Книга: История биологии с древнейших времен до начала XX века
Назад: Глава 31 Развитие биогеографии, экологии и биоценологии
Дальше: Глава 33 Начало перестройки морфологии и систематики растении на эволюционной основе
Глава 32
Развитие эмбриологии растении
Крупные успехи эмбриологии растений во второй половине XIX в. были в значительной степени связаны с прогрессом в изучении клетки, с появлением эволюционной теории Дарвина, совершенствованием микроскопа и микроскопической техники. Учение Дарвина об эволюции органического мира, условия для восприятия которого в ботанике были уже подготовлены исследованиями Гофмейстера, стало теоретической основой эмбриологии растений и научных поисков ботаников-эмбриологов.
В последней четверти XIX в. было выяснено в общих чертах строение и развитие семяпочки, зародышевого мешка, пыльника и пыльцы. Был установлен факт прорастания пыльцы на рыльце пестика, образование пыльцевой трубки и ее прохождение по тканям столбика до семяпочки. Получил окончательное подтверждение и взгляд на оплодотворение как на соединение мужских и женских половых элементов. Однако оставалось неизвестным, каким образом осуществляется это соединение у семенных растений. Поэтому в конце XIX в. внимание ботаников в основном было сосредоточено на углублении исследований механизма оплодотворения, на изучении морфологической стороны этого процесса. Параллельно большое внимание было обращено на условия, необходимые для осуществления оплодотворения, на получение новых, более детальных данных относительно развития, строения и функций половых органов покрытосеменных растений.
Ч. Дарвин и раскрытие значения перекрестного опыления.
Для изучения процессов опыления и оплодотворения растений большое значение имели специальные труды Дарвина в этой области. Дарвин занимался наблюдениями и ставил опыты для выяснения механизма опыления и оплодотворения у растений, главным образом у орхидей. Он изучил структуру цветка, процесс опыления и разнообразные приспособления к нему у 150 видов, относящихся к 60 родам семейства орхидных. Он широко использовал экспериментальный и сравнительно-морфологический методы исследования.
В результате проделанной работы Дарвин установил, «что ни одно органическое существо не ограничивается самооплодотворением в бесконечном ряду поколений, но что, напротив, скрещивание с другой особью время от времени — быть может, через длинные промежутки времени — является необходимым». У покрытосеменных растений, как впервые обнаружил Дарвин, перекрестное опыление распространено гораздо шире, чем самоопыление. Многолетние опыты и наблюдения Дарвина показали, что перекрестное опыление обыкновенно оказывает на растения благоприятное действие, а самоопыление часто бывает вредным. Дарвин считал, что самоопыление является не основным, а вынужденным средством воспроизведения растениями семян при отсутствии условий для перекрестного опыления. Он установил, что семена, полученные в результате перекрестного опыления, крупнее, лучше прорастают, дают более жизнеспособное, сильное, рослое и плодовитое потомство, чем семена, полученные от самоопыления. Благодаря этому перекрестное опыление закрепилось естественным отбором в растительном мире и стало господствующим.
Дарвину удалось показать большое разнообразие приспособлений, выработанных у растений в ходе эволюции для устранения самоопыления. К числу этих приспособлений относится, в частности, особое устройство венчиков и тычинок, например у представителей орхидных, двудомность, дихогамия, гетеростилия, автостерильность. В то же время, в результате естественного отбора у растений закрепился ряд удивительных приспособлений для перекрестного опыления: у энтомофильных растений — форма и размеры цветка, окраска его лепестков, аромат, образование нектара; у анемофильных растений — большие качающиеся пыльники, неслипаемость и легкость пыльцы, большие и перистые рыльца. Цветки самоопыляющихся растений, таких как фиалка, кислица, яснотка, звездчатка, недотрога, сердечник, земляной орех и другие, наоборот, часто лишены запаха, имеют невзрачный вид и не открываются. Проанализировав историю происхождения этих приспособлений, Дарвин пришел к выводу о вторичности самоопыления и первичности перекрестного опыления. Он отметил, что приспособления, обеспечивающие перекрестное опыление, были приобретены растениями в процессе эволюции раньше, чем приспособления для самоопыления.
Дарвин вскрыл и причины положительного эффекта перекрестного опыления. По его представлениям, этот эффект определяется разнокачественностью подовых элементов родительских растений, развивающихся в разных условиях внешней среды. Отрицательное же действие самоопыления вызывается отсутствием «подобной дифференциации половых элементов». Дарвин неоднократно отмечал, что изменение среды оказывает могущественное действие на половые элементы растений. Он говорил, что воспитание материнских и отцовских растений в различных условиях внешней среды благоприятствует получению жизнеспособного и плодовитого потомства.
С эволюционных позиций подходил Дарвин и к решению вопроса о происхождении и сущности пола у растений. Важное значение полового процесса у растений Дарвин усматривал в соединении отличающихся друг от друга половых клеток отцовского и материнского растений, подвергавшихся «на протяжении предыдущих поколений» воздействию различных внешних условий. Появление половой дифференциации у растений сыграло, по его мнению, большую роль в истории всего органического мира. Много внимания уделил Дарвин и вопросу об эволюции форм размножения в мире растений.
Проблеме опыления и оплодотворения у покрытосеменных растений Дарвин посвятил три монографии — «Различные приспособления, при помощи которых орхидеи опыляются насекомыми» (1862), «Действие перекрестного опыления и самоопыления» (1876), «Различные формы цветков у растений одного и того же вида» (1877), а также несколько небольших статей. В этих работах Дарвин детализировал представления о поле растений и привел данные, имеющие важное значение для теории естественного отбора. Его труды обратили внимание ботаников на фактически забытую со времен Шпренгеля проблему биологии цветка и оплодотворения растений. Вслед за Дарвином многие исследователи занялись изучением структуры цветков и поиском у них приспособлений к перекрестному опылению. Возникла новая отрасль ботаники — биология цветка.
Ф. Гильдебранд, изучавший биологию цветка, опубликовал в 1867 г. книгу «Распределение полов у растений», в которой полностью присоединился к взгляду Дарвина на биологическую роль и широкое распространение в растительном мире перекрестного опыления. Ф. Мюллер, один из создателей биогенетического закона, в 70-е годы выявил ряд интересных приспособлений к опылению у цветков тропических растений.
Идеи Дарвина по вопросам опыления и оплодотворения у растений встретили горячую поддержку и получили дальнейшее развитие в России. А.Н. Бекетов изучал протерандрию у зонтичных. А.Ф. Баталин, которого знал и ценил Дарвин, провел большую серию работ по исследованию способов опыления и описал явления дихогамии и клейстогамии. Одновременно изучением биологии цветения в России занимались С.М. Розанов, И.П. Бородин, Я.Я. Вальц, В.И. Беляев и др. С пропагандой дарвиновских идей о значении перекрестного оплодотворения выступили С.А. Рачинский, И.П. Бородин, К.А. Тимирязев.
Однако наряду с ботаниками, давшими высокую оценку трудам Дарвина по биологии цветка и оплодотворению растений, Бэтсон, Вейсман, Гебель, Тревиранус, Генсло, Бюрк и другие подвергли критике дарвиновские выводы. В значительной степени эта критика была, обусловлена той дискуссией, которая развернулась во второй половине XIX в. вокруг проблемы наследственности. Следует, однако, заметить, что в ходе этой дискуссии наряду с ошибочными концепциями, извращавшими правильные представления о сущности полового процесса у растений, обсуждались новые интересные теории и соображения.
Изучение зародышевого мешка и пыльцевых зерен. Выяснение Э. Страсбургером и И.Н. Горожанкиным механизма оплодотворения.
Для дальнейшего развития эмбриологии растений многое сделали Э. Страсбургер, И.Н. Горожанкин, В.И. Беляев, С.Г. Навашин, Л. Гиньяр и другие, с именами которых связаны новые важные открытия в этой области ботаники.
Э. Страсбургер подтвердил генетическую связь папоротникообразных с голосеменными и покрытосеменными растениями, установленную еще Гофмейстером, и показал, что корпускулы (яйцеклетки) голосеменных соответствуют архегониям папоротникообразных. При изучении развития зародышевого мешка покрытосеменных растений Страсбургер выяснил ряд новых деталей в строении этого органа, ускользнувших от внимания предыдущих исследователей (Амичи, Гартига, Гофмейстера). Данные Страсбургера лежат в основе современных представлений о строении зародышевого мешка покрытосеменных.

Эдуард Страсбургер. 1844–1912.
Страсбургер нарисовал следующую картину возникновения и развития зародышевого мешка. Одна из клеток нуцеллуса семяпочки (какая именно, Страсбургер обнаружить не смог) дифференцируется как материнская. Затем она делится два раза и образует ряд из четырех, расположённых друг над другом, клеток. Три верхних, клетки вскоре отмирают, а самая нижняя увеличивается и становится исходной клеткой зародышевого мешка. Ядро этой клетки делится трижды, в результате чего образуются две группы по четыре ядра в каждой. Одна группа располагается у микропилярного, другая у халазального концов зародышевого мешка. Из ядер первой группы, как нашел Страсбургер, возникает яйцевой аппарат, состоящий из яйцеклетки, двух синергид и верхнего ядра. Из ядер другой группы возникают три антиподы и нижнее ядро. Страсбургер впервые установил, что нижнее и верхнее ядра, впоследствии названные Гиньяром полярными, сливаются и возникает одно вторичное ядро зародышевого мешка.
Страсбургер интересовался также вопросом о строении пыльцы покрытосеменных. К тому времени было уже известно, что в зрелой пылинке некоторых покрытосеменных растений имеются два ядра. Но представления о строении и назначении этих ядер были самыми неопределенными. Страсбургер установил, что одно из двух ядер пылинки — меньшее, лежит первоначально в маленькой линзовидной клетке, вблизи пыльцевой оболочки. Большее же ядро располагается в остальной части протоплазмы клетки пылинки. Однако в вопросе о дальнейшей судьбе этих клеток Страсбургер сделал ошибочные выводы. Во-первых, он считал, что по мере развития пылинки оболочка маленькой линзовидной клетки растворяется и оба ядра свободно располагаются в плазме пыльцевого зерна. Во-вторых, он признал меньшую клетку пыльцевого зерна за вегетативную, уподобив ее редуцированному заростку папоротникообразных и голосеменных. Большую же принял за генеративную. На самом деле меньшая клетка является генеративной, а большая — вегетативной, дающей пыльцевую трубку.
В 1875–1876 гг. Страсбургер описал процесс оплодотворения у голосеменных. Он был сторонником представления о диффузном характере оплодотворения, и это представление легло в основу его теории. Подобно Гофмейстеру, Страсбургер считал, что оплодотворение у голосеменных осуществляется через замкнутый конец пыльцевой трубки. По его мнению, половые ядра, содержащиеся в пыльцевой трубке, к моменту оплодотворения якобы растворяются, а образовавшееся ядерное вещество диффундирует через оболочки трубки и корпускулы (яйцеклетки) в плазму последней. Здесь ядерное вещество, по утверждению Страсбургера, уплотняется, восстанавливаясь в форменные элементы — семенные ядра. Вслед за тем одно из них сливается с ядром корпускулы. Продукт слияния — зародышевое ядро перемещается на дно корпускулы, делится и дает начало зародышу. Это неправильное толкование механизма оплодотворения в течение нескольких лет держалось в литературе.

Иван Николаевич Горожанкин. 1848–1904.
Ошибочность воззрений Страсбургера на оплодотворение у голосеменных была установлена в 1880 г. работами профессора Московского университета И.Н. Горожанкина, описавшего истинную картину оплодотворения голосеменных растений. На основании тщательного микроскопического изучения полового процесса у хвойных Горожанкин пришел к выводу, что оплодотворение осуществляется не диффузным путем, через замкнутый конец пыльцевой трубки, как это утверждал Страсбургер, а так же, как и у тайнобрачных, т. е. путем непосредственного слияния протоплазм. Горожанкин показал, что мужские половые ядра, или «спермагены» (по современной терминологии спермин), не растворяются в пыльцевой трубке, а через отверстия в ее оболочке проникают в яйцеклетку архегония, где и сливаются с ее ядром.
Окончательное подтверждение истинности подобного механизма оплодотворения у хвойных Горожанкин представил спустя три года. В 1883 г., работая в Германии в лаборатории известного миколога де Бари, Горожанкин смог, наконец, при изучении процесса оплодотворения у сосны наблюдать непосредственный переход половых ядер из пыльцевых трубок в яйцеклетку. Ему удалось получить микроскопический препарат, на котором часть мужского ядра помещалась еще в трубке, а другая часть — в протоплазме яйцеклетки архегония. Таким образом, проблема оплодотворения у семенных растений была выведена из тупика.
Эмбриологические работы Горожанкина, составившие эпоху в истории изучения полового процесса у высших растений, заставили Страсбургера пересмотреть свои прежние воззрения на оплодотворение. Руководствуясь новыми данными об оплодотворении у хвойных и используя более совершенную, чем в ранних работах, технику микроскопирования, он провел тщательные исследования процесса оплодотворения у подъельника и некоторых других цветковых растений и в 1884 г. полностью подтвердил выводы Горожанкина. Подобно Горожанкину, Страсбургер отчетливо наблюдал у покрытосеменных растений проникновение мужских половых ядер из пыльцевой трубки в зародышевый мешок и последующее слияние одного из них с ядром яйцеклетки. Отказавшись от представления о диффузном характере оплодотворения, Страсбургер исправил некоторые из своих старых ошибок и в отношении строения мужского гаметофита покрытосеменных. Он убедился, в частности, в том, что маленькая линзовидная клетка в пыльцевом зерне покрытосеменных является генеративной, а более крупная — вегетативной. Кроме того, он установил обязательное присутствие в пыльцевой трубке трех ядер: одного вегетативного и двух генеративных, возникающих в результате деления генеративного ядра у одних растений еще в пыльнике, а у других в пыльцевой трубке.
Дальнейшие исследования процесса оплодотворения. Работы В.И. Беляева, М. Трейба, С.Г. Навашина и других.
После классических исследований И.Н. Горожанкина открытия в эмбриологии растений следовали одно за другим. В 1891 г. В.И. Беляев, изучая процесс оплодотворения у хвойных, сделал сенсационное открытие. Он установил, что яйцеклетку хвойных оплодотворяет не большая клетка пыльцевого зерна, как это думали раньше, а маленькая. На этом основании Беляев пришел к выводу, что большая клетка пыльцевого зерна хвойных, дающая пыльцевую трубку, является гомологом клетки стенки антеридия разноспоровых папоротникообразных, а маленькая клетка — гомологом его внутренних сперматогенных клеток. Таким образом, приведя несомненные доказательства родственной связи между высшими споровыми и голосеменными растениями в отношении строения и развития мужского полового поколения, В.И. Беляев окончательно подтвердил воззрения Гофмейстера на единство растительного мира.

Владимир Иванович Беляев. 1855–1911.
Ценные исследования провел В.И. Беляев и по изучению сперматозоидов у высших споровых и голосеменных растений. В противовес утверждениям Страсбургера и Гиньяра он показал, что сперматозоид является не голым ядром, а настоящей клеткой с ядром и протоплазмой.
В 1891 г. в эмбриологии растений произошло еще одно важное событие. Ботаник М. Трейб опроверг общепринятое тогда мнение, что пыльцевая трубка всегда внедряется в семяпочку покрытосеменных через микропиле. Он нашел, что у экзотического семейства казуариновых пыльцевые трубки проникают в семяпочку через халазу. Основываясь на этом, как казалось в те времена, необычном явлении, Трейб предложил даже новую систему покрытосеменных растений. Он разбил покрытосеменные на два класса: халазогамных с казуариновыми в качестве единственных представителей и порогамных. Предложение Трейба встретило сочувственный прием у некоторой части ботаников, и было принято известным систематиком А. Энглером. Однако вскоре была доказана необоснованность подобного разделения покрытосеменных на классы. С.Г. Навашин показал, что в способах внедрения пыльцевых трубок в зародышевый мешок покрытосеменных нет единообразия и поэтому выделение казуариновых в отдельный класс не оправдано. Навашин обнаружил халазогамию у березы, лещины, грецкого ореха, граба, а у вяза установил существование переходной формы между порогамией и халазогамией, называемой теперь мезогамией.
Вслед за Трейбом и Навашиным изучением способов прохождения пыльцевых трубок в тканях пестика стали заниматься и другие исследователи. Главное же внимание по-прежнему привлекала проблема оплодотворения у покрытосеменных растений. Гиньяр исследовал половой процесс у цереуоа и лилии и наблюдал у них слияние яйцеклетки с одним из мужских половых ядер пыльцевой трубки. Кроме того, он выяснил особенности строения и развития пыльцы, семяпочки, зародышевого мешка, зародыша и эндосперма у ряда представителей цветковых. Однако многие детали самого акта оплодотворения оставались недостаточно выясненными. Все еще представляла собой загадку судьба второго мужского ядра пыльцевой трубки. Разрешен этот вопрос был уже на пороге XX в. благодаря классическим исследованиям Навашина.
Открытие С.Г. Навашиным двойного оплодотворения у покрытосеменных.
В противовес данным Шахта, Гофмейстера, Страсбургера, Гиньяра Навашин установил, что в процессе оплодотворения покрытосеменных растений из пыльцевой трубки в зародышевый мешок проникает не одно, а оба мужских половых ядра. Одно из этих ядер сливается с яйцеклеткой, другое же копулирует со вторичным ядром зародышевого мешка.

Сергей Гаврилович Навашин. 1857–1930.
Таким образом, в одном и том же зародышевом мешке одновременно совершаются два акта оплодотворения. В результате первого образуется зародыш будущего растения, а в результате второго — питательная ткань — эндосперм.
Рядом точных эмбриологических исследований на различных представителях покрытосеменных (лилейных, лютиковых, сложноцветных) Навашин убедительно показал, что эндосперм, как и зародыш, является продуктом полового процесса. Это необычное, свойственное лишь покрытосеменным растениям, явление он назвал двойным оплодотворением. О своем открытии он сообщил в августе 1898 г. на проходившем в Киеве X съезде русских естествоиспытателей и врачей, а в ноябре того же года опубликовал на эту тему небольшую статью в «Известиях Петербургской Академии наук».
Мысль о существовании двойного оплодотворения возникла у Навашина еще в 1895 г. в процессе работы над изучением халазогамии у грецкого ореха. Окончательное же подтверждение и оформление в стройную теорию эта мысль получила при исследовании оплодотворения у лилии Fritillaria. Позднее Навашин описал двойное оплодотворение и у других цветковых растений, систематически далеко отстоящих друг от друга — у представителей лютиковых, сложноцветных, орехоцветных, — доказав тем самым общность этого явления для всех покрытосеменных. Двойное оплодотворение и стало их отличительным признаком, отделившим покрытосеменные от голосеменных. Открытие двойного оплодотворения внесло ясность не только в вопрос происхождения эндосперма, но и разъяснило загадочность такого явления, как ксенип у кукурузы.
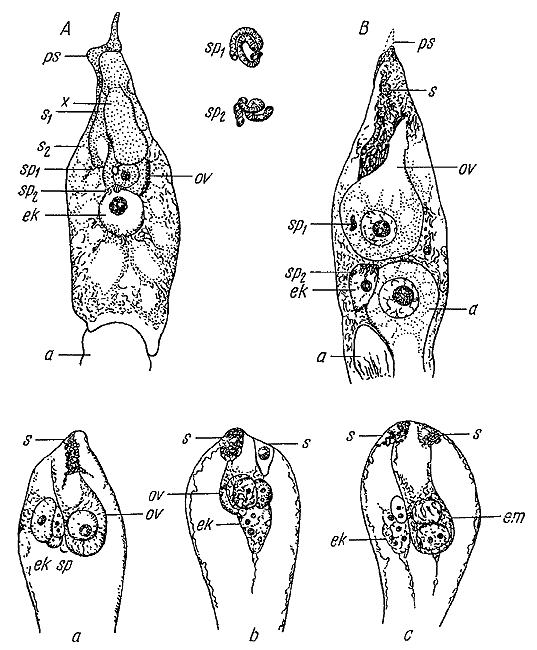
Двойное оплодотворение у растений Рисунки С.Г. Навашина.
Работы Навашина по оплодотворению покрытосеменных растений были встречены ботаниками всего мира с огромным интересом. В.И. Беляев, А.С. Фаминцын и М.С. Воронин оценивали открытие С.Г. Навашина как крупный вклад в науку, изменивший господствовавшие до того взгляды на оплодотворение у растений. Страсбургер назвал открытие двойного оплодотворения сюрпризом, делающим честь проницательности и наблюдательности исследователя, сделавшего это открытие.
После Навашина, с 1899 г. двойное оплодотворение у Leguminosae и некоторых других видов покрытосеменных стал усиленно изучать Гиньяр, а за ним и другие эмбриологи. Работы Гиньяра иллюстрировались хорошими рисунками. Вместе с тем в своей первой публикации о двойном оплодотворении в апреле 1899 г. он не упомянул об аналогичных исследованиях Навашина и претендовал на приоритет в этом открытии.
К выяснению факта двойного оплодотворения действительно близко подошли В.М. Арнольди, Гиньяр, Страсбургер, Мотье. Более того, некоторые из этих исследователей даже располагали микроскопическими препаратами с очень ясными картинами двойного оплодотворения. Но им не удалось сделать из них правильные выводы. Только после опубликования работ Навашина они поняли все значение и своеобразие оплодотворения покрытосеменных растений. Таким образом, заслуга в открытии двойного оплодотворения у покрытосеменных всецело принадлежит Навашину.
Открытие Навашиным двойного оплодотворения сразу же поставило вопрос, в чем состоит сущность этого своеобразного явления. Сам Навашин, как уже отмечалось, рассматривал процесс слияния спермы с центральным ядром зародышевого мешка как половой акт, вполне равноценный оплодотворению яйцеклетки и сравнивал это явление с полиэмбрионией. Гиньяр и Страсбургер развивали иной взгляд. Гиньяр утверждал, что процесс слияния второго спермин со вторичным ядром зародышевого мешка не является половым. Страсбургер расценивал это явление как вегетативное оплодотворение и только оплодотворение яйцеклетки принимал за генеративное.
Дискуссии о сущности двойного оплодотворения продолжались и в начале XX в. В ходе этих дискуссий, а главное благодаря новым исследованиям, выводы Навашина получили полное подтверждение и дальнейшее развитие.
Назад: Глава 31 Развитие биогеографии, экологии и биоценологии
Дальше: Глава 33 Начало перестройки морфологии и систематики растении на эволюционной основе

