в исходной модели Гросса и Бхаттачарьи.
В 2016 году Александр Марков и Илья Казначеев при помощи компьютерного моделирования попытались выяснить, что будет происходить с популяцией полиплоидных архей, если она попадет в сильно мутагенную среду (то есть в условия, провоцирующие ускоренный мутагенез). Выяснилось, что эволюционно-генетические последствия полиплоидности у прокариот, обитающих в мутагенной среде, шаг за шагом логически приводят к появлению полового размножения (Markov, Kaznacheev, 2016).
Для начала нужно пояснить, что у полиплоидных архей нет митоза. Напомним, что при митозе дочерние клетки получают ровно по одной копии каждой родительской хромосомы. У полиплоидных архей при делении родительской клетки хромосомы распределяются между потомками примерно поровну, поэтому уровень плоидности остается более или менее постоянным, но то, какие именно хромосомы достанутся каждому потомку, по-видимому, никак не регулируется и определяется случайностью (рис. 5.2). Из этого вытекают интересные эволюционные следствия. В частности, копии гена, расположенные на разных хромосомах, не могут поделить между собой функции, как это часто происходит у имеющих митоз эукариот после полиплоидизации (полногеномной дупликации). Ведь если нет митоза, то нет и никакой гарантии, что каждый потомок получит оба специализированных варианта гена.
Разработанная компьютерная модель имитировала эволюцию популяции одноклеточных организмов с различной плоидностью. У каждой клетки предполагалось определенное число исходно одинаковых кольцевых хромосом, каждая хромосома содержала заданное число локусов (генов). Каждый ген характеризовался “качеством работы”, которое могло ухудшаться из-за вредных мутаций и улучшаться благодаря полезным. От качества работы генов зависела конкурентоспособность клетки и ее шансы на выживание в каждом поколении. В модели можно было регулировать скорость мутагенеза, соотношение вредных и полезных мутаций, численность популяции, способ распределения хромосом по дочерним клеткам при делении и многое другое. Кроме того, модель позволяла имитировать разные варианты генетического обмена и гомологичной рекомбинации как внутри клетки, так и между клетками. Для простоты все вредные мутации считались рецессивными, и поэтому для вычисления приспособленности полиплоидной клетки использовалась “лучшая” из копий каждого гена. Это справедливо для многих мутаций, снижающих функциональность кодируемого белка. Таким образом, проведенный анализ относится только к тем генам, одной исправной копии которых достаточно для выживания клетки.
Моделирование показало, что при высокой скорости мутагенеза моноплоиды в долгосрочной перспективе более жизнеспособны, чем полиплоиды. Однако поначалу (первые несколько сотен поколений) полиплоиды опережают моноплоидов по приспособленности (рис. 5.3).
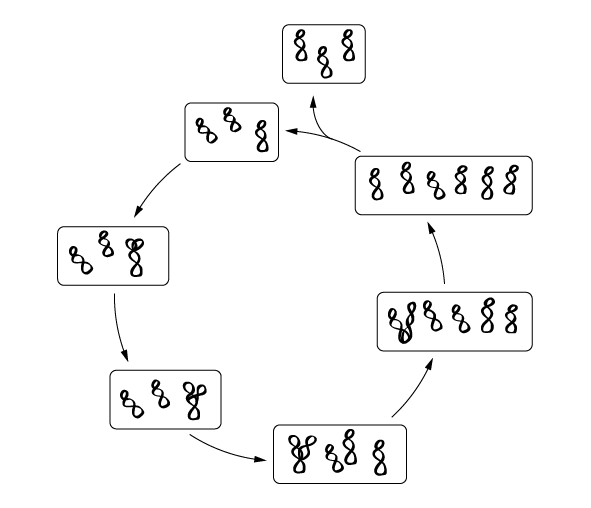
рис. 5.2. Схема жизненного цикла полиплоидной (в данном случае – триплоидной) бактерии, не имеющей митоза. Три кольцевые хромосомы, каждая из которых содержит весь геном и может отличаться от двух других отдельными мутациями, реплицируются (удваиваются) в произвольном порядке, пока число хромосом не станет равно шести. После этого клетка делится на две дочерние триплоидные клетки. Поскольку митоз отсутствует, каждой из дочерних клеток достаются три случайно выбранные хромосомы из шести (при митозе каждой дочерней клетке досталось бы ровно по одной копии каждой из трех родительских хромосом).
Причин кратковременного преимущества полиплоидов две. Первая – отложенное проявление в фенотипе вредных рецессивных мутаций. Такая мутация у полиплоидов не дает о себе знать, пока не начнут появляться клетки, несущие ее в каждой копии генома (гомозиготные). Вторая причина – ускоренное накопление редких доминантных полезных мутаций. При фиксированной частоте их возникновения в расчете на локус вероятность приобретения клеткой полезной мутации прямо пропорциональна плоидности.
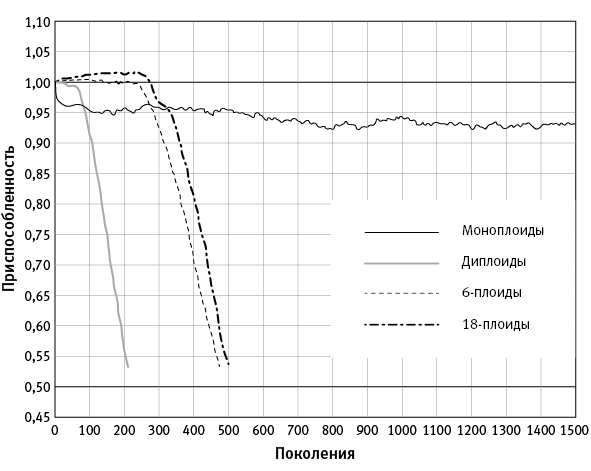
рис. 5.3. Результат компьютерного моделирования, показывающий эволюционную судьбу популяций одноклеточных организмов с разной плоидностью при высокой скорости мутагенеза. Митоза нет, генетического обмена и рекомбинации тоже нет. По вертикальной оси – средняя приспособленность особей в популяции. Моноплоиды выходят на равновесный уровень приспособленности (при котором порча генофонда за счет мутагенеза уравновешивается его улучшением за счет отбора) и существуют неопределенно долго. Полиплоиды сначала имеют сильное преимущество, но затем быстро деградируют и вымирают. Из статьи Markov, Kaznacheev, 2016.
Причины последующего вырождения полиплоидов – слабый очищающий отбор, быстрое накопление рецессивных вредных мутаций и увеличение сегрегационного груза. Это значит, что в какой-то момент жизнеспособные полиплоидные клетки начинают производить нежизнеспособных потомков. Это неизбежное следствие накопления рецессивных вредных мутаций при отсутствии митоза. Термин “сегрегационный груз” обычно применяют к эукариотам в ситуации, когда гетерозиготы имеют более высокую приспособленность, чем гомозиготы (сегрегация – это разделение, в данном случае имеется в виду распределение родительских хромосом по геномам потомков). При скрещивании двух гетерозигот часть потомков оказываются гомозиготами и имеют пониженную приспособленность – это и называют сегрегационным грузом. В случае полиплоидных архей имеется в виду следующее. Представим себе триплоидную клетку, у которой из трех жизненно необходимых генов A, B и C первый исправен только на первой хромосоме, второй исправен на первой и третьей хромосомах, третий – только на второй. Такая клетка, сама по себе вполне жизнеспособная, может, если ей не повезет, поделиться на двух нежизнеспособных потомков:
У первого потомка нет ни одной работающей копии гена C, а второму не досталось рабочих копий гена A. Заметим, что при наличии митоза этой проблемы не было бы: оба потомка имели бы точно такой же генотип, как у родительской клетки, и жили бы припеваючи.
Поскольку полиплоидность приводит к вырождению не сразу, а сначала дает сильное преимущество, она может стать своеобразной “эволюционной ловушкой” для прокариот в мутагенной среде. Если разрешить модельным микробам иногда при делении распределять хромосомы не поровну, то есть менять свою плоидность, то полиплоиды сначала быстро вытесняют моноплоидов, а затем сами деградируют и вымирают. Это происходит даже в том случае, если подавляющее большинство клеток в исходной популяции – облигатные моноплоиды и лишь немногие клетки способны иногда производить потомков, имеющих на одну хромосому больше. Полиплоидность распространяется как инфекция – и приводит популяцию к гибели. При тех же параметрах популяция, состоящая только из облигатных моноплоидов, может жить неопределенно долго.
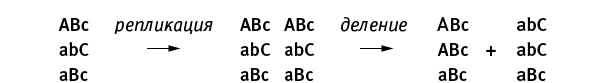
Четыре способа защиты полиплоидов от вырождения. Чтобы выжить, такие полиплоиды должны выработать специальные адаптации, замедляющие накопление вредных мутаций. Они могут использовать для этого как минимум четыре разные стратегии. Вот тут-то и начинается самое интересное. Дело в том, что все эти четыре способа защиты полиплоидных прокариот от вырождения подозрительно напоминают те или иные аспекты эукариотического секса.
Первый способ – “циклы плоидности”. Можно периодически сбрасывать уровень плоидности, например, делясь чаще, чем происходит репликация хромосом. Если заниматься этим достаточно интенсивно, то в популяции будет постоянно присутствовать (или периодически возникать) заметная доля моноплоидов, у которых все вредные рецессивные мутации проявляются в фенотипе и потому эффективно вычищаются отбором.
Второй способ – внутригеномная рекомбинация, то есть перетасовка генетической информации между хромосомами. Есть два основных варианта такой рекомбинации. Первый вариант называется генной конверсией. В этом случае фрагмент одной хромосомы копируется в гомологичный участок другой, причем аллели, находящиеся на второй хромосоме, “затираются” аллелями первой. Интенсивная генная конверсия ведет к унификации копий генома. Любая новая мутация либо быстро затирается и исчезает, либо распространяется на все хромосомы, переходит в гомозиготное состояние и становится видимой для отбора. Моделирование показывает, что генная конверсия, если ее интенсивность существенно превышает темп мутагенеза, может спасти полиплоидную популяцию от вырождения.
Полиплоидные галофильные и метаногенные археи активно используют генную конверсию, предположительно, именно для того, чтобы унифицировать свои хромосомы и тем самым защититься от вырождения. Эту стратегию, вероятно, с той же целью применяют и пластиды растений (они тоже полиплоидные и не имеют митоза).
Второй вариант внутригеномной рекомбинации – кроссинговер. В этом случае гомологичные участки двух хромосом не затирают друг друга, а меняются местами. Кроссинговер не может ни уничтожить вредную мутацию, ни перевести ее в гомозиготное состояние. Поэтому сам по себе он бесполезен для полиплоидных микробов, но в сочетании с ГПГ дает сильный положительный эффект (см. ниже).
Третий способ – интенсивный генетический обмен (ГПГ) между близкородственными клетками. Моделирование показывает, что горизонтальный перенос генов (такой, как на рис. 5.1) хорошо защищает полиплоидов от вырождения, особенно если осуществляется с высокой частотой. Полиплоидные археи действительно меняются друг с другом генами на полную катушку. Эволюционный эффект интенсивного близкородственного ГПГ в целом такой же, как у эукариотического секса (мы рассказывали об этом в книге “Эволюция. Классические идеи в свете новых открытий”). Интересно, что кроссинговер, бесполезный для популяций, не практикующих ГПГ, оказывается очень полезен для микробов, периодически заимствующих гены друг у друга.
Польза, приносимая ГПГ, имеет ту же природу, что и польза, приносимая половым размножением: оба процесса позволяют отбору отделять полезные аллели от вредных, эффективно закрепляя первые и выбраковывая вторые (см. Исследование № 7). При этом ГПГ тем полезнее, чем чаще он происходит. При высоком темпе мутирования это особенно актуально. Однако у прокариотического ГПГ есть встроенный конструктивный дефект, не позволяющий этому процессу достигать оптимальной (то есть высокой) частоты. Дефект кроется в асимметричности (“нечестности”) прокариотического ГПГ, которую хорошо иллюстрирует рис. 5.1. На рисунке видно, что чужой (донорский) аллель B заместил и уничтожил аллель b в геноме реципиента. Ситуация, когда свои аллели систематически замещаются чужими, может оказаться эволюционно нестабильной. Чтобы понять это, нужно подумать о судьбе генов, влияющих на интенсивность (частоту) захвата чужой ДНК и замещения собственных аллелей чужими. Допустим, у такого гена есть два аллеля: один способствует ГПГ, другой препятствует. Какой из них победит в конкуренции? Моделирование показывает, что аллели, препятствующие ГПГ, могут распространяться в генофонде и вытеснять аллели, способствующие ГПГ, несмотря на всю пользу, которую получают от ГПГ отдельные организмы и популяция в целом. Ведь аллели, способствующие ГПГ, будут то и дело “затираться” конкурирующими аллелями, которые ГПГ блокируют. А вот в обратную сторону замещение происходить не будет – аллели, блокирующие захват чужой ДНК и замещение фрагментов своей хромосомы чужими, не будут затираться как раз потому, что они блокируют ГПГ. В результате аллели, препятствующие ГПГ, будут вести себя как “эгоистичные гены”, наращивая свою частоту в генофонде, – несмотря на то, что это вредно для особей и популяции в целом.
Могут ли полиплоидные микробы обойти это препятствие, чтобы получить возможность осуществлять межорганизменный генетический обмен с высокой частотой? По-видимому, да. Для этого им нужно, во-первых, начать обмениваться не кусочками хромосом, а целыми хромосомами, и во-вторых – отказаться от асимметричной генной конверсии, исключить “затирание” одних аллелей другими и использовать для перемешивания фрагментов хромосом только кроссинговер. Умеют ли полиплоидные археи меняться целыми хромосомами, точно не известно, но это представляется вполне вероятным, исходя из того, что известно о половом процессе у Haloferax.
С кроссинговером, правда, возникает еще одна проблема: кольцевые хромосомы плохо для него подходят. При нечетном числе перекрестов они не могут нормально разойтись после рекомбинации и превращаются в одно большое кольцо. Поэтому, если вы хотите часто использовать кроссинговер, вам нужно отказаться от кольцевых хромосом и заменить их линейными. Идея о том, что линейные хромосомы понадобились эукариотам именно для частого кроссинговера, а не для чего-то еще, уже высказывалась ранее рядом специалистов, и с ней трудно спорить, учитывая, что во всех прочих отношениях кольцевые хромосомы удобнее.
Четвертый способ, помогающий полиплоидным амитотическим микробам защититься от вырождения, – самый радикальный. Он состоит в том, чтобы изобрести митоз – механизм аккуратного и точного распределения хромосом по дочерним клеткам, гарантирующий, что каждый потомок получит ровно по одной копии каждой родительской хромосомы. Это моментально снимает все проблемы, связанные с накоплением сегрегационного груза.
После изобретения митоза все выгоды частого обмена хромосомами и кроссинговера сохраняются в полной мере. Поэтому у полиплоидных архей, научившихся аккуратно распределять хромосомы по дочерним клеткам, не было оснований отказываться от хромосомного обмена. Но со временем это закономерно привело к новому конфликту, для разрешения которого пришлось изобрести мейоз.
Изобретение митоза приводит к диверсификации хромосом, что порождает новые проблемы, для решения которых нужно изобрести мейоз. Неизбежным следствием изобретения митоза полиплоидными археями – предками эукариот – должна была стать быстрая диверсификация хромосом. Митоз снимает проблему сегрегационного груза, и поэтому избыточные копии генов, расположенные на разных хромосомах, получают небывалую эволюционную свободу. В рассматриваемой компьютерной модели они просто начинают деградировать, беспрепятственно накапливая вредные мутации, так что скоро у каждого гена остается только одна неиспорченная копия, расположенная на любой из хромосом. Каждая хромосома при этом становится уникальной и незаменимой, потому что те гены, которые остались на ней неиспорченными, безнадежно испорчены на всех остальных хромосомах.

В модели не предусмотрено приобретение генами новых функций. Однако это именно то, что наверняка будет происходить в подобной ситуации у реальных живых организмов. Многие избыточные гены будут потеряны или безнадежно испорчены прежде, чем в них возникнет полезная мутация, но многие поделят функции или выработают новые.
Таким образом, изобретение митоза фактически превращает полиплоидный организм в моноплоидный, обладающий несколькими разными хромосомами, с высоким уровнем генетической избыточности. Здесь уместно вспомнить, что множественные хромосомы и генетическая избыточность – характерные эукариотические черты, происхождение которых долго оставалось загадкой. Предложенная гипотеза дает им внятное историческое объяснение.
По мере того как хромосомы специализируются и становятся уникальными, унаследованные от предков способы свободного хромосомного обмена и рекомбинации становятся все менее выгодными, а затем и вовсе начинают вредить. Действительно, если все ваши хромосомы уникальны и незаменимы, вы уже не можете просто передать в другую клетку по цитоплазматическому мостику одну-две случайно выбранные хромосомы и получить какие-то другие взамен. Необходимо сделать обмен хромосомами и рекомбинацию более избирательными, чтобы в них участвовали только очень похожие (гомологичные) хромосомы. Моделирование подтверждает эти рассуждения. Кроме того, хорошо бы еще позаботиться о том, чтобы каждая хромосома принимала участие в рекомбинации с оптимальной частотой.
Очевидное решение проблемы состоит в развитии клеточного слияния (которое, возможно, уже имелось у предков эукариот, ведь его наличие предполагается у Haloferax) и спаривания гомологичных хромосом двух клеток с кроссинговером и последующим аккуратным распределением по дочерним клеткам. Важно, чтобы в ходе клеточного деления, следующего за попарной рекомбинацией хромосом, каждая дочерняя клетка получила строго по одной хромосоме из каждой гомологичной пары. Уже имеющийся механизм митоза служит идеальной заготовкой (преадаптацией) для эволюции такого клеточного деления. И вот они – сингамия и мейоз.
Изучение молекулярных механизмов мейоза уже давно привело специалистов к выводу, что мейоз наверняка развился на основе митоза. Также есть веские аргументы в пользу того, что эволюция мейоза началась с развития механизма спаривания гомологичных хромосом и что смысл этого новшества был не в том, чтобы повысить интенсивность рекомбинации, а в том, чтобы ограничить ее, запретив рекомбинацию между непохожими хромосомами (Wilkins, Holliday, 2009). Так что результаты моделирования хорошо согласуются с этой идеей и объясняют, откуда взялась потребность в такой адаптации.
Предлагаемый эволюционный сценарий.
1) Предки эукариот были полиплоидными археями, не имевшими митоза. Они жили в мутагенной среде на мелководьях во времена “кислородной революции”, когда на планете впервые стал расти уровень свободного кислорода. В таких условиях быть полиплоидом полезно в краткосрочной перспективе, но чревато вымиранием в долгосрочной.
2) Отбор способствовал выработке адаптаций, уменьшающих негативные эффекты полиплоидности при сохранении ее преимуществ. В результате выработался набор средств, снижающих риск вырождения полиплоидов в мутагенной среде: унификация хромосом путем генной конверсии, интенсивный горизонтальный перенос генов между родственниками (спаривание с образованием цитоплазматических мостиков, возможно – временное слияние клеток), циклы плоидности (периодические редукционные деления).
3) Поскольку эти относительно простые полумеры, по-видимому, не решили проблему полностью, в дальнейшем развивались еще более эффективные средства защиты от генетической деградации: обмен целыми хромосомами в сочетании с рекомбинацией, переход от конверсии к кроссинговеру (поскольку он эволюционно стабилен и позволяет осуществлять генетический обмен часто), замена кольцевых хромосом линейными.
4) Наконец, был изобретен митоз – аккуратное распределение хромосом при делении, так что каждая дочерняя клетка стала гарантированно получать ровно одну копию каждой родительской хромосомы. Это сразу сняло проблему сегрегационного груза. Однако обмен хромосомами в сочетании с кроссинговером по-прежнему был весьма полезен, так что у предков эукариот не было оснований от него отказываться.
5) Специализация и диверсификация хромосом, являющиеся неизбежным следствием изобретения митоза полиплоидами, постепенно вступили в конфликт со старыми способами безвыборочного генетического обмена и рекомбинации. Эти способы “устарели”, и отбор способствовал их модернизации. В результате развились механизмы, обеспечивающие обмен, спаривание и рекомбинацию только очень похожих (гомологичных) хромосом. В конечном счете это привело к развитию сингамии и мейоза, то есть настоящего эукариотического полового размножения.
6) Одновременно должно было происходить совершенствование механизмов выбора брачного партнера, поскольку, когда вы подходите к межорганизменной рекомбинации столь серьезно, спариваться с кем попало опасно. Подобно тому как хромосомы стали спариваться для обмена участками только с очень похожими хромосомами, клетки должны были начать спариваться только с клетками, имеющими такой же хромосомный набор. Результат – появление “биологических видов” с хорошо перемешиваемыми и в меру изолированными генофондами. Но пока механизмы выбора партнеров были еще несовершенны, эукариоты могли нахватать много генов от неродственных линий (что они, судя по всему, и сделали).
Испытание на прочность. Всякая гипотеза, чтобы получить признание, должна пройти проверку временем и новыми фактами. Это дело будущего. Впрочем, несколько испытаний гипотеза Маркова и Казначеева уже благополучно прошла.
Во-первых, в 2015 году, когда шла работа над моделью, появилось сообщение о том, что у архей обнаружена строгая корреляция между полиплоидностью и наличием гистонов (Spaans et al., 2015). То, что у некоторых архей есть гистоны, было известно давно, и этот факт всегда считался важным аргументом в пользу того, что предками эукариот были именно археи. Новые данные показывают, что этот аргумент приложим только к полиплоидным археям. Связь между гистонами и полиплоидностью, по-видимому, объясняется тем, что гистоны помогают упаковывать множество копий генома в одной маленькой прокариотической клетке. Плоидность ближайших прокариотических родственников эукариот – асгардархей – напрямую не измерялась, поскольку эти микробы известны только по геномным последовательностям. Однако в геномах представителей всех четырех типов асгардархей имеются гены гистонов. Значит, асгардархеи, скорее всего, полиплоиды.
Во-вторых, из гипотезы вытекает одно очень специфическое проверяемое следствие. Если эукариоты произошли от полиплоидных архей, которые приобрели митоз, это обязательно должно было привести к массовому появлению новых семейств близкородственных генов у базальных (примитивных) эукариот. Подобный процесс, как мы знаем, происходит у эукариот в результате полногеномных дупликаций. Если в геноме появляется множество “избыточных” копий генов, какие-то из них наверняка приобретут новые функции – и тогда вместо одного гена получится целое семейство похожих, но все же различающихся генов – так называемых паралогов. И что замечательно, сравнительная геномика подтверждает это предсказание: еще в 2005 году Евгений Кунин и его коллеги показали, что на заре эволюции эукариот был период массового приобретения новых семейств паралогичных генов (Makarova et al., 2005).
Так что пока все сходится, и эукариоты, возможно, действительно произошли от полиплоидных архей, которые изобрели митоз. Хотя не исключено, что будущие исследователи предложат еще более интересный и обоснованный сценарий.

