Исследование № 4
Ранние этапы адаптации предсказуемы, поздние – случайны
Эксперимент Ленски показал, что в бесполых популяциях даже в неизменных условиях идет непрерывный рост приспособленности. Происходит это за счет накопления и закрепления полезных мутаций. Хорошо бы разобраться подробнее в этом процессе: что за мутации, как и в какой последовательности они распространяются в популяции. Эту непростую задачу удалось решить с помощью новой методики “генетического штрихкодирования”. Применив ее, американские ученые смогли в небывалых подробностях изучить процесс накопления полезных мутаций в большой бесполой популяции дрожжей при адаптации к новой среде. Как выяснилось, на начальных этапах общий рост приспособленности популяции идет за счет высоковероятных мутаций со слабым положительным эффектом, которые возникают независимо у множества особей. На этой стадии процесс адаптации предсказуем: его можно описать простыми формулами. В дальнейшем роль случайности возрастает, потому что на первый план выходят маловероятные мутации с сильным полезным эффектом. Кроме того, исследование наглядно показало, что темп появления полезных мутаций может быть весьма высоким.


В популяциях, насчитывающих миллионы особей, в каждом поколении возникает множество новых мутаций – и вредных, и полезных, и нейтральных (напомним, что категория мутации определяется ее влиянием на приспособленность, то есть на эффективность передачи особью своих генов следующим поколениям). Все эти мутации вносят вклад в среднюю приспособленность особей, от которой зависит скорость роста численности популяции. Возникновение новых мутаций и изменение частоты их встречаемости под действием отбора и генетического дрейфа – самые фундаментальные эволюционные процессы. Нельзя понять эволюцию, не изучив их во всех подробностях.
Но как уследить за тысячами мутаций, происходящих у миллионов особей? Секвенировать целиком миллионы геномов – неподъемная задача даже при современном уровне развития биотехнологий. Если же применять выборочное секвенирование, то в поле зрения исследователей попадут только те мутации, которые достигли высокой частоты встречаемости (например, как в Исследовании № 3). Картина получится весьма неполной. Ведь многие возникающие полезные мутации, вероятно, никогда не становятся массовыми, однако свой вклад в общую приспособленность тем не менее вносят.
Альтернативный подход состоит в том, чтобы пометить отдельные клоны (клетки, произошедшие от одной и той же родительской клетки) наследуемой генетической меткой, а потом следить, как меняется численность каждого из них. Если численность какого-то клона вдруг начала экспоненциально расти, в то время как число всех особей популяции остается постоянным, значит, у одного из представителей этого клона возникла полезная мутация. При этом скорость роста является мерой полезности мутации. Например, если рост численности клона описывается уравнением N = N0 × (1 + 0,05)t, где время t измеряется в поколениях, значит, мутация повысила приспособленность на 5 % (в таких случаях говорят, что полезность мутации, обозначаемая буквой s, равна 0,05).
Именно такое маркирование и осуществили американские биологи, продемонстрировав настоящий прорыв в технике наблюдений за эволюцией многомиллионных популяций (Levy et al., 2015). Ученые работали с двумя бесполыми популяциями дрожжей (их искусственно лишили способности к половому размножению, так что они размножались только почкованием) численностью по 108 клеток. Популяции были произведены от одной-единственной предковой клетки, то есть изначально геномы всех дрожжей были одинаковыми. В каждой популяции были помечены индивидуальными генетическими метками примерно по 500 000 клонов. Как это удалось сделать? Сначала изготовили большую коллекцию кольцевых молекул ДНК – плазмид, – содержащих случайные двадцатинуклеотидные последовательности (генетический “штрихкод”). Эти плазмиды внедрялись в дрожжевые клетки, геномы которых были предварительно модифицированы таким образом, чтобы плазмиды встраивались в строго определенное место генома при помощи особого фермента – Cre-рекомбиназы. В итоге удалось получить две популяции численностью по 108 клеток, в которых каждая клетка принадлежала к одному из полумиллиона помеченных клонов.
Затем в течение 168 поколений обе популяции адаптировались к “голодной” среде, где размножение ограничивалось количеством глюкозы (как и в эксперименте Ленски). Численность каждого клона отслеживалась путем массового секвенирования небольшого фрагмента генома, содержащего “штрихкод”. Секвенировать приходилось лишь 0,002 % генома, что позволило резко увеличить разрешающую способность метода по сравнению с полногеномным секвенированием. В поле зрения исследователей попали даже те мутации, частота встречаемости которых в популяции никогда не превышала 10–5, тогда как секвенирование полных геномов позволило бы отследить лишь клоны с относительной численностью 10–2 и выше. В результате вместо 25 000 зарегистрированных мутаций исследователи сумели бы обнаружить лишь около 15 (для сравнения вспомним, что в Исследовании № 3 удалось проследить судьбу только тех мутаций, чья частота встречаемости достигала 10 %, то есть 10–1, или более).
Впрочем, даже зная численность каждого клона в разные моменты времени, определить, в каком из них возникла полезная мутация, – не такая простая задача (рис. 4.1). Каждая мутация возникает сначала у одной особи. Пока число потомков удачного мутанта невелико, динамика их численности определяется не столько приспособленностью (и следовательно, отбором), сколько случайными колебаниями (дрейфом). Большая часть вновь возникающих полезных мутаций теряется из-за дрейфа: потомки удачного мутанта просто не успевают достичь такой численности, при которой отбор “заметит” их полезное свойство и начнет его поддерживать. Мутация становится заметна для отбора (и выходит из-под власти дрейфа) лишь по достижении численности мутантов, сопоставимой с 1/s. Например, мутация с полезностью 0,01 (повышающая приспособленность на 1 %) становится заметна для отбора при числе мутантов около 100. Мутантному клону должно повезти, чтобы его численность случайно (то есть за счет дрейфа) доросла до этого порога, – и лишь тогда за дело возьмется отбор. Такую мутацию называют “установившейся”. Это значит, что она уже не потеряется из-за дрейфа.
После того как мутация “установилась”, численность клона будет экспоненциально расти. По скорости роста можно оценить полезность мутации (s), а экстраполяцией в прошлое примерно определить время возникновения мутации (τ). Впрочем, этот удобный для количественного анализа период в истории клона будет продолжаться лишь до тех пор, пока в геноме, уже содержащем одну полезную мутацию, не возникнет вторая. После этого все расчеты резко затрудняются и картина смазывается. Именно поэтому данная методика позволяет анализировать лишь начальные этапы адаптации – пока вероятность возникновения дополнительных полезных мутаций пренебрежимо мала (а она перестает быть таковой, когда численность клона приближается к 1/Ub, где Ub – частота возникновения полезной мутации в расчете на особь за поколение).
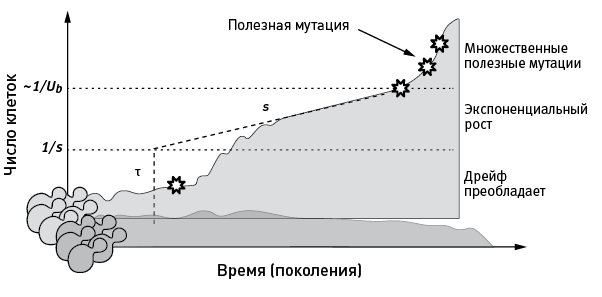
рис. 4.1. Типичная судьба клонов бесполых организмов в ходе адаптации. Темно-серым цветом показана динамика численности клона дрожжей, в котором не возникло полезных мутаций. Такой “нейтральный” клон, скорее всего, не достигнет высокой численности и будет вытеснен более успешными конкурентами. Светло-серым цветом показана судьба клона, в котором в момент времени τ возникла полезная мутация (изображена звездочкой). Пока численность такого клона мала, он может вымереть просто случайно (из-за дрейфа). Чтобы отбор начал поддерживать носителей полезной мутации, клон должен достичь (опять-таки случайно) пороговой численности, которая для мутации с полезным эффектом s примерно равна 1/s. После прохождения этого порога клон с полезной мутацией вступает в фазу экспоненциального роста численности. Когда численность клона приблизится к 1/Ub, где Ub – частота возникновения полезных мутаций, в нем с большой вероятностью начнут появляться дополнительные полезные мутации. Шкала по вертикальной оси логарифмическая. По рисунку из Levy et al., 2015.
После того как был получен гигантский массив данных по скоростям роста численности отдельных клонов (а мы помним, что их полмиллиона в каждой из двух подопытных популяций), в дело пошла статистика и компьютерное моделирование. Выводы получились следующие.
Численность большинства клонов с течением времени снижалась, нередко до полного исчезновения клона. Такая судьба характерна для “нейтральных” клонов, в которых не возникло полезных мутаций. Однако примерно у 5 % клонов за первую сотню поколений появилась и “установилась” хотя бы одна полезная мутация.
По мере размножения клеток с полезными мутациями средняя приспособленность популяции росла. Это, в свою очередь, приводило к тому, что “нейтральные” клоны, в которых не было полезных мутаций, оказывались во все более проигрышном положении. Их численность продолжала убывать. Более того, многие “удачливые” клоны, поначалу бодро размножавшиеся благодаря рано возникшей слабополезной мутации, в дальнейшем приходили в упадок и вымирали. Это объясняется тем, что на фоне возросшей приспособленности конкурентов их полезная мутация уже не давала преимущества.
Средняя приспособленность популяций за 100 поколений выросла на 5 %. Соответственно, к этому времени все обладатели мутаций с s < 0,05 оказались в проигрыше, хотя на ранних этапах именно эти многочисленные клоны вносили основной вклад в общий рост приспособленности.
Эти результаты говорят о том, что ключевым процессом, определяющим эволюционную динамику большой бесполой популяции, является безжалостная конкуренция между клонами – обладателями разных полезных мутаций (клональная интерференция). При этом ранними стадиями адаптации заправляют высоковероятные и потому многочисленные слабополезные мутации. Таких мутаций, повышающих приспособленность на 2–5 % (0,02 < s < 0,05, мутации с более слабым эффектом остались незамеченными), в первой из двух подопытных популяций установилось около 20 000, во второй – 11 000. На этом этапе ход адаптации в значительной мере предсказуем: его можно описать уравнениями и при повторных эволюционных экспериментах результаты получаются схожими. Затем, по мере роста средней приспособленности, многочисленные слабополезные мутации фактически перестают быть полезными и начинают отбраковываться. Продолжает нарастать лишь численность тех немногих клонов, в которых возникли редкие мутации с сильным положительным эффектом. К 70-му поколению динамика общей приспособленности определялась в основном мутациями с полезностью около 0,1. Таких мутаций в двух подопытных популяциях было 5000 и 3000 соответственно. После 110-го поколения на первый план вышли примерно по 100 клонов с наиболее полезными мутациями (s > 0,1). На этом этапе ход адаптации становится все менее предсказуемым, поскольку то, когда и в каком количестве возникнут эти маловероятные мутации, зависит от случая.
Внимательный читатель может заметить кажущееся противоречие между этими результатами и теми, которые были получены в эксперименте Ричарда Ленски (см. Исследование № 1). Там сначала закреплялись мутации с сильным положительным эффектом, а лишь затем начинали фиксироваться слабополезные. На самом деле расхождений тут нет, просто речь идет о разном временном масштабе и о разных этапах адаптации: у Ленски – десятки тысяч поколений, в рассматриваемой работе – 168. Иными словами, в данном исследовании внимание фокусируется на том начальном этапе, который Ленски пролетел на полном ходу. Разнятся также отслеживаемые параметры и разрешающая способность методов. В работе Ленски регистрировались уже зафиксировавшиеся, то есть достигшие стопроцентной частоты, полезные мутации, а здесь говорится о мутациях, только начавших свой путь. Подавляющее большинство из них никогда не зафиксируется, а будет вытеснено в ходе клональной интерференции. Если бы эксперимент продлился еще несколько тысяч поколений, то зафиксировались бы как раз немногочисленные мутации с самым сильным положительным эффектом, точно как в эксперименте Ленски.
Исследование раскрыло тонкие детали начальных этапов адаптации. В частности, оно объяснило, почему многочисленные слабополезные мутации поначалу не имеют шанса зафиксироваться: они неизбежно отбраковываются по мере роста общей приспособленности популяции.
Исследование также наглядно показало, что полезные мутации – вовсе не редкость. После того как дрожжи были помещены в “голодные” условия, полезные мутации с s > 0,05 (то есть очень полезные) возникали с частотой одна мутация на 1 000 000 клеток за поколение. Базовая частота мутирования (возникновения любых мутаций) составляла одну мутацию на 200 клеток за поколение. Получается, что очень полезной оказывалась в среднем одна мутация из каждых 5000. Чуть менее полезных мутаций (0,02 < s < 0,05) возникало на порядок больше: умеренный полезный эффект давала каждая пятисотая мутация. Это очень много! И ведь это только мутации, которым повезло быть подхваченными отбором, без учета тех, что сразу потерялись из-за дрейфа.
Впрочем, частоты появления полезных мутаций зависят от условий, к которым нужно приспосабливаться. Мутации случайны, но вероятность того, что та или иная случайная мутация окажется полезной, зависит от обстановки. Понятно, что, если поместить дрожжи в идеальные для них условия, вероятность того, что случайная мутация окажется полезной, будет крайне низка. Когда все и так прекрасно, трудно что-то улучшить (и очень легко испортить). Соответственно, в плохих условиях вероятность того, что случайная мутация произведет положительный эффект, будет выше. Этот общий закон жизни иногда называют “принципом Анны Карениной”, поскольку первая фраза романа Льва Толстого наглядно его иллюстрирует: существует много разных способов все испортить, но очень мало способов достичь совершенства.
Таким образом, начальный рост общей приспособленности бесполой популяции обусловлен теми слабополезными мутациями, которым не суждено достичь высоких частот и которые вскоре будут вытеснены. Таких мутаций оказалось неожиданно много. Однако бесполая популяция бездарно растрачивает почти все это “генетическое богатство” из-за клональной интерференции. Настоящая гонка начинается уже после того, как слабые игроки вышли игры.

