Книга: История биологии с древнейших времен до начала XX века
Назад: Глава 35 Формирование микробиологии как самостоятельной науки
Дальше: Глава 37 Эволюционная теория во второй половине XIX века
Глава 36
Изучение процесса размножения клеток
Одной из центральных проблем биологии была и остается проблема возникновения клеток в течение индивидуальной жизни растительного и животного организма, непосредственно связанная с общей проблемой непрерывной преемственности жизни на Земле, а также с выяснением закономерностей индивидуального развития и наследственности. Клеточная теория обосновывает мысль о структурном единстве двух основных ветвей органического мира; это единство представляет одну из основ эволюционной доктрины, так как оно свидетельствует об общности происхождения растений и животных.
Представления о способах возникновения клеток до начала 70-х годов.
В эпоху создания клеточной теории на рубеже 40-х годов XIX в. еще не было сколько-нибудь обоснованных взглядов на способы возникновения новых клеток. Шлейден и Шванн с их теориями клеткообразования за счет бесструктурного вещества, или цитобластемы, недалеко ушли от фантастических представлений их непосредственных предшественников — К. Шпренгеля, Л. Тревирануса, Ф. Распайля, П. Тюрпена и др.
Противоречивость и часто полная необоснованность суждении относительно клеткообразования, высказывавшихся в 20-30-е годы, породила скептическое отношение (например, у О.П. Декандоля) к возможности выяснения этого процесса вообще. Впрочем, вскоре (1832) появилась работа Б. Дюмортье, впервые описавшего деление клеток нитчатых водорослей путем образования перегородки посередине удлинившейся перед делением клетки. Свои наблюдения Дюмортье противопоставил ошибочным представлениям авторов, писавших об образовании клеток из зерен, находящихся внутри или вне клеток.
Первое описание деления клеток, сделанное Дюмортье, звучит так: «Развитие водорослей осуществляется добавлением новых клеток к старым… Концевая клетка удлиняется…, затем во внутренней жидкости происходит образование перегородки, которая разделяет клетку на две части; нижняя из них остается стационарной, тогда как концевая снова удлиняется, затем возникает новая перегородка и т. д… Этот факт образования перегородки у нитчатых водорослей хорошо объясняет процессы происхождения и развития клеток, остававшиеся до сих пор неизвестными». Три года спустя Гуто Моль подтвердил наблюдения Дюмортье и изобразил процесс деления клеток у нитчатых водорослей. Эти открытия не сразу встретили признание отчасти потому, что упомянутые авторы не видели в делящихся клетках ядер, а после обнаружения Броуном обязательного наличия ядер в любых растительных клетках работы, в которых не было упомянуто о ядрах, считались неполноценными. Так же объясняется, по-видимому, успех теории клеткообразования, созданной Шлейденом (1838). Шлейден считал ядро важнейшей составной частью вновь образовавшейся клетки и поэтому предложил заменить неопределенное название «ядро» термином «цитобласт» (клеткообразователь), надолго удержавшимся в ботанической цитологии.
В период с 40-х и до 70-х годов представления о способах возникновения новых клеток оставались довольно смутными. Допускали как «свободное» клеткообразование, не связанное с предсуществующими клетками, так и различные формы образования клеток из ранее существовавших — путем почкования, эндогенного возникновения и т. п. Наряду с этим считалось также возможным и размножение клеток делением.
Примеры деления клеток обнаруживались прежде всего на растительных объектах. Ф. Унгер, считавший, что «происхождение и рост клетки, ее размножение… в растении является одним из наиболее таинственных жизненных процессов», привел в своей книге, напечатанной в 1840 г. в России и премированной Петербургской Академией наук, помимо данных Моля также и свои данные о делении клеток у водорослей и зародышей, в сердцевинных клетках и клетках волосков высших растений. «Рост древесины, — писал Унгер, — покоится на последовательном делении камбиальных клеток». Впрочем, наряду с делением он признавал и «новообразование клеток в гомогенном слизистом веществе». Представление об одновременном существовании двух способов образования клеток — деления и спонтанного их возникновения — разделяли К. Негели, Н. Прингсгейм, а вначале также и Э. Страсбургер.
На зоологическом материале вопрос о размножении клеток делением решался прежде всего при наблюдении дробления яиц (см. главу 14).
Механизм клеточного деления долго оставался невыясненным. Особенно неясным представлялось поведение ядер. Длительное время господствовало убеждение, что зародышевый пузырек, т. е. ядро ооцита, при созревании исчезает, а перед началом первого дробления в зрелом яйце появляется снова. Бедность хроматином ядер ооцита, зрелых яиц и первых бластомеров затрудняла наблюдения над этими ядрами не только на живых объектах, но и на фиксированных и окрашенных препаратах. И хотя внимательные наблюдатели уже в 60-е годы видели деление яйцевого ядра и ядер в бластомерах (например, см. И.И. Мечников «Эмбриологические исследования на насекомых», 1866), от внимания других исследователей (А. Вейсман, П.Т. Степанов и др.) деление ядер при созревании половых клеток и дроблении яиц ускользнуло.
Новообразование ядер допускалось еще в 70-е годы в эмбриональных клетках цыпленка (П.И. Перемежко) или в яйце стерляди (В.В. Заленский). Даже в конце 90-х годов можно было встретить утверждения, что клетки зародышевых листков возникают из безъядерных клеточных элементов (М.Д. Лавдовский, Н. Тишуткин).
Первые неполные описания митозов в начале 70-х годов.
К началу 70-х годов относятся первые серьезные исследования, касающиеся поведения ядер в делящихся клетках. Впрочем, еще в конце 40-х годов публиковались описания и рисунки, показывающие, что отдельные этапы сложных процессов непрямого деления клеток наблюдались уже тогда и были только неверно истолкованы. Так, В. Гофмейстер (1848, 1849, 1861) описывал в делящихся клетках фигуры, возникновение которых он сам объяснял коагуляцией белка, а на самом деле, как видно из его рисунков, он видел метафазы и анафазы митоза и изобразил эти фигуры как сбоку со стороны экватора, так и с полюса клетки.
На смену считавшейся одно время общепринятой «схеме Ремака», согласно которой при делении клетки сначала делится ядрышко, затем перетяжкой делится ядро и, наконец, клеточное тело, пришли наблюдения, касающиеся сложных явлений в ядре, предшествующих разделению тела клетки на две части перегородкой или перетяжкой. Открытие этих явлений применительно к растительным клеткам одно время безоговорочно связывали с именем московского ботаника И.Д. Чистякова. На самом деле работе Чистякова (1874) предшествовали исследования доцента Дерптского университета Э. Руссова по гистологии вегетативных и спорообразовательных органов, а также о развитии спор сосудистых тайнобрачных с соображениями о гистологии явнобрачных растений (1872). В материнских споровых клетках папоротников и хвощей, а также в материнских клетках пыльцы лилии Руссов видел на месте ядер круглые пластинки, состоящие из удлиненных, сильно преломляющих свет, палочек. У лилии эти «палочковые пластинки» состоят из особенно четких и крупных червеобразных телец. Когда материнская клетка споры или пыльцы разделяется посредине мелкозернистой протоплазменной перегородкой, с обеих сторон последней оказывается по отдельной «палочковой пластинке». Упомянув об аналогичных «палочковых пластинках», которые наблюдал Гофмейстер в материнских споровых клетках плауна Psilotum, Руссов заметил, что это образование не может быть артефактом, так как пластинки имеют правильную форму и характеризуются постоянным положением в клетках. В работе Руссова, едва ли не впервые, отчетливо описаны и изображены метафазные и анафазные пластинки, состоящие из отдельных хромосом, — картины, воспринятые этим автором как закономерное изменение клеточного ядра при его делении. Рассмотреть переход от одиночной «палочковой пластинки» к двойной Руссову не удалось.
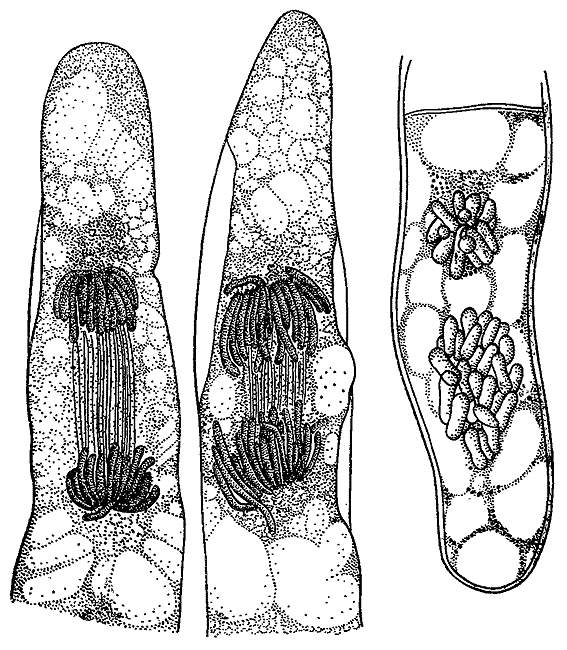
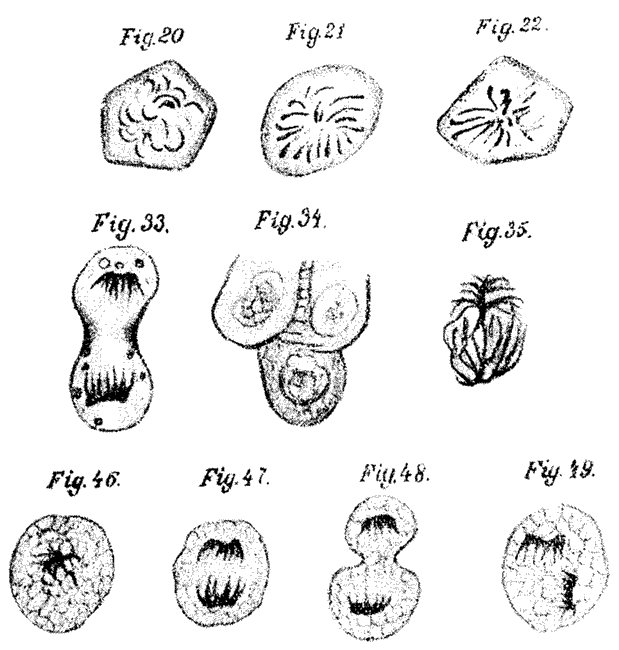
Телофаза и анафаза в клетках тычиночной нити традесканции. Справа рисунок Б. Гофмейстера. 1849.
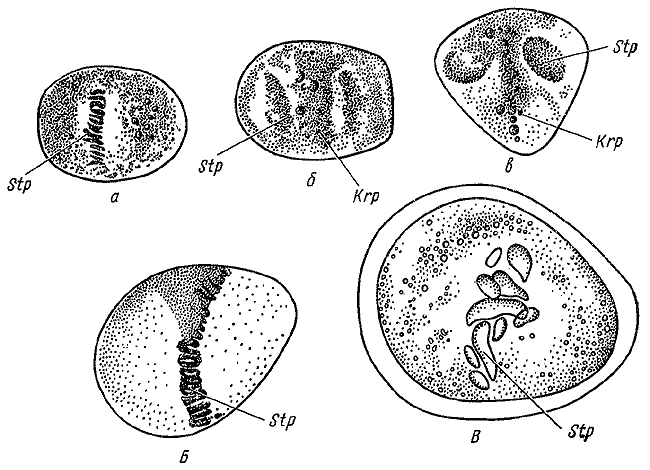
Деление клеток по Э. Руссову. 1872.
Еще более отчетливо деление клеток описал годом позже Руссова немецкий зоолог А. Шнейдер на дробящихся яйцах прямокишечной турбеллярии Mesostomum. В обширной статье Шнейдера, посвященной строению прямокишечных турбеллярий, их сопоставлению с остальными плоскими червями и системе плоских червей, описание дробления яиц занимает всего четыре страницы, но оно, бесспорно, явилось выдающимся событием в истории открытия непрямого деления клеток. По наблюдениям Шнейдера, разумеется не во всех деталях точных, ядро оплодотворенного яйца «превращается в скопление сильно изогнутых нитей», которые в свою очередь превращаются в «толстые тяжи, расположенные сначала беспорядочно, а затем в виде розетки, которая лежит в… экваториальной плоскости… Зернышки, находящиеся в яйце, собираются в… меридиональных плоскостях… Когда начинается деление надвое, число тяжей увеличивается так, что часть их направлена к одному, а часть к другому полюсу. Наконец ядро перешнуровывается, тяжи поступают в дочерние клетки. Ряды зернышек вытягиваются в длину и могут быть прослежены из одной клетки в другую». При следующих дроблениях «ядро и клетка проделывают те же изменения, как при делении надвое… Таким образом, яйцевая клетка превращается в скопление клеток…. из которых, в конце концов, строится зародыш… Эти наблюдения дают давно желаемое объяснение клеточного деления и особенно процессов дробления. Они впервые ясно показывают, какие сложные превращения может испытывать ядро (зародышевый пузырек) при клеточном делении». Из описаний и рисунков Шнейдера видно, что он в сущности установил основные стадии митоза — профазу, метафазу, образование ахроматиновой фигуры, увеличение числа хромосом перед расхождением и анафазу (раннюю и позднюю) и изобразил их в правильной последовательности. Шнейдер видел митозы не только в дробящихся яйцах, но и на поздних эмбриональных стадиях, а также у взрослых червей. Он справедливо полагал, что процессы, характерные для деления клеток у прямокишечных турбеллярий, имеют место и у других организмов.
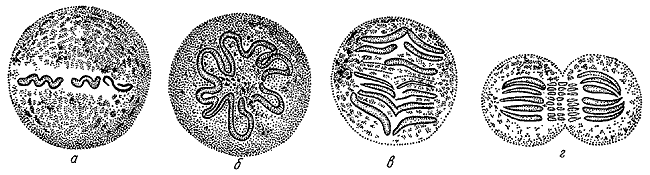
Деление клетки по А. Шнейдеру. 1873.
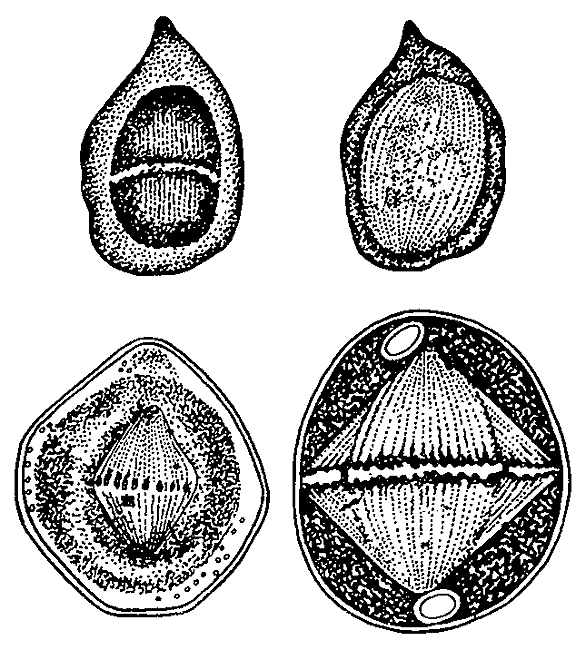
Деление клетки по И.Д. Чистякову. 1874.
Авторы исторических очерков, посвященных развитию представлений о делении клеток Э. Марк (1881) и Дж. Бейкер (1953–1955) также считают, что Шнейдер достаточно отчетливо, хотя, конечно, и не совсем полно, описал митотическое деление. Вполне четкой последовательной картины этого процесса ни Руссову, ни Шнейдеру нарисовать еще не удалось. Не удалось этого сделать и Чистякову. Из приведенных в его работе (1874) описаний развития спор у хвоща и плауна следует, что Чистяков видел ахроматиновое веретено, по экватору которого располагается зернистый пояс (иногда двойной), состоящий из хромосом — «сосочков», и что половина веретена («полусферы») превращается в дочернее ядро. Важной заслугой Чистякова является опровержение распространенного тогда мнения о растворении ядра материнской клетки и о «свободном образовании» ядер дочерних клеток, а также утверждение, что дочерние ядра образуются в результате деления материнского ядра; впрочем, эту мысль Чистяков проводил не везде с достаточной последовательностью. Преемственность фаз митоза осталась для Чистякова неясной. Еще менее отчетливо охарактеризованы Чистяковым процессы клеточного деления в статье о развитии спор и пыльцевых зерен (1875). Ядра в материнских споровых клетках, по его словам, «возникают внутри породившего их ядра, что может быть сравнено со свободным образованием клеток внутри плазмы материнской клетки». Экваториальную хромосомную пластинку в микроспоре Isoëtes Чистяков принял за образующийся фрагмопласт, т. е. пластинку, разделяющую дочерние клетки, тогда как Руссов сумел ясно отличить пограничное сгущение протоплазмы от материнской и дочерних хромосомных пластинок.
Детальные описания митозов во второй половине 70-х годов.
1875 г. следует считать переломным и для изучения процессов деления ядра, так как в этом году вышли работы Э. Страсбургера, О. Бючли и В. Майзеля, содержавшие более детальные, чем у Э. Руссова, А. Шнейдера и Д. Чистякова, описания митоза. Страсбургер в клетках зеленой водоросли спирогиры, в материнских клетках пыльцы лука и в материнских споровых клетках плауна, видел деление ядерной пластинки на две части, впоследствии расходящиеся. В своей книге, Страсбургер привел рисунки Бючли, не вошедшие в предварительное сообщение последнего (Бючли дал описание цитологических картин в дробящихся яйцах круглых червей и моллюсков и в сперматогенных клетках насекомых). Новых существенных деталей митоза Бючли не видел; Страсбургер сослался на описания рисунка Бючли с тем, чтобы подчеркнуть единство процессов клеточного деления у растений и животных.
Наблюдения Страсбургера и Бючли не замедлили подтвердиться в описаниях ассистента кафедры гистологии Варшавского университета В. Майзеля, который изучал строение клеток в роговице и коже лягушки, а также в роговице млекопитающих. Смысл наблюдавшихся ядерных структур сначала, как писал Майзель, был для него неясным и только после знакомства с данными Страсбургера и Бючли он понял, что имеет дело с разными стадиями деления клеток. Майзель не был уверен, в каком порядке следует располагать описанные им картины. Бесспорно, что он отчетливо видел профазу и метафазу, начало разделения материнской звезды на две дочерние с образованием ахроматинового веретена и, наконец, анафазу и процесс реконструкции дочерних ядер. Однако вместе с тем Майзель допускал и возможность «свободного образования ядер» из протоплазмы.
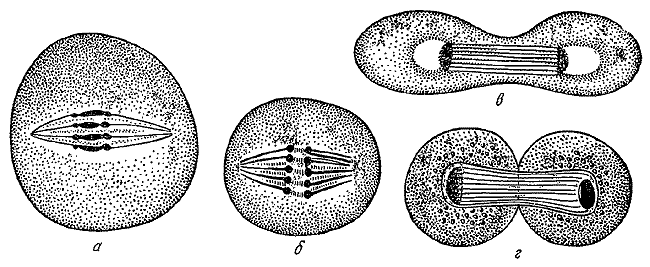
Деление клеток по Э. Страсбургеру. 1875.
Особенно насыщенным исследованиями, все ближе подводившими к правильному пониманию процессов деления ядра, был 1878 г., когда почти одновременно вышли предварительные сообщения В. Шлейхера, П.И. Перемежко и В. Флемминга, а также начало работы Майзеля (ее окончание и относящиеся к ней рисунки почему-то не были опубликованы); в конце 1878 г. и начале 1879 г. появились подробные работы Шлейхера (о делении хрящевых клеток амфибий), Флемминга (о размножении клеток в разных тканях саламандры и ее личинок) и Перемежко (о делении клеток в эпидермисе личинок тритона).
Шлейхер видел радиальное расположение образующихся в ядре «палочек» и «зерен» по экватору клетки; эти палочки претерпевают, по его мнению, поперечное деление и «почти с одинаковой скоростью отходят друг от друга». Для обозначения сложных процессов клеточного деления, имея в виду перемещения составных частей ядра, Шлейхер предложил термин «кариокинез». Перемежко главным образом на основании прижизненных наблюдений описал наряду с еще не делившимися и только что разделившимися ядрами «совершенно особые» ядерные образования, состоящие из нитей различной толщины, которые собраны в разнообразные звездовидные, клубкообразные, древовидные и иные фигуры, расположенные иногда вокруг одного центра; нередко в одной клетке имеются два таких образования, лежащие или рядом друг с другом, или на некотором расстоянии одно от другого. Описав и изобразив вне всякой последовательности эти структуры, Перемежко пришел к заключению, что они являются отдельными стадиями деления ядер, и предположительно наметил последовательность составляющих деление процессов.
Деление клеток эпидермиса хвоста тритона по П.И. Перемежко. 1879.
Более четкие результаты удалось получить Флеммингу, отчасти потому, что он избрал весьма удачный объект исследования — личинок саламандры. Флемминг ясно различал последовательное разрыхление клубка нитей, образующих содержимое ядра, а затем их утолщение; звездообразную установку петель ядерных нитей в экваториальной плоскости и разрыв каждой петли на периферии; продольное расщепление каждой нити и расхождение их половин с образованием дочерних звезд, которые снова переходят в стадию клубка, одновременно с чем тело клетки делится перетяжкой. «При образовании нового ядра, — писал Флемминг, — происходит повторение начальных фаз деления в обратном порядке». Непрямое клеточное деление Флемминг назвал митозом. Этот термин вскоре стал общепринятым.
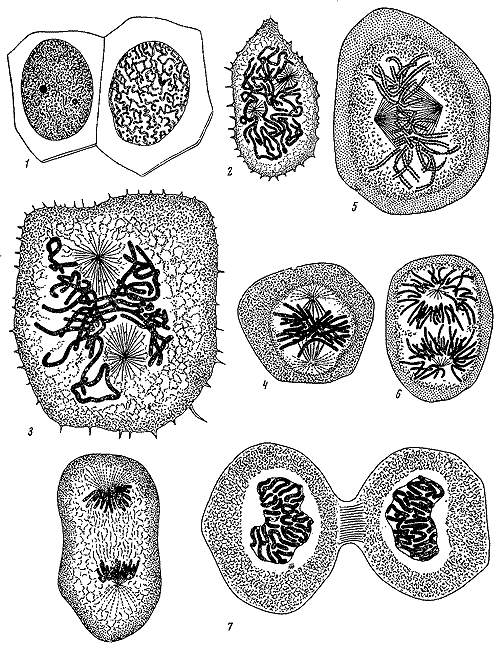
Деление клеток по В. Флеммингу. 1882.
Выяснение невозможности «свободного образования» клеток.
Б конце 70-х годов в последний раз в серьезной научной форме был поставлен вопрос, существуют ли иные способы возникновения клеток, кроме размножения делением, другими словами, вопрос — существует ли их «свободное образование». Страсбургер в монографии «Об образовании и делении клеток» (1875) признавал возможность «свободного клеткообразования» для ряда объектов, в особенности для зародышевого мешка цветковых растений. Однако в работе 1879 г. он должен был решительно пересмотреть прежние ошибочные представления. В этой статье Страсбургер писал: «Меня особенно радует, что я теперь могу правильно представить себе процесс, о котором я сам ранее сообщал неверные сведения. Свободное клеткообразование в зародышевых мешках не имеет места, все ядра возникают друг от друга посредством деления».
В начале 80-х годов Флемминг окончательно сформулировал представление о митозе как циклическом процессе, кульминационным пунктом которого является расщепление каждой хромосомы на две дочерние и распределение их между двумя вновь образующимися клетками. Представление о переходе одной из парных дочерних хромосом к одному, а другой — к другому полюсу ахроматического веретена митоза Флемминг высказал недостаточно отчетливо. В том виде, в каком оно впоследствии вошло в учебники, это представление сформулировали Э. ван Бенеден (1883) для животных и Э. Гейзер (1884) для растительных клеток. Разделение надвое каждого из хроматиновых зерен, из которых, по описанию Флемминга, состоит хромосома, вскоре показал на том же объекте — клетках личинки саламандры — В. Пфицнер (1881). Годом раньше О.В. Баранецкий установил спиральное строение хромосом, выделенных из материнских пыльцовых клеток традесканции. Эти открытия подготовили почву для разработанных цитологами и генетиками первой трети XX в. представлений о дискретном строении хромосом, об их спирализации и деспирализации во время митотического цикла.
Таким образом, начало нового периода в истории цитологии датируется рубежом 70-х и 80-х годов XIX в. С того времени отошли в прошлое споры о том, могут ли клетки, имеющие столь же длинную филогенетическую историю, как и состоящие из них растения и животные, возникать в онтогенезе из бесструктурного вещества.
Назад: Глава 35 Формирование микробиологии как самостоятельной науки
Дальше: Глава 37 Эволюционная теория во второй половине XIX века

