Книга: История биологии с древнейших времен до начала XX века
Назад: Глава 28 Перестройка сравнительной анатомии на основе дарвинизма
Дальше: Глава 30 Развитие физиологии животных и человека
Глава 29
Развитие филогенетической систематики животных
Вторая половина XIX в. отмечена бурным развитием систематики.
С момента опубликования теории происхождения видов Дарвина вопрос о том, действительно ли в природе происходит эволюция, и являются ли современные животные и растительные организмы потомками общих предков, оказался в центре внимания биологов. Неудивительно, что научные интересы систематиков этого периода были сосредоточены главным образом на филогенетических исследованиях. Такая направленность исследований способствовала, с одной стороны, более полному обоснованию эволюции в органическом мире, с другой — привела к упрочению и распространению особого способа изображения филогенетических связей в виде так называемых родословных древ.
Метод изображения отношений организмов при помощи разветвленных схем применялся в самой общей форме и раньше. Намек на него встречается уже в работе П.С. Лапласа «Elenchus Zoophytorum» (1766). Ламарк в дополнениях в первому тому «Философии зоологии» привел графическое изображение родственных связей между различными группами животных в виде ветвящейся схемы (см. стр. 183). В 1829 г. Э.И. Эйхвальд в своем курсе зоологии поместил схему животного мира в виде ветвящегося дерева (см. стр. 277).
Представления Ч. Дарвина о принципах естественной систематики.
Вопросы систематики занимали в творчестве Дарвина значительное место. Ему принадлежит, в частности, систематическое исследование подкласса усоногих раков. Вполне понятно, что, занимаясь практической систематикой, Дарвин изучил наиболее распространенные в то время классификационные схемы. Например, уже в первой «Записной книжке» (1837–1838) Дарвин анализировал числовые циклические системы У. Мак-Лея и его продолжателей — Э. Фриса и Э. Ньюмена. Эти крайне искусственные системы привлекли внимание Дарвина тем, что в них отмечалось наличие аналогичных признаков у организмов, живущих в сходных условиях. В «Происхождении видов» Дарвин вернулся к анализу числовых систем, причем им руководило стремление выделить их рациональное зерно — идею параллелизма, иногда наблюдаемую «в числе подгрупп различных классов», которая появляется у видов различных классов в результате их приспособления «к жизни в одной из трех сред — на суше, в воздухе или в воде…» Дарвин писал, что «натуралист, сталкиваясь с параллелизмом подобного рода, произвольным увеличением или уменьшением значения групп в разных классах (а весь наш опыт говорит нам, что установление этого значения все еще совершенно произвольно), может легко расширить этот параллелизм, и отсюда, по всей вероятности, произошли различные системы классификаций, в основу которых положены параллельные деления на три, на четыре, на пять, на семь и т. д.». С пониманием относясь к идее числовых циклических классификаций, Дарвин в то же время прекрасно осознавал всю искусственность этих построений. На вопрос о том, какая система организмов может быть названа естественной и какие принципы должны быть положены в ее основу, Дарвин уже в 1842 г. определенно отвечал: «…естественная система должна основываться на генеалогии». Одновременно он указал на значение сравнительно-анатомических, эмбриологических и палеонтологических данных для выяснения истинного родства животных. Более полные соображения о принципах и методах естественной классификации были высказаны им в «Происхождении видов»: «… я думаю, — писал он, — что общность происхождения, единственная известная причина близкого сходства организмов, и есть та связь между ними, которая, хотя и выражена разными степенями изменений, до некоторой степени раскрывается перед нами при помощи наших классификаций». И далее: «расширив пользование идей общего происхождения, которое, несомненно, представляет собой единственную известную причину сходства организмов, мы поймем и то, что разумеется под естественной системой: это — попытки генеалогической классификации, в которой разные степени приобретенного различия выражаются в терминах — разновидности, виды, роды, семейства, отряды и классы».
Развитие филогенетической систематики Э. Геккелем.
Ревностным поборником филогенетического направления в систематике был Э. Геккель. Под влиянием его работ значение данных анатомии, палеонтологии и особенно онтогении для установления родства между систематическими группами получает широкое признание. Опираясь на метод тройного параллелизма, Геккель обосновал возможность построения естественной системы в виде генеалогического древа. Он же первый построил «обобщающее филогенетическое древо» органического мира.
Геккель пришел к выводу, что различные категории систематики являются искусственными подразделениями и что различия между ними не абсолютны, а относительны. Он рассматривал главные систематические категории как самостоятельные стволы (Stämme, Phyla), каждый из которых развивается из собственного единого корня. Все формы, относящиеся к одному стволу (типу), т. е. классы, отряды, семейства, роды и виды, рассматривались Геккелем как потомки общего «самовоспроизводящего первичного организма».
Основную цель филогенетических исследований Геккель видел в прослеживании родственных связей между формами и построении родословного древа органического мира. С помощью этих «древ» он стремился воссоздать общую картину эволюции. Родство между формами, относящимися к одному типу, Геккель усматривал не в «плане строения», а в их происхождении от общего предка. Благодаря наличию промежуточных ступеней они находятся друг с другом в непрерывной связи. Определяя сущность и значение естественной системы организмов, Геккель писал, что существует единственная сложная система организмов, выражающая реальные соотношения между ними. Эти реальные соотношения связывают все живущие и вымершие организмы в главные группы естественной системы, которые имеют генеалогическую природу. Их родство по форме есть и родство по крови. Вследствие этого естественная система организмов есть их родословное «древо», или «генеалогия».
Геккель делил органический мир на три царства — протистов, животных и растений. Его родословное древо начиналось единым стволом и завершалось детализированными ветвями для отдельных типов, классов и даже отрядов. В пределах ветвей были объединены вое организмы, в общем происхождении которых от единого предка Геккель не сомневался. Он утверждал, что ветви в самых общих чертах соответствуют тем «большим», или «главным, классам», для которых Дарвин также допускал только кровное родство. Величину расхождения этих ветвей он определял степенью отклонения родственных по крови организмов друг от друга и от общей родовой формы. Последовательно распространяя этот принцип на весь органический мир, Геккель вслед за Дарвином пришел к утверждению об общем происхождении обоих царств — животного и растительного.
В основании разработанного Геккелем родословного древа животного мира помещены так называемые монеры — воображаемые доклеточные безъядерные организмы. Далее следуют первичные одноклеточные организмы («амебы»), клеточные колонии («мореады») и полые шары («бластеады»). Следующий этап эволюции, приведший к образованию многоклеточных, связан по Геккелю, с появлением их общего гипотетического предка — гастреи. Следующими последовательными ступенями развития, приводящими в конечном итоге к человеку, являются плоские, круглые и кишечно-жаберные черви, первичнохордовые, бесчерепные, круглоротые, первичные (селахии), хрящевые и двоякодышащие рыбы, первичные земноводные, чешуйчатые гады, первичные пресмыкающиеся, млекопитающие-рептилии, первичные млекопитающие, сумчатые, полуобезьяны, собакоголовые и человекоподобные обезьяны и, наконец, обезьяноподобные люди.
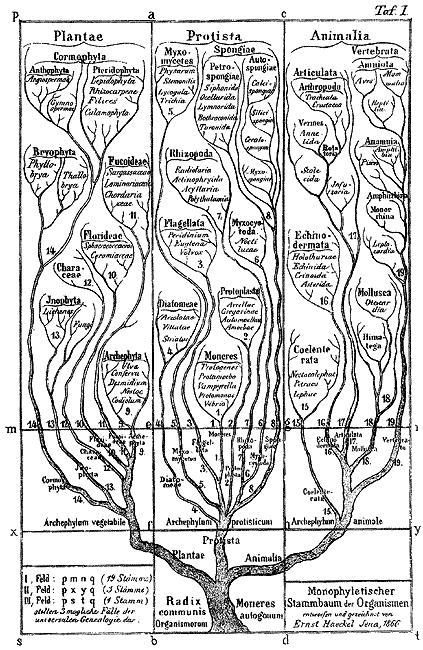
Филогенетическое древо по Э. Геккелю. 1866.
В филогенетических схемах Геккеля было много произвольного. Недостающие научные данные о переходных группах Геккель щедро восполнял богатой фантазией. Виною многих неточностей и прямых ошибок в его системе была склонность к поспешности в филогенетических обобщениях. Главный же источник погрешностей коренился в односторонности его метода. Геккель слишком переоценивал данные онтогенетического развития, придавая им абсолютное значение, а в самом онтогенезе видел только одну сторону — пассивное отражение филогенеза. Однако в целом исследования Геккеля сыграли выдающуюся роль в разработке филогенетической систематики.
Эмбриологическое направление в систематике.
Наряду с филогенетическим направлением в систематике второй половины XIX в. продолжали развиваться эмбриологическое и традиционное типологическое направления.
Как было указано выше (см. главу 15), основы эмбриологического направления в систематике были заложены еще в 20-е годы XIX в. К.М. Бэром. Особое развитие это направление получило под влиянием эволюционного учения. В данных эмбриологии Дарвин и его последователи видели наиболее веские и точные доказательства родства организмов.
«Даже два главных подразделения усоногих — стебельчатые и сидячие, — отмечал Дарвин, — как ни сильно разнятся они по своему наружному виду, имеют личинки едва различимые на любой стадии их развития».
Крайним выражением этого направления было построение ряда классификационных схем на основе одного эмбриологического критерия. Примером может служить искусственная система животных крупнейшего бельгийского гистолога второй половины XIX в. Э. ван Бенедена. Она была основана на различных формах дробления желтка. Исходя из одного этого признака, ван Бенеден выделил не только классы, но даже разбил отдельные семейства (например, семейство бокоплавов) на несколько групп, распределив их по различным отделам.
Идея создания естественной системы животных на основе одного лишь эмбриологического критерия была так сильна, что ей отдали дань даже Геккель и Гексли. Так, в 1873 г. Геккель, подобно ван Бенедену, предложил систему животных, базировавшуюся целиком на различных формах дробления желтка. Односторонний подбор систематических признаков снова привел к неудаче. Так, в одном подразделении оказались животные (губки, медузы, нематоды, многие иглокожие, плеченогие, оболочники и бесчерепные), относящиеся не только к различным классам и отрядам, но даже к разным типам. В другом подразделении вместе с сифонофорами, ктенофорами, планариями, кольчатыми червями и большинством моллюсков находились круглоротые, ганоидные и амфибии.
Более удачную эмбриологическую систему предложил Т. Гексли (1874). При ее построении он впервые ввел понятия вторичноротые (Deuterostomxa) и энтероцельные (Enterocoelia). В дальнейшем систематики использовали эти термины в несколько ином толковании. Однако сама попытка Гексли классифицировать животных на основе таких черт развития, как способы образования полости тела и вторичного рта, свидетельствовала об осознании им необходимости использования в систематике филогенетических признаков.
В системе Гексли Deuterostomia в зависимости от способа образования вторичной полости тела подразделялись на:
1) Schizocoelia, у которых полость рта образуется путем расщепления мезодермы (Arthropoda, Annelida, Mollusca, Bryozoa);
2) Enterocoelia, у которых полость тела развивается энтероцельным путем, т. е. за счет выпячивания кишечника (Chaetognatha, Brachiopoda, Enteropneusta, Echinodermata);
3) Epicoelia, у которых внутренняя и внешняя полости тела имеются хотя бы на стадии личинки (Tunicata, Leptocardia, Vertebrata).
Гексли относил ко вторичным животным Polychaeta, Arthropoda, Mollusca, Brachiopoda и Bryozoa, считая, что дефинитивный рот является у них новообразованием, а полость тела возникает в результате расщепления мезодермы. Этой группе в его системе противопоставлялись животные с общей полостью тела — Scolecimorfa, которые объединяли Platodes, Nematodes, Olygochaeta, Hirudinea, Rotifera и Gephyrea. Такое подразделение было очень неудачным и повлекло за собой ряд ошибок. Например, класс пиявок (Hirudinea) был выведен из подтипа кольчатых червей (Annelida) и искусственно перенесен в группу Scolecimorfa.
Выяснить причины неудач, постигших авторов эмбриологических классификаций, пытался уже Ф. Мюллер. Он отмечал (1864), что «эмбриоманы» весьма односторонне подходили к оценке эмбриональных признаков организмов, совсем не учитывая тех отношений и соотношений, которые возникали между развивавшимся индивидуумом и внешними условиями. Он указывал, что так как эмбриология еще только начинала свое развитие, то многие положения этой науки, на основании которых строились классификации, еще не были окончательно установлены, а иногда были просто неверными.
Таким образом, все попытки построения эмбриологических классификаций окончились неудачно. В дальнейшем от эмбриологических систем в таксономии сохранилось очень немногое. Примером могут служить такие подразделения, как Diploblastica и Triploblastica, Protos tomia и Deuterostomia.
Пересмотр основных типов в систематике животных.
После того как к концу XIX в. в основном закончился период бурного открытия и описания новых видов, родов и отрядов, систематики начали пересматривать и перестраивать уже существующие таксономические группы. Такие перестройки не всегда были оправданны. Нередки были случаи, когда дополнительно вводимые подразделения, как, например, группа Nephridia в системе Перье (1893) или тип Appendiculata в классификации Ланкестера (1900), носили искусственный характер и лишь вносили путаницу в естественные отношения между организмами. Понятно, что подобные группировки не могли удержаться в систематике на сколько-нибудь продолжительное время. Однако ряд систем, в первую очередь схемы, предложенные Гексли (1875), Клаусом (1877), Геккелем (1894–1896) и Ланкестером (1890–1891), имели большое распространение и являлись в какой-то мере исходными для будущих построений.
В классификации, предложенной Гексли, животные разделялись на восемь типов:
1) Protozoa (Rhizopoda, Sporozoa, Porifera); 2) Infusoria (Infusoria + Mastigophora); 3) Coelenterata; 4) Annuloida (Plathelmintbes, Nemathelminthes, Rotifera, Echinodermata); 5) Annulosa (Arthropoda и Annulata); 6) Molluscoidea (Brachiopoda, Polyzoa и Tunicata); 7) Mollusca; 8) Vertebrata.
Как видно из приведенной классификации, иглокожие были объединены с низшими червями (плоскими и круглыми), сюда же отнесены коловратки. Такое странное объединение объяснялось тем, что в данном случае учитывались лишь данные сравнительной эмбриологии, обнаружившие якобы сходство личинок плоских червей, иглокожих и коловраток. Губок Гексли рассматривал как колониальных простейших, вследствие чего они были отнесены к типу Protozoa. В системе была еще одна существенная неточность: простейшие разделялись на два типа — Protozoa и Infusoria. К последним были присоединены жгутиковые. В основе такого деления лежало ошибочное представление о том, будто макронуклеус инфузории является яичником, а микронуклеус — семенником.
В 80-90-х годах XIX в. широкое распространение получила система К. Клауса, которая в упрощенном виде использовалась даже специалистами нашего века. Клаус (1877) разделил весь животный мир на девять типов: 1) простейшие; 2) кишечнополостные вместе с губками; 3) иглокожие; 4) черви; 5) членистоногие; 6) моллюски; 7) моллюсковидные; 8) оболочники и 9) позвоночные. В этой системе губки еще не были выделены в самостоятельный тип, а продолжали рассматриваться как отдельный класс кишечнополостных. В системе, предложенной в 1894 г. Геккелем, количество типов было увеличено до десяти. Губки впервые были выделены в особый тип, который сохранился до настоящего времени.
Еще более сложной была система Ланкестера, в которой количество типов достигало 11. Весь животный мир был разбит в ней на два подцарства — простейших с одним типом и многоклеточных, куда входили все остальные типы. Многоклеточные животные в свою очередь составляли два раздела: Parazoa (только губки) и Enterozoa (все остальные многоклеточные). В Enterozoa были выделены две «ступени» — Enterocoela — кишечнополостные и Coelomata, куда входили вое типы, кроме простейших, губок и кишечнополостных. Черви были разбиты на три самостоятельных типа — плоских, круглых и кольчатых. Это деление сохранилось во многих современных системах.
Создание «больших» естественных систем.
В итоге к концу XIX в. в систематике животных благодаря прогрессу филогенетического направления были созданы необходимые предпосылки для построения «большой» естественной классификации животных, основанной на генетических признаках. Попытки построения такой системы явились конкретным воплощением идеи создания целостной картины филогенетического родства организмов, которая имела широкое распространение среди систематиков конца XIX — первой половины XX в.
При создании «больших» систем учитывалось несколько главных признаков строения животных (образование дифинитивного рта, способ закладки вторичной полости тела, соотношение главной оси тела и оси гаструлы), определявших их генетическое родство. В соответствии с этими признаками К. Гроббен (1908) и Б. Гатчек (1888, 1911) подразделяли двусторонне-симметричных животных на первичноротых (Protostomia) и вторичноротых (Deuterostomia), энтероцельных (Enterocoelia) и энтероцельных (Ecterocoelia), первичноостных (Protaxonia) и разноостных (Heteraxonia).
Важную роль при выделении высших таксономических категорий сыграла «трохофорная теория», автором которой был Б. Гатчек (1888). Согласно этой теории, большое филогенетическое значение придавалось личинке кольчатых червей — трохофоре, которая рассматривалась как «онтогенетическое повторение» (В.А. Догель, 1947) строения предков кольчатых червей. Наличие стадии трохофорообразной личинки и сходное дробление яиц связывало плоских червей и коловраток, с одной стороны, с кольчецами и моллюсками, с другой.
Все «большие» зоологические системы подразделяются обычно на две группы в зависимости от того, какое значение авторами этих систем придается признакам вторичноротости и энтероцельному способу закладки целома. К первой группе относят системы Гатчек-Гроббеновского направления, сторонники которого отделяли первичноротых от вторичноротых на уровне кишечнополостных. К. Шнейдер (1902) пошел еще дальше; он утверждал, что первичноротые и вторичноротые животные имеют независимое происхождение. Авторы систем второй группы считали, что признаки вторичноротости и энтероцельного способа закладки целома у животных, примыкающих к червям, развивались конвергентно. Поэтому в системах этого направления энтероцельные вторичноротые животные вместе с энтероцельными и первичноротыми объединялись в один тип и даже в один класс.
Рассмотрим вначале системы второго направления, берущие свое начало от классификаций Каруса, Гексли, Геккеля и Ланкестра.
Последователь Геккеля зоолог А. Ланг, развивая гипотезу о происхождении турбеллярий от ктенофорообразных предков, использовал ее для объяснения филогенеза кольчатых червей. В опубликованной им системе (1888–1894) беспозвоночные были разделены на семь типов. Первый тип — простейшие; второй тип — зоофиты, или кишечнополостные, куда в виде одного из классов были включены губки; третий тип — плоские черви состоял из трех классов: турбеллярий, сосальщиков и лентецов; четвертый тип червей объединял шесть классов: немертин, круглых червей, кольчецов, переднеанальных, а также коловраток и щетинкочелюстных; пятый тип — членистоногие; в шестом типе были сгруппированы моллюски; в седьмом — иглокожие. В типе червей были объединены круглые и кольчатые черви с червеобразными коловратками, щетинкочелюстными, которые являлись переходными группами ко вторичноротым животным. Подобное же деление червей на два типа (Platodes и Vermes) встречается в системе В. Кюкенталя (1898). Таким образом, Ланг и Кюкенталь смешивали в объемистом типе первично- и вторичноротных, энтеро- и эктероцельных животных.
Среди русских зоологов «трохофорную теорию» поддерживал Н.А. Холодковский, что нашло отражение в предложенной им системе животных (1905). Тип Vermes состоял у него из пяти подтипов и объединял червей, начиная от бескишечных турбеллярий и кончая кольчатыми. Кишечнодышащие животные были в виде дополнения отнесены к типу иглокожих. Но сами иглокожие помещались между первичноротыми животными на большом удалении от типа хордовых. Последний был разделен на три подтипа — оболочников, бесчерепных и позвоночных. Губок и червеобразных Холодковский выделял в отдельные типы.
В системах первого направления была сделана попытка сочетать порядковое перечисление типов, филогенетическое «древо» и удобную определительную таблицу, что привело к созданию дробных систематических категорий (Cladus, Subregnum, Divisio, Phylum и Subphylum). Эти неоправданные усложнения вызвали, как выразился Н.Н. Плавильщиков, «перетасовку» крупных таксономических категорий и исказили истинные объемы типов.
Основоположник этого направления Б. Гатчек в 1888 г. предложил классификацию, основанную на принципах симметрии и на трохофорной теории. Многоклеточные животные разделялись у него на первичноостных (Protaxonia), куда входили губки, стрекающие, ктенофоры, и разноостных (Heteraxonia), объединявших трохофорных, иглокожих и хордовых. В группу трохофорных (Zygoneura) Гатчек ввел червей, червеобразных, моллюсков и членистоногих. Этих животных помимо трохофорной стадии развития или трохофорной организации взрослого животного объединяла общность строения двусторонне-симметричной нервной системы. В этой системе почти полностью было выдержано разделение животных на первично- и вторичноротых.
В 1911 г. Гатчек предложил второй вариант классификации, в котором ясно проявилось желание автора совместить определительную таблицу и филогенетическую систему. Он применил своеобразный принцип формирования высших группировок на основании «плана строения», «типа развития» и «типа организации». Вследствие этого система Гатчека отличалась серьезными изменениями в объемах типов, а подтипы четвертого подразделения, «тин организации», представляли сочетание типов, подтипов и классов обычных систем.
Но попытка Гатчека изменить объем систематических категорий оказалась неудачной. В этой системе он провел разграничение стрекающих и ктенофор, считая, что эти ветви полостных животных отличаются различными способами образования мезобласта, вследствие чего генетически они якобы совершенно независимы. Все черты их сходства, по мнению Гатчека, можно было объяснить явлениями конвергенции. Таким образом, Гатчек и Шнейдер воплотили на конкретном материале идею дифелитического происхождения многоклеточных животных, выдвинутую еще Ламарком.
Идеи Гатчека положил в основу своей системы К. Гроббен (1905). Эта система в переработанном и дополненном виде была опубликована им совместно с К. Клаусом в 1916 г. В этой классификации также имелись надтиповые группировки (Subregnum, Divisio, Phylum, Subphylum). Отличие этой системы от гатчековской заключалось в той трактовке, которая давалась в ней понятиям Ecterocoelia, Enterocoelia, Protoslomia, Deuterostomia. Гатчек в своей системе разделил червеобразных на две группы, одну из которых (Brachiopoda) он отнес к Enterocoelia, другую (Phoronida и Bryozoa) — к Ecterocoelia. Учитывая, что все Vermoidea — Tentaculata являются первичноротыми животными, а способ закладки целома у Phoronida и Bryozoa еще окончательно не установлен, специалисты признают разделение типа искусственным. Гроббен (1916) сосредоточил весь ряд (Cladus) Tentaculata в типе (Phylum) первичноротых животных, что является более естественным. Основные подразделения систем Гатчека-Гроббена были использованы многими зоологами (А. Гетте, К. Гейдером, В.М. Шимкевичем и др.).
* * *
В итоге можно сказать, что попытки систематиков второй половины XIX в. построить «большую естественную классификацию» животных не увенчались успехом. Главная причина неудачи состояла в том, что большинство авторов стремилось совместить в одной схеме два направления систематики — филогенетическое и типологическое. Сочетание в их системах отдельных этапов филогенетического развития и чисто классификационных двухмерно расположенных таксонов часто приводило к созданию таких группировок, которые не отражали естественных генеалогических связей между животными.
Это порождало споры среди зоологов о характере создаваемых систем — считать ли их искусственными или естественными. Л.А. Зенкевич писал в связи с этим: «В настоящее время всякая система является более или менее искусственной; чем более она основана на филогенетических взаимоотношениях, тем она менее искусственна: мерилом же искусственности системы является недостаточное использование признаков, свойственных организму, и неправильная их оценка. Филогенетическая система должна быть синтезом наибольшего возможного числа признаков. И в силу этих моментов искусственных систем может быть сколько угодно, а филогенетическая система только одна, поэтому правильнее всего эту последнюю называть естественной, т. е. дающей представление о естественной близости между отдельными группами».
Дальнейшее расширение и совершенствование методов выявления генеалогической близости между отдельными группами, а тем самым достижение новых успехов в построении естественной классификации животных произошло уже в XX столетии.
Назад: Глава 28 Перестройка сравнительной анатомии на основе дарвинизма
Дальше: Глава 30 Развитие физиологии животных и человека

