Книга: История биологии с древнейших времен до начала XX века
Назад: Глава 13 Возникновение палеонтологии
Дальше: Глава 15 Развитие систематики животных
Глава 14
Индивидуальное развитие животных
Общее состояние эмбриологии животных к началу XIX века.
Кризис «старой метафизики», отчетливо наметившийся во второй половине XVIII в., о чем подробно говорилось в предыдущих главах, проявился в этот период и в учении об индивидуальном развитии организмов.
Почти одновременно с «нападением» Канта (1755) на учение о вечности солнечной системы Каспар Вольф (1759) выдвинул принцип эпигенеза, согласно которому развитие организмов представляет собой не просто развертывание и рост зачатков, изначально присутствующих в зародыше, но и процесс новообразования. Тем самым была заложена основа науки о развитии индивидуума — эмбриологии. Время между работами Вольфа и тем периодом, когда эмбриология благодаря трудам Пандера, Бэра, Ратке и Рускони оформилась в самостоятельную отрасль биологии, отмечено главным образом распространением натурфилософий, сыгравшей в истории учения об индивидуальном развитии в значительной мере отрицательную роль. Натурфилософы придавали значение не столько эмпирическому изучению явлений природы, сколько интуиции и суждениям по аналогии. На этих путях делались произвольные и часто совершенно фантастические допущения, наряду, впрочем, с правильными, но не аргументированными догадками. Подобного рода догадками натурфилософы (Окен в Германии, Велланский в России) подменяли подлинное знание также и в области явлений развития зародыша. Поэтому столь важно было проследить эти явления на легко доступном объекте, описать их и точно изобразить с помощью тщательно выполненных рисунков. Многие детали развития, особенно относящиеся к ранним стадиям, оставались неизвестными и требовали тщательного изучения. По инициативе профессора Вюрцбургского университета И. Деллингера это исследование провел в 1816 г. уроженец Риги X.И. Пандер, который вместе с К.М. Бэром изучал у Деллингера сравнительную анатомию. Только в работах Пандера и особенно Бэра учение о развитии зародыша впервые встало на почву хорошо установленных фактов, и одновременно в эмбриологию был введен наряду со сравнением отдельных стадий метод прослеживания всего процесса развития зародыша на всех его стадиях, начиная от яйца.
Исследования X. Пандера. Открытие зародышевых листков.
Сочинение Пандера «Об истории превращений насиженного яйца в течение первых пяти дней» (1817) сообщало новые факты и продемонстрировало новые методы исследования развивающегося зародыша (отделение желточной оболочки от бластодермы, а бластодермы от желтка при погружении его в воду; наблюдение с помощью лупы и сложного микроскопа в падающем свете на черном фоне); оно иллюстрировано превосходно выполненными рисунками.
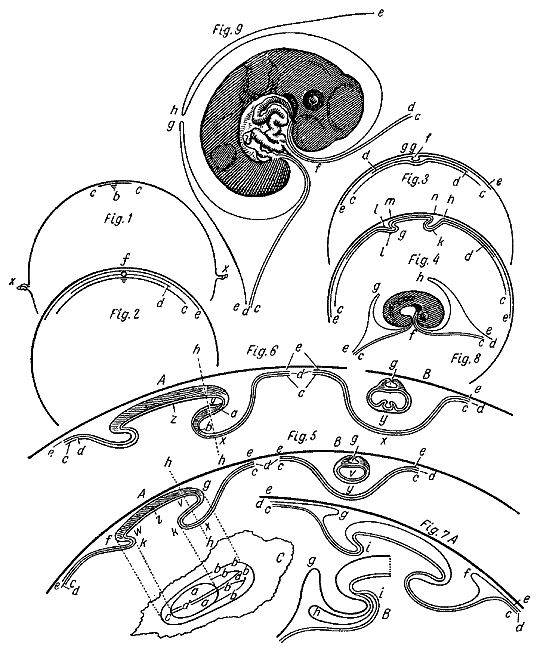
«Идеальные разрезы» по Пандеру. 1818.
Еще более поучительными были схемы, которые Пандер приложил к своей статье «Развитие цыпленка» (журнал «Isis», 1818), раскрывавшие соотношение зародыша с его оболочками. Важнейшим итогом наблюдений Пандера явилось утверждение, что ранний куриный зародыш состоит из двух слоев: наружного — серозного и внутреннего — слизистого; к ним позднее присоединяется средний, сосудистый, слой. Упомянутые слои бластодермы, по мнению Пандера, являются единственным источником образования различных органов зародыша и его оболочек. Пандер показал, что из серозного слоя развивается стенка тела и амнион, а из слизистого и сосудистого слоев — кишечный канал и брыжейка. Он подтвердил наблюдение Вольфа, впервые показавшего, что кишечный канал возникает из свертывающегося в трубку первоначально распластанного на желтке слоя бластодермы. Вольф также видел, что бластодерма состоит из лежащих друг на друге слоев, однако он не сделал из этого наблюдения существенных выводов. Только Пандер со всей определенностью высказал мысль, что эти слои или листки бластодермы (он часто называл их оболочками) являются зачатками определенных систем органов зародыша. Пандер установил далее, что первый признак формирования зародыша из гладкой до того бластодермы — это возникновение первичных складок. Пандеру, впрочем, не удалось ни выяснить смысла этих образований, ни установить их связь с развитием головного и спинного мозга, так как он ошибочно принял хорду за зачаток спинного мозга.
Открытие яйца млекопитающих и последующие эмбриологические исследования К. Бэра.
Бэр имел возможность непосредственно следить за работой Пандера и обсуждать с ним результаты наблюдений над развитием куриных зародышей. Однако не личное общение с Пандером, ни внимательное чтение его работы не привело Бэра к ясному пониманию описывавшихся Пандером явлений. Бэр пришел к выводу, что развитие зародыша требует более углубленного изучения, однако, он сам смог взяться за эту работу лишь несколько лет спустя. Исследуя эмбриональное развитие птиц и млекопитающих, Бэр сделал в 1827 г. важное открытие: он обнаружил яйца в яичнике млекопитающих и человека. До этого за яйца принимали крупные пузырьки на поверхности яичника, которые в XVII в. описал голландский анатом Р. де Грааф. Сначала у собаки, а затем и у других млекопитающих (коровы, свиньи, овцы и кролика, а также у человека) Бэр установил, что истинным яйцом является маленькое округлое тельце, прилежащее изнутри к стенке фолликула яичника, или граафова пузырька.
Вслед за сочинением, в котором было описано это открытие, Бэр выпустил первый том своего классического труда «История развития животных. Наблюдения и размышления» (1828), в котором детальное описание развития куриного зародыша сопровождалось теоретическими рассуждениями об общих закономерностях строения и развития животных. Бэр подтвердил и уточнил наблюдения Пандера, показав, что зародыш на ранней стадии разделяется на два слоя, или листка — серозный и слизистый, между которыми вскоре закладывается средний, сосудистый, листок. Одновременно с этим обнаруживаются первые признаки формирования зародыша — появление «первичной полоски», из которой образуются продольные валики, вскоре смыкающиеся друг с другом. Из спинных валиков, как показал Бэр, развивается центральная нервная система и сегментирующиеся зачатки осевого скелета и мышц. На дне желобка, разделяющего эти валики, находится тонкий тяж — хорда. Последняя сохраняется на всю жизнь у хрящевых рыб, а у высших позвоночных имеется только в зародышевом периоде. Образование хорды куриных зародышей, как справедливо отметил Бэр, является наиболее ранним признаком, характерным для зародышей всех позвоночных. Все три слоя распластанного на желтке зародыша разрастаются по поверхности желтка и смыкаются на его нижнем полюсе. Прилегающий к желтку слизистый листок образует затем желобовидное впячивание вдоль лежащей над ним хорды, давая начало закладке пищеварительной системы. После этого передний, а затем и задний концы зародыша приподнимаются, и он начинает отшнуровываться от желтка. В головной части зародыша нервная трубка образует расширения — мозговые пузыри (зачаток головного мозга); боковые выросты переднего мозгового пузыря дают начало глазам.
Проследив в деталях дальнейшее развитие зародыша, Бэр описал формирование всех систем его органов. Наблюдения над развитием куриного зародыша, описанные в первом томе «Истории развития животных», еще более детально изложены во втором томе этого труда, опубликованном в 1837 г. Теоретические размышления, заключающие первый том, касаются, прежде всего, общей характеристики трех основных процессов развития, которые Бэр назвал «внутренним преобразованием особи». На всех этапах формирования животного развитие, по мнению Бэра, идет в направлении от гомогенного (однородного) состояния к гетерогенному (разнородному) или от общего к частному. Бэр различал три формы возникновения гетерогенного, т. е. дифференцирования, которые он назвал первичным, гистологическим и морфологическим обособлением. Первичное обособление — это разделение раннего зародыша на слои — серозный, сосудистый и слизистый. Гистологическое обособление в каждом из отделившихся друг от друга листков приводит к образованию различных тканей. Морфологическое обособление расчленяет однородные вначале закладки систем органов на сформированные органы. Например, первичная пищеварительная трубка разделяется на пищевод, желудок, разные отделы кишечника, на закладки печени, легких и временного зародышевого органа — аллантоиса. Организация зародыша любого позвоночного может быть, по мысли Бэра, сведена к одной общей схеме — системе трубок, лежащих одна над другой или заключенных одна в другой. Друг над другом расположены трубки центральной нервной системы, главных сосудов и пищеварительной системы; эта совокупность внутренних трубок окружена общей трубкой сосудистого слоя, дающего начало мышцам туловища, а на поверхности она одета кожной трубкой. Указанное взаимное расположение частей и органов, складывающееся у позвоночных в процессе эмбрионального развития, дало Бэру основание выделить позвоночных животных в самостоятельный тип, противопоставив ему остальные три типа животного царства — удлиненный (или членистый), массивный и периферический (или лучистый).

Карл Максимович Бэр. 1792–1876.
Из сравнения способов развития животных, принадлежащих к разным типам, Бэр вывел заключение, что «каждый главный тип следует особому плану развития». Обнаружение связи между основными признаками типа и присущим каждому типу планом развития Бэр облек в форму следующего положения: «План развития есть не что иное, как становящийся тип, и тип есть результат плана развития. Именно поэтому, — утверждал Бэр, — тип можно познать в полноте только из способа его развития. Этот последний и выявляет различия в зародышах, первоначально сходных в своих существенных чертах».
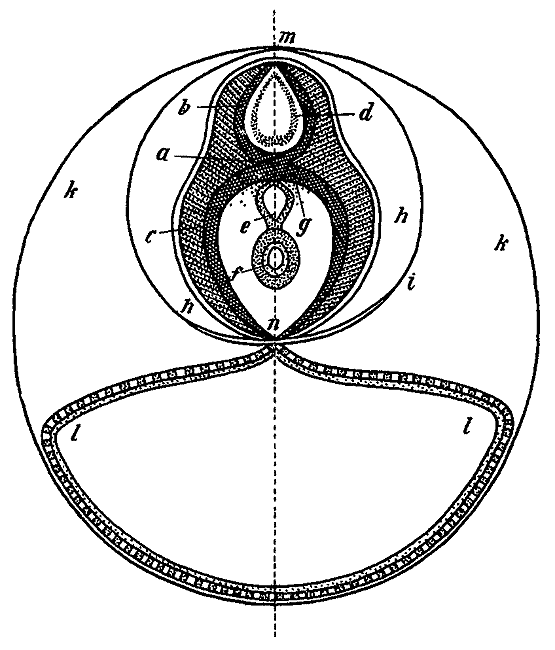
Идеальный вертикальный разрез через эмбрион позвоночного по Бэру. 1828.
Кроме признаков типа, т. е. характерного взаимного расположения органов, Бэр различал в организации животных «степень образования животного тела», т. е. уровень разнородности (гетерогенности) частей, или степень гистологического и морфологического обособления. Например, насекомые имеют более гетерогенное строение, т. е. более высокую «степень образования тела», чем рыбы, хотя последние принадлежат к более высокому, чем насекомые, типу организации. В процессе зародышевого развития признаки типа обнаруживаются очень рано, а «степень образования» повышается лишь постепенно. Так, куриный зародыш, прежде всего, обнаруживает основные признаки позвоночного, к которому он относится, — образование хорды и лежащей над ней нервной трубки. Однако на ранних стадиях еще нельзя решить, к какому классу позвоночных принадлежит данный зародыш; лишь позднее появляются признаки класса птиц, затем признаки отряда куриных, далее признаки рода и вида, и, наконец, признаки породы и чисто индивидуальные особенности данной курицы. Чем моложе зародыши, относящиеся к разным классам одного типа, тем они более сходны между собой. Соображения о сходстве между зародышами, относящимися к разным классам позвоночных, Бэр высказал в форме четырех фундаментальных положений.
1. «В каждой большой группе общее образуется раньше, чем специальное».
2. «Из всеобщего образуется менее общее и т. д., пока, наконец, не выступает самое специальное».
3. «Каждый эмбрион определенной животной формы вместо того, чтобы проводить через другие определенные формы, напротив, отходит от них».
4. «Эмбрион высшей формы никогда не походит на другую животную форму, но только на ее эмбрионы».
Эту последнюю закономерность, ссылаясь на Бэра, использовал Ч. Дарвин в качестве одного из доказательств эволюции и дал ей название «закона зародышевого сходства».
Замечательным признаком зародышевого сходства является наличие жаберных щелей у зародышей всех позвоночных, в том числе и наземных — рептилий, птиц и млекопитающих, включая человека. Жаберные щели у высших позвоночных (млекопитающих и птиц) впервые обнаружил Г. Ратке; вскоре это открытие подтвердил и детализировал К.М. Бэр, изучавший жаберные щели у зародышей человека, собаки и ящерицы. Прижизненные наблюдения над кровообращением у зародышей ящерицы привели Бэра к выводу, что у рептилий имеется одновременно пять пар жаберных дуг; у зародышей птиц и млекопитающих Бэру не удалось одновременно видеть более четырех пар дуг. Он сделал заключение, что у высших позвоночных жаберные дуги появляются и исчезают в известной последовательности.
Топографическая теория зародышевых листков.
Одно из центральных положений эмбриологии начала XIX в. (намеченное Вольфом, твердо установленное Пандером и детально разработанное Бэром) состоит в утверждении, что ранний зародыш образован лежащими друг поверх друга слоями или зародышевыми листками. Предшественники Бэра (Вольф и Пандер) констатировали этот факт только применительно к куриному зародышу, а Бэр распространил учение о зародышевых листках на всех позвоночных.
Стремление применить теорию зародышевых листков к беспозвоночным было характерно для работ Г. Ратке, прежде всего для его исследования, посвященного развитию речного рака (1829). Позднее Ратке коснулся эмбрионального развития морских ракообразных (веслоногих, равноногих, бокоплавов и десятиногих) и скорпиона.
Вслед за Пандером и Бэром, Ратке говорил применительно к зародышам членистоногих о зачатковом диске, или бластодерме, о первичной полоске и о двух зародышевых листках (серозном и слизистом), на которые разделяется бластодерма.
Наряду со сторонниками теория зародышевых листков с самого начала встретилась и с решительными противниками. Наиболее активным из них был К. Рейхерт. Его взгляды, основанные на неточных наблюдениях ¿развития амфибий и на туманных априорных соображениях, бесспорно являлись шагом назад по сравнению с представлениями Пандера и Бэра. Рейхерт, по сути дела, отрицал какую бы то ни было гомологию в развитии амфибий и птиц. Соображения Рейхерта встретили резко отрицательное отношение его современников — Бэра, Ратке, Ремака, Фогта и др. — Э. Геккель впоследствии отмечал, что Рейхерт в своих многочисленных «писаниях… отверг теорию зародышевых листков и связанные с нею наиболее существенные принципы зоогенеза и пытался поставить на их место хаотический конгломерат фантастических выдумок, не заслуживающих даже названия научной гипотезы, а тем более — теории».
В начале 50-х годов учение о зародышевых листках как основе для сравнения эмбриональных стадий различных позвоночных возродилось в работах Р. Ремака, установившего клеточное строение зародышевых листков в зародышевом диске птиц и в яйце амфибий. У этих двух классов позвоночных Ремак различал вполне соответствующие друг другу по значению зародышевые листки: верхний (чувствительный), средний (двигательно-герминативный) и нижний (трофический, или железистый). Критика воззрений Рейхерта, которой Ремак уделил в своем труде о развитии позвоночных много внимания, была направлена в защиту представлений о соответствии зародышевых листков у разных представителей позвоночных.
Первые исследования дробления.
Во втором томе «Истории развития животных» (1837) Бэра помимо сведений о формировании яйца в яичнике курицы и о строении отложенного яйца, а также детального описания формирования отдельных органов и систем органов куриного зародыша, изложены результаты наблюдений над развитием млекопитающих, рептилий, амфибий и рыб. Наибольшее значение для последующего прогресса эмбриологии имело исследование Бэра, касающееся ранних стадий развития амфибий и, в частности, процессов дробления яиц. Дробление яиц лягушки примерно за десять лет до того наблюдали французские исследователи Прево и Дюма, однако существо этого процесса осталось для них непонятным. По меткому выражению Бэра, Прево и Дюма «в буквальном смысле слова остались на поверхности явления», так как они описали в яйце только сеть поверхностных борозд, не подозревая, что эти борозды проникают в глубь яйца и разделяют его на обособленные части.
Через два года после Прево и Дюма итальянский эмбриолог Моро Рускони опубликовал на французском языке работу «Развитие бурой лягушки» (1826), где описал полное разделение яйца бороздами дробления. Рускони позднее писал: «Мы нашли…, что все вещество, из которого построен зачаток, делится сначала на две, затем на четыре части, а последние делятся и подразделяются на другие, все более мелкие». Из рисунков Рускони видно, что он действительно наблюдал полное неравномерное дробление яйца лягушки, равно как и последующие этапы развития — гаструляцию и нейруляцию. В упомянутом уже сочинении «О происхождении яйца млекопитающих и человека» К.М. Бэр, описывая развивающиеся яйца собаки, извлеченные из яйцеводов, отметил, что «шарик под микроскопом обладает не гладкой, а зернистой поверхностью, он весь состоит из плотно уложенных зерен». На это наблюдение Бэра и иллюстрирующий его рисунок позднее обратил внимание Т. Бишоф и в вышедшей в 1842 г. монографии «История развития яйца кролика» сделал вывод: «Приложенное изображение не позволяет сомневаться, что Бэр с определенностью установил разделение желтка на шарики… Так как он использовал слишком слабое увеличение, то и не обратил внимания на природу установленного им внешнего вида яйца». Почти одновременно с Бэром о дроблении яйца на примере медицинской пиявки писал Эрн. Вебер. На ранних стадиях развития Вебер обнаружил в яйце пиявки две зоны — наружную, из более крупных, неправильной формы клеток, и центральную, из многочисленных мелких клеток. Таким образом, Вебер видел в яйце пиявки макро- и микромеры, которые вскоре (1844) значительно более точно описал у другого вида пиявки профессор Дерптского университета А. Трубе.
В 1834 г. Бэр опубликовал работу о развитии яйца лягушки, в которой подробно описал ранние этапы этого процесса. Он окончательно убедился, что щели, видимые на поверхности желтка, представляют границы делений, которые испытывает весь желточный шар. Бэр ввел для борозд дробления обозначения, вошедшие с тех пор в эмбриологическую терминологию: борозды, соединяющие полюсы яйца, он назвал меридиональными, а борозды, пересекающие ось яйца под прямым углом, — экваториальными, или параллельными.
Общие соображения о значении процесса дробления яйца Бэр высказал в следующей форме: «Исходный индивидуум, яйцо, разделяется на бесчисленное количество индивидуальностей, каждая из которых имеет ничтожное значение и оказывается лишь элементарной составной частью нового индивидуума; жизненный процесс растворяет исходную индивидуальность, которая, однако, совсем не разрушается, так как из ее обломков возникает новый индивидуум». В этих словах содержится выражение того единства целостности и расчлененности многоклеточного организма, которое было понято значительно позднее, после окончательного торжества клеточной теории.
Представление, что комочки, или шарики, на которые распадается дробящееся яйцо, являются клетками, сложилось далеко не сразу. Одним из первых к этой мысли пришел Бергман, изучавший дробление яйца лягушки. Он писал: «Желточные комочки, снабженные своеобразным светлым пятном, из которого состоит весь желток (дробящееся яйцо. — Авт.) по своим составным частям должны бесспорно считаться идентичными клеткам, из которых построен зародыш». Бергман сопоставлял дробление яйца с делением растительных клеток и возражал Шванну, предполагавшему, что во время дробления яйца внутри желтка возникают две клетки, в каждой из которых вновь образуются две клетки и т. д.
Еще более необоснованными были представления Рейхерта, сравнивавшего процесс дробления с последовательно совершающимся родовым актом многократно вложенных друг в друга клеток, и Бэрри, считавшего, что ядрышко («зародышевое пятно») яйца наполнено зачатками клеток, две из которых усиленно развиваются и заполняют ядро («зародышевый пузырек»); внутри этих клеток повторяется тот же измышленный Бэрри процесс эндогенного клеткообразования. Бишоф, критиковавший упомянутые фантазии Бэрри, в описании дробления яиц млекопитающих был близок к истине, хотя и не признавал тождества между бластомерами дробящегося яйца и клетками, так как не мог обнаружить у бластомеров оболочки, считавшейся тогда обязательным атрибутом клетки. «На этом основании, — писал Бишоф, — я не могу называть клетками шары, возникающие при делении желтка кроличьего яйца, и во всем этом явлении не могу признать процесс клеткообразования».
Относительно природы светлых пятен в недробившемся яйце и затем в образовавшихся из него шарах дробления, как ранее называли бластомеры, эмбриологи долгое время не могли прийти к согласию. Путаные и противоречивые суждения на эту тему можно найти и в разных работах одного и того же автора, например у Келликера и Бишофа. В исследовании развития круглых червей Келликер описывал исчезновение зародышевого пузырька вместе с зародышевым пятном после оплодотворения, вслед за чем, по его мнению, из желтка образуется ядро, вокруг которого обособляется первая эмбриональная клетка. Она, как считал Келликер, «порождает» две дочерние клетки, для чего ее ядро делится, и вокруг каждой его половины образуется новая клетка, а первичная клетка растворяется. Такое повторное эндогенное клеткообразование продолжается, по словам Келликера, и позднее. Бишоф сначала критически относился к утверждениям Келликера и Бэрри, как можно судить по монографиям о развитии кролика (1842) и о развитии собаки (1845), однако почти 20 лет спустя он вернулся к ошибочным взглядам Бэрри и писал, что «зародышевый пузырек является единственной… полноценной клеткой, которая обнаруживается во всей истории развития яйца… Я рассматриваю, — продолжал он, — зародышевый пузырек как центральное образование клеточной природы, вокруг которого развиваются все прочие части яйца».
Изучением дробления яиц беспозвоночных, главным образом моллюсков, в конце 30-х и начале 40-х годов занимались С. Ловен (1839), М. Саре (1840), а также П. ван Бенеден и Ш. Виндишман (1841). Они оставили точные описания ранних этапов развития, не затрагивая внутренних процессов, совершающихся в шарах дробления, и их судьбы, в последующем развитии.
Существенный прогресс в понимании явлений дробления яйца знаменует собой монография Келликера «История развития головоногих» (1844). Правда, и здесь он использовал без нужды усложненную терминологию и различал «первичные клетки», т. е. шары дробления, и «вторичные», или «шванновские», клетки, однако ясно видел генетическую преемственность описываемых частей зародыша. «Все эти… элементы животных, — писал Келликер, — возникают в непрерывном ряду один от другого».
Одновременно (1844) вышло в свет тщательное исследование эмбрионального развития хоботной пиявки Clepsine, выполненное А. Грубе. В этой работе описано дробление яиц, причем Грубе удалось различить макромеры («сегменты желтка») и микромеры («шарики, образующие стенку тела»). Микромеры, по наблюдениям Грубе, возникают как в результате отшнурования от макромеров, так и путем размножения. Взаимные перемещения бластомеров изображены так, что очень напоминают позднейшие описания эпиболической гаструляции. Грубе обнаружил также зародышевые полоски, начинающиеся каждая тремя концевыми клетками; можно считать, что он открыл явление телобластического развития.
Явления, происходящие в ядрах при дроблении яиц, долго оставались неясными. Длительное время господствовало убеждение, что зародышевый пузырек, т. е. ядро ооцита, при созревании исчезает и появляется снова перед началом дробления в зрелом яйце. Значение яйцевого ядра, которое чешский биолог Ян Пуркине описал в работе «История яйца птиц до насиживания» (1825) (Бэр подтвердил его существование в яйцах млекопитающих и насекомых), по мнению Бэра, очень велико. В сочинении «Об образовании яйца у млекопитающих и человека» Бэр высказал предположение, что «пузырек Пуркине есть действенная часть яйца, посредством которой женский принцип обнаруживает свою силу, подобно тому, как мужской принцип присущ мужскому семени». Говоря об исчезновении пузырька Пуркине, которое Бэр обозначал как «выталкивание» и «растворение», он заметил, что «после оплодотворения бластодерма развивается в том месте, где излилось жидкое содержимое пузырька». Много позднее в эти описания Бэра было внесено новое содержание: «выталкивание» пузырька Пуркине соответствует отделению полярного тельца, а «растворение» пузырька обозначает исчезновение ядерной оболочки в метафазе деления созревания.
Поведение яйцевого ядра до и во время дробления Бэр впоследствии (1845) проследил на яйцах морского ежа. Он видел на живом объекте, как после искусственного осеменения, но еще до начала дробления, внутри яйца обнаруживается светлое сияние, которое вскоре исчезает. Слегка сдавливая такие яйца, Бэр наблюдал в каждом из них вытянутый пузырек или два лежащих рядом пузырька. Установив направление длинной оси этого светлого участка, Бэр отметил, что после деления «желтка» на две части центры половин располагаются на той же оси. Из этих наблюдений он сделал вывод, что «процессы во внутреннем участке яйца предшествуют делению желтка и обусловливают его». Изучение последующих стадий дробления привело Бэра к выводу, что внешне заметным проявлениям дробления всегда предшествует деление внутреннего прозрачного ядра.
Процесс деления ядер в дробящемся яйце Бэр описал с той точностью, какая возможна при наблюдении на нефиксированных и неокрашенных яйцах: «Ядро, имеющее до того шарообразную форму, довольно быстро удлиняется и одновременно с обеих его сторон появляются отростки или выпуклости; эти концы ядра набухают, а середина его истончается и скоро совсем разрывается, так что обнаруживаются два кометообразных ядра с направленными друг к другу хвостами. Хвостовой придаток очень быстро втягивается внутрь своей шаровидной массой, и тогда имеются налицо два ядра… Только после того, как оба новых ядра отделятся одно от другого, начинается перешнуровка яйца, вследствие чего оно разделяется на две прилежащих друг к другу половины». Бэр заметил, что ему не приходилось констатировать момент, когда ядро совершенно отсутствует. Он был уверен, что в непрозрачных яйцах, например у лягушки, зародышевый пузырек удастся обнаружить и в том периоде, когда, как многие полагают, он полностью исчезает. Вплоть до поздних стадий дробления и даже у свободно плавающего зародыша морского ежа Бэр видел сходные фигуры деления клеток и считал весьма вероятным, что все вновь возникающие ядра происходят из исходного яйцевого ядра.
Основные итоги эмбриологических исследований в первой половине XIX века.
Исследования эмбриологов рассматриваемого периода сделались источником обобщений, оказавших существенное влияние на развитие различных областей биологии в последующее время. Первой половиной XIX в. датируется введение в эмбриологию сравнительного метода, позволившего не только сопоставить ход зародышевого развития представителей разных классов позвоночных, но и сравнить морфологические закономерности онтогенеза позвоночных и беспозвоночных. При этом были обнаружены как признаки различия в эмбриональном развитии животных разных типов и классов, подкреплявшие сложившиеся у сравнительных анатомов представления о типах организации, так и признаки сходства, наличие которых позволило сформулировать справедливую для всех многоклеточных животных теорию зародышевых листков. Эта теория, ставшая во второй половине XIX в. одной из основ теории Дарвина, выросла на базе топографической теории зародышевых листков, созданной в первой половине века.
Другим важным итогом изучения эмбрионального развития в рассматриваемом периоде было описание процессов дробления яйца, что имело большое значение для доказательства универсальности клеточной теории, распространенной тем самым с дифференцированных тканей взрослых организмов на яйцо и первые стадии эмбрионального развития.
Таким образом, в первой половине XIX в. были намечены связи первоначально изолированных областей биологии, в частности, эмбриологии с систематикой (учение о типах строения и развития), с ранним эволюционным учением (единство закономерностей эмбрионального развития различных животных) и цитологией (учением о клеточном строении животных о размножении клеток делением).
Изучение оплодотворения.
После того как К. Бэр открыл яйцо у млекопитающих (1827), окончательно сложилось представление о том, что именно яйцо является материальным началом образования зародыша в женском организме. В то же время вопрос о роли мужского начала в образовании зародыша по-прежнему оставался спорным. Продолжало господствовать представление о том, что не сперматозоиды, а семенная жидкость несет оплодотворяющее начало. Сперматозоидам отводили второстепенную, вспомогательную роль в процессе оплодотворения, считая, что, они лишь, смешивают семенную жидкость и переносят ее к яйцу, хотя еще исследования Якоби, Спалланцани, Прево и Дюма убедительно показали непосредственное участие сперматозоидов в процессе оплодотворения.
К. Бэр в «Комментарии» к сочинению «Об образовании яйца млекопитающих и человека» (1828), подводя итоги изучению проблем зарождения животных, предвосхитил правильное понимание сущности процесса оплодотворения, правда, в форме натурфилософского обобщения. Он писал, что зародыш возникает в результате соединения между собой шара и линии, вещества и движения, понимая под шаром и веществом яйцо, а под линией и движением — сперматозоид. Поясняя это обобщение, Бэр писал, что сперматозоид, «победивший в неустанной борьбе», стремится к «соединению с яйцом».
Однако это правильное понимание содержания процесса оплодотворения не нашло признания, так как в то время не были установлены происхождение и природа сперматозоидов. Их все еще считали маленькими животными, паразитирующими в семенной жидкости, и отождествляли с инфузориями и другими микроскопическим организмами, обитающими в различных жидкостях. Также не был еще установлен факт проникновения сперматозоида внутрь яйца и соединения с ним. Спалланцани, ошибочно истолковав результаты своих исследований, тоже считал, что не сперматозоиды, а семенная жидкость оплодотворяет яйцо. Да и Бэр в «Истории развития животных» (т. 2, 1837), писал, что яйца оплодотворяют «не семенные зверьки, а жидкие или даже более тонкие части спермы».

Ладзаро Спалланцани. 1729–1799.
В последующие годы стали появляться одна за другой работы, посвященные изучению происхождения сперматозоидов. В 1838 г. Р. Вагнер опубликовал статью «Происхождение семенных зверьков у птиц», а Ф. Дюжарден — об изучении морфологии сперматозоидов тритона. В 1841 г. вышла работа М. Пельтье об образовании сперматозоидов в семеннике лягушки. В этих трудах подробно освещались процессы образования и развития сперматозоидов в половых железах амфибий и птиц, а также доказывалось, что сперматозоиды являются производными внутреннего слоя извитых канальцев семенников и образуются из округлых семенных телец (сперматогоний, по современной терминологии), заложенных в этом слое.
Обстоятельное исследование К. Лаллемана «Наблюдения над происхождением и способом развития сперматозоидов» (1841), проведенное на большом фактическом материале, выявило картину образования и развития сперматозоидов в семенниках различных видов животных — от моллюсков до человека. Автор пришел к выводу, что сперматозоиды, являясь продуктами жизнедеятельности организма, несут оплодотворяющее начало. В том же году Лаллеман в другой работе «Наблюдение над ролью сперматозоидов при зарождении» убедительно показал, что семенная жидкость не может оплодотворять яйцо и тем более передавать зародышу наследственные задатки отца. Он решительно утверждал, что зародыш может образоваться лишь в результате слияния яйца и сперматозоида. Наконец, в 1841 и 1847 гг. появились классические исследования А. Келликера, раскрывшие детальную картину сперматогенеза у животных различных классов. Результаты всех этих исследований послужили толчком к изучению роли сперматозоидов в процессе оплодотворения яйца.
В 1842 г. появилась монография Ф.А. Пуше «Положительная теория оплодотворения у млекопитающих», а в 1847 г. — его вторая обширная монография «Положительная теория спонтанной овуляции и оплодотворения млекопитающих и человека», в которых на основании обобщения результатов изучения оплодотворения у различных видов животных, проведенного многими учеными и самим автором, Пуше пришел к выводу, что сперматозоиды, проникнув в матку, а затем в яйцеводы, оплодотворяют яйца.
Вопреки представлению о том, что выделение яиц из фолликулов (овуляция) происходит у млекопитающих в результате полового акта, а оплодотворение яиц осуществляется непосредственно в яичниках проникшей в них семенной жидкостью, Пуше убедительно доказал, что овуляция у подавляющего большинства млекопитающих наступает спонтанно в периоды половой активности самок, независимо от полового акта и оплодотворения, которое происходит после выделения яиц и продвижения их по яйцеводам к матке. Это положение было установлено также исследованиями Рациборского и Коста, а затем Бишофа, который посвятил этому вопросу книгу «Доказательство независимых от оплодотворения периодических созреваний и выхода яиц у млекопитающих и человека, как первого условия их размножения» (1844). После длительной дискуссии правильность этого положения была окончательно подтверждена.
Однако и после этих исследований правильное представление о сущности процесса оплодотворения часто оспаривалось, так как еще не было окончательно доказано проникновение сперматозоидов в яйца. Напротив, многочисленные исследования Бишофа показывали, что сперматозоиды, накапливаясь в большом количестве в наружной прозрачной оболочке яйца, приходят контакт с его желчной оболочкой, не проникая внутрь яйца. Поэтому Келликер предположил, что сперматозоиды выделяют особое вещество, которое при контакте последних с желточной оболочкой оказывает на яйцо оплодотворяющее действие. Это положение получило дальнейшее развитие в трудах Бишофа (1847), который считал действие оплодотворяющего вещества сперматозоидов на яйцо каталитическим.
Созданная таким путем Келликером и Бишофом контактно-каталитическая теория оплодотворения получила широкое распространение. Однако она вызывала существенные возражения, поскольку базировалась лишь па предположении о выделении сперматозоидами оплодотворяющего вещества, фактическое существование которого не было установлено. Эта теория не раскрывала также сущности процессов воздействия оплодотворяющего вещества на яйцо, а главное, она не объяснила, каким образом оно может передавать наследственные задатки самца образующемуся из яйца зародышу. Она находилась в явном противоречии с накапливавшимися в 40-х годах данными о проникновении сперматозоидов в яйца различных видов животных. Еще в 1843 г. были опубликованы одно за другим сообщения Бэрри под названием «Сперматозоиды, наблюдаемые в яйцах млекопитающих» и «Сперматозоиды, вторично наблюдаемые в яйце». В более поздней работе (1850) этот автор описал свои наблюдения за процессом внедрения сперматозоида в яйцо кролика и утверждал, что после объединения мужского и женского зарождающего начала происходит дробление яйца на две части.
Дж. Ньюпорт на основании многочисленных и тщательно проведенных опытов в начале 50-х годов XIX в. установил, что сперматозоиды после контакта с яйцом при активном движении хвоста в течение нескольких секунд проникают в него своей головкой, которая находится всегда под острым углом к оболочке яйца. Проникновение происходило в любом месте оболочки яйца. Тотчас после этого автор наблюдал разрушение структуры сперматозоида и его слияние с яйцом. Основываясь на полученных им результатах, Ньюпорт решительно возражал против контактно-каталитической теории оплодотворения.
В монографии «О размножении аскарид» (1851–1852) X. Нельсон также описывал, что сперматозоиды при встрече в яйцеводе с яйцом прилипают к его оболочке по всей поверхности и постепенно внедряются в желток яйца. Здесь они набухают, становятся прозрачными и постепенно распадаются. Одновременно разрушается и зародышевой пузырек (ядро) яйца, после чего желток делится, и образуются две клетки.
В 1853 г. Ф. Кебер описал проникновение сперматозоидов в яйцо как сложный физиологический процесс взаимодействия мужской и женской половых клеток. По его наблюдениям, сперматозоид, проникнув в яйцо, сначала находится у края желтка, а затем, продвигаясь глубже, приближается к зародышевому пузырьку и распадается на большое число угловатых, неравных по величине зернышек.
Ученые того времени единодушно отмечали, что после проникновения сперматозоидов в яйцо их едва можно обнаружить под микроскопом. С еще большими трудностями они столкнулись при наблюдении процессов превращения сперматозоидов в яйце. Таким образом, точно проследить изменения в структуре яйца после проникновения в него сперматозоида не представлялось возможным. Дюма в 1827 г. справедливо писал, что при изучении процессов оплодотворения у животных оставался невыясненным важнейший вопрос — проникает ли сперматозоид в яйцо и какова его роль в процессе оплодотворения.
Трудности изучения сложных процессов оплодотворения привели к тому, что многие ученые начали сомневаться в правильности описанных наблюдений и отказывались признать новую теорию оплодотворения, основывавшуюся на фактах проникновения сперматозоидов в яйца и слияния с ними. В 50-х годах появились даже публикации, в которых эти факты оспаривались. Многие считали, что при несовершенстве микроскопической техники Бэрри, Ньюпорт, Нельсон и другие исследователи, как писал в 1854 г. Бишоф, могли принять за сперматозоиды (с хвостом и без хвоста) какую-нибудь структуру в результате неправильного отражения света, или же принять за сперматозоиды остатки эпителия. Т. Хесслинг в том же году утверждал, что описанные структурные образования в яйце не имеют никакого отношения к оплодотворению. Годом раньше О. Функе априорно утверждал, что проникновение сперматозоидов в яйцо вообще не представляется возможным, а поэтому его и нельзя доказать.
Однако даже Бишоф, опубликовавший в 1854 г. книгу под названием «Опровержение утверждений доктора Кебера, обнаружившего у моллюсков, и доктора Нельсона — у аскарид, проникновение сперматозоидов в яйцо», после тщательной проверки фактов в том же году выпустил другую книгу под названием: «Подтверждение наблюдений доктора Ньюпорта при исследовании лягушек и доктора Бэрри при исследовании кроликов о проникновении сперматозоидов в яйцо». По мнению Бишофа, не может быть никакого сомнения в том, что сперматозоиды действительно проникают в яйцо лягушки и кролика. Он отметил также, что Бэрри первый увидел сперматозоид в яйце и что ему принадлежит честь этого открытия.
В том же году появились новые монографии Ф. Кебера и Р. Вагнера, а затем в 1855 г. обстоятельное экспериментальное исследование Г. Мейснера. В них убедительно доказывалось проникновение сперматозоидов в яйца, описывались их последующие превращения и протекающие одновременно сложные изменения в структуре яиц, за которыми происходили последовательные дробления яйца.
Таким образом, в 50-х годах прошлого столетия факт проникновения сперматозоидов в яйцо был окончательно доказан. Было также дано первоначальное описание картины превращений сперматозоидов и яиц при их соединении. Однако ввиду несовершенства микроскопической техники ученые того времени не могли всесторонне и достаточно детально изучить эти превращения и понять до конца сущность процесса оплодотворения. Очень важно, тем не менее, что они связывали процессы превращений, происходившие в сперматозоидах и яйцах при их слиянии, с оплодотворением и последующим дроблением яиц. Следовательно, уже в 50-х годах ученые подошли к правильному пониманию процесса оплодотворения и подготовили почву для выдающихся открытий в этой области, которые были сделаны в последующие годы благодаря применению более совершенной микроскопической техники с ее методами фиксации и окраски исследуемого материала.

