Исследование № 11
Гордые красавцы против невзрачных проныр
В нагрузку к половому размножению и половому отбору раздельнополые организмы получили и так называемый конфликт полов. Он возникает из-за того, что оптимальные стратегии поведения самцов и самок, как правило, различаются. Самцы, производящие много “дешевых”, маленьких сперматозоидов, могут максимизировать свой репродуктивный успех путем оплодотворения максимального числа партнерш. Самки, вкладывающие больше ресурсов в каждого потомка, заинтересованы не столько в количестве, сколько в “качестве” партнеров. Это различие “эволюционных интересов” и создает почву для конфликта полов, который проявляется в развитии признаков, выгодных одному полу, но вредных другому. Между самцами и самками может даже начаться эволюционная гонка вооружений, почти как между хищниками и жертвами. Конфликт полов развивается вопреки тому, что любой ген попадает попеременно то в женский, то в мужской организм и поэтому должен, по идее, “заботиться” в равной мере о тех и других. Ничего личного: естественный отбор (включая и такую его разновидность, как половой отбор) автоматически поддерживает аллели, у которых сумма положительных эффектов хоть немного превышает сумму отрицательных. То, какие формы примет конфликт полов в каждом конкретном случае, зависит от множества факторов, в том числе от того, в какой момент самка вкладывает ресурсы в потомство – до оплодотворения (как у животных, чьи эмбрионы развиваются за счет запасов желтка в яйце) или после (как у плацентарных). Теория предсказывает, что в первом случае будут развиваться самцовые турниры за самок и броские брачные наряды у будущих отцов, а самки будут становиться до крайности привередливыми и разборчивыми. Во втором случае открываются иные возможности. Самкам может стать выгодно спариваться с несколькими партнерами и вынашивать одновременно потомство от разных отцов. Это позволяет отбирать уже не женихов, а зародыши. Арена действия полового отбора как бы перемещается внутрь женского организма: теперь самка может отбирать наилучшие эмбрионы в ходе внутриутробного развития, вкладывая больше ресурсов в самые быстрорастущие и жизнеспособные. В такой ситуации брачные наряды самцов, скорее всего, потускнеют за ненадобностью, брачные турниры и танцы сойдут на нет и самцы начнут искать другие, менее “рыцарские” пути максимизации своего репродуктивного успеха. Но постойте, не слишком ли разыгралась фантазия эволюционистов-теоретиков? Бывает ли такое в природе? На примере живородящих рыбок семейства пецилиевых, у которых характер материнского вклада в потомство в ходе эволюции многократно менялся, американским биологам удалось показать, что эти теории – не вымысел кабинетных ученых, а работающая модель, хорошо объясняющая некоторые причудливые зигзаги эволюции.


В 2000 году биологи из Университета Невады в Рино (США) Дэвид и Джин Це предложили гипотезу “конфликта, связанного с живорождением” (Zeh, Zeh, 2000). Суть идеи в следующем. Если самка вкладывает свои ресурсы в потомство еще до оплодотворения (презиготически) – в виде запасов желтка в яйце, – то ей выгодно тщательно выбирать полового партнера до спаривания, чтобы доверить свои дорогостоящие яйцеклетки кандидату с наилучшими генами. Следовательно, у самок должна развиваться презиготическая половая избирательность, а у самцов – разнообразные средства соблазнения привередливых самок, в том числе яркие наряды и сложные ритуалы ухаживания.
Если же большую часть питательных веществ эмбрион получает от матери уже после оплодотворения (постзиготически), например, через специальный орган – плаценту, то ситуация будет иной. Яйца теперь можно сделать маленькими и “дешевыми”, а выбор партнера – постзиготическим. Самка получает возможность вкладывать в эмбрион больше или меньше своих ресурсов в зависимости от его свойств, обусловленных работой отцовских генов. Простейший вариант – лучше “кормить” те эмбрионы, которые быстро растут. Это позволяет самке оптимальнее распределить свои ресурсы, потому что скорость развития эмбриона и его биохимические и иммунологические свойства могут нести более адекватную информацию о генах отца, чем яркие перья и причудливые танцы.
Чтобы реализовать это преимущество, самке нужно спариваться не с одним, а со многими самцами (такое поведение биологи называют полиандрией) и вынашивать одновременно потомство, зачатое в разное время (это называют суперфетацией). При этом роль презиготического выбора, женской привередливости, брачных нарядов и ритуалов должна снижаться. Теперь у самцов вместо украшений будут развиваться адаптации, направленные на то, чтобы обеспечить спаривание с максимальным числом партнерш вне зависимости от эстетических предпочтений невест. Образно говоря, из гордых красавцев самцы начнут превращаться в невзрачных проныр.
Переход от лецитотрофного (обеспечиваемого желтком) развития к плацентарности имеет еще одно важное следствие. Плацентарность создает арену для генетических конфликтов и эволюционной гонки вооружений (антагонистической коэволюции) между матерью и эмбрионом, а также между материнскими и отцовскими генами эмбриона. Дело в том, что в интересах эмбриона – добыть как можно больше ресурсов из материнского организма, тогда как в интересах матери – частично обуздать эти притязания, дабы сохранить здоровье и силы для рождения других детей.
Из гипотезы Дэвида и Джин Це вытекает несколько проверяемых следствий. Например, у плацентарных организмов из-за постоянной гонки вооружений (в ходе которой и матери, и эмбрионы вырабатывают все новые и новые средства нападения и защиты) должна быстрее развиваться генетическая несовместимость между популяциями. Если в геноме гибридного потомка встретятся “сильные” аллели, играющие на стороне матери, и “слабые” аллели, играющие на стороне зародыша, последний погибнет от голода, тогда как противоположное сочетание плохо кончится для матери. Из этого следует, что у плацентарных между разделившимися видами должна быстрее развиваться постзиготическая репродуктивная изоляция (пониженная приспособленность, бесплодие или полная нежизнеспособность гибридов). И действительно, как показали сами супруги Це в той же статье, у птиц и лягушек виды могут сохранять частичную совместимость (способность производить живое гибридное потомство) десятки миллионов лет после своего расхождения, тогда как у млекопитающих полная нескрещиваемость развивается на порядок быстрее (рис. 11.1). Этот факт хорошо согласуется с гипотезой Це, хотя у него, в принципе, могут быть и другие объяснения.

рис. 11.1. Диаграмма, показывающая, что у лягушек и птиц полная постзиготическая репродуктивная изоляция (неспособность производить живое гибридное потомство) развивается медленнее, чем у млекопитающих. По горизонтальной оси – время с момента разделения пары видов (в млн лет), по вертикальной – число пар, способных к гибридизации. Учитывались только виды, способные к межвидовой гибридизации. По рисунку из Zeh, Zeh, 2000.
Предсказания гипотезы Це также получили ряд эмпирических подтверждений (Coleman et al., 2009). Среди них работа американского эволюциониста Дэвида Резника и его коллег, опубликованная в 2014 году, выглядит наиболее показательной (Pollux et al., 2014). Ученые проанализировали распределение признаков у 94 видов живородящих рыб семейства пецилиевых. К этому семейству относятся такие известные аквариумные рыбки, как меченосцы и моллинезии. Пецилиевые – прекрасный объект для подобных исследований, потому что в ходе эволюции у рыб из этого семейства несколько раз независимо развивалась и утрачивалась плацентарность (рис. 11.2). Более того, в пределах семейства представлен полный спектр репродуктивных стратегий: от полного отсутствия постзиготического материнского вклада (рост эмбриона только за счет желтка) до развитой плацентарности, со всеми промежуточными стадиями (Reznick et al., 2002).
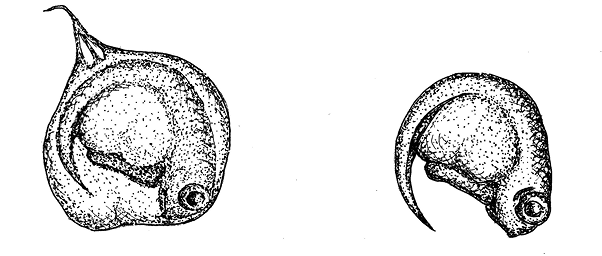
рис. 11.2. Эмбрионы плацентарной рыбки Poeciliopsis presidionis. Слева – эмбрион, окруженный плацентой; справа – плацента удалена.
Величину постзиготического материнского вклада оценивали по отношению массы новорожденного малька к массе яйцеклетки в момент оплодотворения (индексу матротрофии, MI). У лецитотрофных видов, чье внутриутробное развитие идет за счет желтка, новорожденный малек весит меньше оплодотворенной икринки: MI < 1 (обычно от 0,5 до 0,75). У плацентарных видов MI варьирует примерно от 1 (если малек получает лишь незначительную поддержку от матери во время беременности) до 100 и более.
Резник и его коллеги построили эволюционное дерево пецилиевых по последовательностям двадцати ядерных и восьми митохондриальных генов и сопоставили его с данными по MI. Анализ подтвердил, что в ходе эволюции пецилиевых постзиготический материнский вклад в разных линиях то рос, то уменьшался. Плацентарность была развита у последнего общего предка пецилиевых, а затем многократно независимо усиливалась или слабела и вовсе терялась.
Сопоставление MI с признаками действия полового отбора на самцов подтвердило предсказания гипотезы Це. Использовались три характеристики, указывающие на выбор самками самцов по внешнему виду и поведению: 1) половой диморфизм по окраске (когда самцы окрашены иначе, чем самки), 2) сложное брачное поведение и брачные демонстрации, 3) наличие у самцов украшений, таких как гипертрофированные спинные плавники, “усы” на верхней губе или “меч” на хвостовом плавнике, как у меченосцев. Оказалось, что все три показателя отрицательно коррелируют с MI. Иными словами, чем больше питательных веществ получают эмбрионы от матери через плаценту, тем слабее выражены у самцов специфическая “мужская” окраска, сложное брачное поведение и гротескные украшения.
Выяснилось также, что суперфетация (одновременное вынашивание детенышей, зачатых в разное время) у пецилиевых, напротив, положительно коррелирует с MI. Если самка способна к суперфетации, значит, она может одновременно вынашивать потомство от нескольких отцов. В таком случае самец не может монополизировать репродуктивный потенциал самки, оплодотворив сразу все ее яйцеклетки, а у самки появляется возможность осуществлять постзиготический выбор оптимальных отцовских генотипов в ходе внутриутробного развития. Роль презиготического выбора при этом должна снижаться: незачем быть очень уж привередливой, если ваша цель – спариться со множеством кавалеров, а не выбрать лучшего. Все это отлично согласуется с обсуждаемой гипотезой.
Наконец, исследователи обнаружили положительную корреляцию между MI и выраженностью у самцов признаков, способствующих стратегии “невзрачных проныр” (в противоположность “гордым красавцам”). А именно: у видов с высокими значениями MI самцы и мельче по размеру, и имеют более длинный гоноподий (копулятивный орган). Что ж, действительно, быть мелким и шустрым обладателем длинного копулятивного органа должно быть выгодно в ситуации, когда на вашу красоту и величие всем плевать. Показано, что малые размеры тела и длинные гениталии и впрямь помогают самцам живородящих рыб совершать “копуляции украдкой” (sneak copulations), незаметно подплывая к самкам сзади.
Таким образом, эти результаты хорошо согласуются с гипотезой Дэвида и Джин Це о влиянии плацентарности на направленность полового отбора. Впрочем, не до конца прояснен вопрос о том, где здесь причина, а где следствие. Переход к плацентарности рассматривается как причина редукции прекопуляционной женской избирательности и связанного с ней комплекса признаков (мужских украшений и брачных ритуалов). Но можно предположить и обратное: произошедшее по какой-либо причине снижение действенности прекопуляционного выбора (например, в результате развития у самцов эффективных адаптаций для “копуляций украдкой”) становится стимулом для развития механизмов посткопуляционного выбора – таких как избирательная “подкормка” оптимальных эмбрионов материнским организмом.

