Книга: ДНК. История генетической революции
Назад: Глава 3 Читаем код: воплощение ДНК
Дальше: Глава 5 Биотехнология: ДНК, доллары и биопрепараты
Глава 4
Подобные Богу: работа над молекулой ДНК
Молекула ДНК невероятно длинная. В любой хромосоме присутствует всего одна непрерывная двойная спираль ДНК. Когда требуется «популярно» объяснить масштабность этой молекулы, нуклеотидную наполненность ДНК сравнивают с количеством записей в телефонном справочнике Нью-Йорка или с длиной Дуная. Мне такие сравнения ни о чем не говорят – я не знаю, сколько номеров в телефонной книге Нью-Йорка, а Дунай ассоциируется у меня скорее с вальсами Штрауса, а не с какой-то линейной дистанцией.
Все человеческие хромосомы (кроме половых: X и Y) нумеруются в зависимости от размера. Хромосома 1 – самая крупная, а хромосомы 21 и 22 – мельчайшие. На хромосому 1 в каждой клетке приходится 8 % ДНК, примерно четверть миллиарда пар оснований. В хромосомах 21 и 22 содержится соответственно 48 и 51 миллион пар оснований. Даже в самых маленьких молекулах ДНК (у вирусов маленьких размеров) как минимум несколько тысяч пар оснований.
На заре молекулярной биологии огромные размеры молекулы ДНК представляли серьезную проблему. Чтобы разобраться с тем или иным геном, то есть конкретным отрезком ДНК, нужно было каким-то образом отделить его от всей остальной ДНК, которая простирается по обе стороны от этого гена. Но этим дело не ограничивалось: ген нужно было не только выделить, но и, так сказать, увеличить: получить настолько крупный фрагмент гена, чтобы с ним можно было работать. В сущности, нам требовался инструмент для молекулярного редактирования – пара молекулярных ножниц, которые позволяли бы кроить текст ДНК, получая удобоваримые фрагменты. Нужен был молекулярный клей, чтобы соединять полученные фрагменты, и, наконец, молекулярная «копировальная машина» для увеличения нарезанных и выделенных нами молекулярных фрагментов. Мы хотели делать примерно то, что сегодня позволяет делать современный текстовый редактор: вырезать, копировать и вставлять кусочки ДНК.
Разработка базовых инструментов для таких процедур казалась отчаянно сложной задачей даже после расшифровки генетического кода. Однако после ряда открытий, сделанных в конце 60-х – начале 70-х годов, для нас словно «звезды сошлись»: в 1973 году появилась так называемая технология рекомбинантных ДНК, что дало возможность редактировать ДНК. Это был не просто прорыв в методах молекулярной биологии. Ученые разом обрели инструмент для адаптации молекул ДНК к потребностям исследователя путем создания таких ДНК, которых никогда не существовало в природе. Мы смогли опробовать себя в роли Бога, экспериментируя с молекулярной основой самой жизни. Такая идея многим казалась некорректной. Так, всегда настороженный и остро реагирующий на любые новаторские идеи Джереми Рифкин, которому каждая новая генетическая технология казалась скользкой дорожкой к созданию монстра, наподобие Франкенштейна, очень верно отметил, что «технология рекомбинантной ДНК может поспорить по значимости с приручением огня».
Артур Корнберг первым «создал жизнь» в пробирке. Как мы уже знаем, в 1950-е годы он открыл ДНК-полимеразу, фермент, обеспечивающий репликацию ДНК и выстраивающий комплементарную копию из расплетенной исходной нити. Позже, работая с вирусной ДНК, он наконец смог осуществить репликацию всех 5300 пар оснований ДНК этого вируса. Однако полученный продукт не был «живым»: несмотря на то что последовательность оснований ДНК не отличалась от исходной, молекула была биологически инертна. Чего-то не хватало. Это недостающее звено удалось найти лишь в 1967 году, причем это одновременно сделали Мартин Геллерт из Национальных институтов здравоохранения и Боб Леман из Стэнфорда. Фермент назвали лигазой. Лигаза позволяет склеивать концевые участки молекул ДНК.
Артуру Корнбергу удалось реплицировать вирусную ДНК при помощи ДНК-полимеразы, а добавив фермент лигазу, он сформировал из ДНК непрерывный контур, как это и было устроено в «подопытном вирусе». Теперь «искусственная» вирусная ДНК вела себя точно так же, как и исходная вирусная: обычный вирус размножается в E. coli, и ДНК, выведенная Корнбергом in vitro, вела себя точно так же. Воспользовавшись лишь парой ферментов, простейшими химическими ингредиентами и вирусной ДНК, с которой была снята копия, Корнберг синтезировал биологически активную молекулу. Средства массовой информации тут же сообщили, что Корнберг создал «жизнь в пробирке», а президент Линдон Джонсон назвал этот прорыв «ошеломительным достижением».
Вклад Вернера Арбера в разработку технологии рекомбинантной ДНК, сделанный в 1960-е годы, был не столь предсказуемым. Швейцарский биохимик Вернер Арбер интересовался не грандиозными вопросами о молекулярной природе жизни, а загадочными аспектами эволюции вирусов. Он изучал процесс, в ходе которого некоторые вирусные ДНК деградировали после внедрения в бактериальные клетки. Некоторые клетки-хозяева (но не все – иначе вирусы не могли бы воспроизводиться) распознавали вирусные ДНК как чужеродные тела и избирательно атаковали их. Но как – и почему? Все ДНК в природе – это разновидности одной и той же молекулы, кому бы они ни принадлежали: бактериям, вирусам, растениям или животным. Почему бактерия не атакует собственную ДНК так же, как вирусную?
Первые ответы на поставленные вопросы появились после того, как Арбер открыл новую группу ферментов, расщепляющих ДНК, – так называемые рестриктазы. Они присутствуют в бактериальных клетках и подавляют размножение вирусов, разрезая на фрагменты чужеродную ДНК. Такое разрезание ДНК – это специфическая реакция на конкретные последовательности: фермент разрезает нить ДНК, лишь если обнаружит в ней искомую последовательность. EcoRI была одной из первых открытых рестриктаз – она находит и обрезает нить оснований ГААТТЦ.
Однако почему бактерия при этом не обрезает собственную ДНК везде, где в ней встречается последовательность ГААТТЦ? Здесь Арбер совершил второе великое открытие. Бактерия синтезирует не только рестриктазу, нацеленную на конкретные последовательности, но и второй фермент, химически модифицирующий те самые последовательности в собственной ДНК, как только они ему попадаются. Измененные таким образом последовательности ГААТТЦ, присутствующие в бактериальной ДНК, не привлекают рестриктазу EcoRI, даже когда фермент словно катком проносится по клетке, повсюду разрезая замеченные вирусные ДНК. Защита бактериального генома от собственной рестриктазы осуществляется с помощью метилирования нуклеотидных остатков аденина и цитозина и называется маскированием.
В основе следующего этапа революции в молекулярной биологии, связанной с рекомбинантной ДНК, было изучение развития у бактерий антибиотикорезистентности. В 1960-х годах выяснилось, что у многих бактерий такая резистентность возникает не по стандартной схеме через мутацию бактериального генома, а путем импорта так называемой плазмиды: это небольшие молекулы ДНК, находящиеся внутри бактерии, физически отдельные от геномных хромосом и способные реплицироваться автономно и передаваться потомству вместе с остальным бактериальным геномом при делении клеток. В некоторых случаях бактерии сами могут обмениваться плазмидами, и в таком случае бактерия-получатель приобретает «информационный набор», которого у нее не было «при рождении». Таким образом, плазмиды служат средством горизонтального переноса генов. В передаваемом информационном комплекте часто имеются гены, как раз и обеспечивающие резистентность к антибиотикам. Естественный отбор у бактерий, работающий в направлении антибиотикорезистентности, благоприятствует тем клеткам, у которых имеется плазмидный фактор антибиотикоустойчивости.

Плазмида под электронным микроскопом
Первопроходцем в исследовании плазмид был Стенли Коэн из Стэнфордского университета. Коэн выбрал медицинскую карьеру, поскольку его вдохновил на этот путь школьный учитель биологии. Закончив медицинский университет, он подумывал заняться внутренними болезнями, но забросил эти планы, когда перед ним встал выбор – пойти служить армейским врачом или занять должность в Национальных институтах здравоохранения. Вскоре он осознал, что исследовательская деятельность ему более интересна, чем практическая медицина. Первый серьезный успех ждал его в 1971 году, когда Коэн научился управлять захватом бактериями E. coli плазмид вне пределов клетки. Фактически Стенли Коэн «трансформировал» E. coli подобно тому, как Фред Гриффит сорока годами ранее превратил жизнеспособные пневмококки в нежизнеспособные, «заставив» их поглотить участок ДНК. В опытах Коэна бактерия поглощала плазмиду с генами устойчивости к антибиотикам, и штамм, ранее погибавший от антибиотика, терял восприимчивость к нему. Штамм сохранял устойчивость к антибиотику и в последующих поколениях – копии плазмидной ДНК в целости и сохранности передавались потомкам при делении клеток.
К началу 1970-х годов имелись все составляющие для получения рекомбинантной ДНК. Сначала было нужно разрезать молекулу ДНК при помощи рестриктаз и выделить интересующие нас последовательности (гены), а затем скопировать интересующий нас фрагмент ДНК и вставить плазмиду в бактериальную клетку, как USB-флешку в подготовленный для нее разъем. За этим процессом последует обычное бинарное деление бактерий, и плазмида с выбранным нами фрагментом ДНК будет реплицироваться точно так же, как и собственный генетический материал, унаследованный бактериальной клеткой. Таким образом, после пересадки единственной плазмиды в бактериальную клетку в процессе последующего деления бактерии в огромных количествах будет воспроизводиться выбранная нами последовательность ДНК. Поскольку мы сами способствуем воспроизводству выбранной клетки, то через короткий промежуток времени мы создадим колонию из миллиардов бактериальных особей, а значит, и миллиарды копий интересующего нас фрагмента ДНК. Соответственно, полученная нам колония – это завод по производству ДНК.
Все три операции – вырезание, вставка и копирование – были выполнены в 1972 году в Гонолулу. Это произошло на конференции, посвященной исследованию плазмид. На этой конференции присутствовали Герб Бойер, молодой ученый, недавно получивший пост штатного профессора в Калифорнийском университете в Сан-Франциско, и, что ожидаемо, Стенли Коэн, один из первых исследователей плазмид. Оба они были выходцами с востока США. Бойер происходил из Западной Пенсильвании и в старших классах играл в футбол – был нападающим. Пожалуй, ему очень повезло, что тренер по футболу одновременно был учителем естествознания. Как и Коэн, Бойер являлся представителем нового поколения ученых, воспитанных на идее двойной спирали. Он так увлекался изучением ДНК, что даже назвал своих сиамских котов Уотсон и Крик. Поэтому никто, включая тренера, не удивился, когда по окончании колледжа Бойер решил заняться генетикой бактерий.
Хотя и Коэн, и Бойер в те времена работали на берегах бухты Сан-Франциско, до гавайской конференции они не встречались. Бойер был экспертом по рестриктазам уже тогда, когда о них практически никто еще не слышал; именно он и его коллеги определили последовательность оснований на участке, вырезаемом рестриктазой EcoRI. Вскоре Бойер и Коэн осознали, что в альянсе друг с другом смогут вывести молекулярную биологию на совершенно новый уровень – в мир копирования, вырезанияи вставок. Как-то поздним вечером они зашли в ресторанчик в районе Вайкики и принялись фантазировать о зарождении технологии рекомбинантной ДНК, кратко конспектируя свои идеи прямо на салфетках. Такую форму предвидения будущего окрестили «от солонины к клонированию».
Через несколько месяцев наладилось сотрудничество между лабораториями Бойера (в Сан-Франциско) и Коэна (в 64 километрах к югу от Пало-Альто). Герб Бойер продолжал работать над исследованием рестриктаз, а Стенли Коэн ставил опыты с плазмидами. На их удачу, у Коэна была лаборантка Энни Чанг, которая жила в Сан-Франциско и успешно осуществляла обмен драгоценной информацией о результатах исследований, проходящих в этих лабораториях. На первом этапе ученые решили создать гибрид – рекомбинант, состоящий из двух разных плазмид, каждая из которых была устойчива к конкретному антибиотику. В одной плазмиде имелся ген (участок ДНК), обеспечивавший устойчивость к тетрациклину, а в другой – ген устойчивости к канамицину. (Как вы уже догадываетесь, исходно бактерии с первой плазмидой погибали от канамицина, а бактерии с второй плазмидой – от тетрациклина.) Предполагалось сконструировать единую «суперплазмиду», которая бы обеспечивала устойчивость к обоим антибиотикам.
Сначала при помощи рестриктаз разрезали две неизмененные плазмиды. Затем эти плазмиды смешивались в одной пробирке, куда добавлялся фермент лигаза, которая должна была запустить склеивание обрезанных концевых остатков. Некоторые молекулы в пробирке под действием лигазы просто восстанавливали целостность – то есть склеивались два концевых остатка одной и той же плазмиды. Но иногда лигаза срабатывала так, что в разрезанную плазмиду попадали фрагменты ДНК другой плазмиды – так и получался желаемый гибрид. Когда эта задача была решена, требовалось внедрить все плазмиды в бактерии, и это успешно было проделано с использованием технологий Коэна. Полученные от рекомбинантов колонии выращивались на агаровых пластинах, покрытых одновременно тетрациклином и канамицином. Те плазмиды, которые просто восстановились свою структуру, по-прежнему обеспечивали устойчивость лишь к одному из двух антибиотиков, и, соответственно, бактерии с такими плазмидами не выживали в среде, содержащей два антибиотика. В такой среде могли выжить только бактерии с рекомбинантными плазмидами, сконструированными из двух имевшихся разновидностей ДНК, одна из которых кодировала устойчивость к тетрациклину, а другая – к канамицину.

Герб Бойер и Стенли Коэн – первые в мире генные инженеры
Следующий вызов сложившемуся в обществе укладу заключался в создании гибридной плазмиды с использованием ДНК не бактерий, а иного организма, например человека. В одном из первых успешных экспериментов ген африканских шпорцевых лягушек удалось добавить в плазмиду E. coli и трансплантировать ее в бактерию. Всякий раз при делении клеток в такой бактериальной колонии реплицировался лягушачий фрагмент ДНК. Если не применять сложную молекулярно-биологическую терминологию, а просто описать происходящее, то это выглядит как «клонирование ДНК лягушки». Как стало известно, ДНК млекопитающих также успешно клонируется. Ретроспективный анализ показал, что в этом нет ничего особенно удивительного: любой фрагмент ДНК – это, в конечном счете, просто ДНК, его химические свойства не изменяются в зависимости от источника. Вскоре стало понятно, что протоколы Коэна и Бойера, описывающие клонирование фрагментов ДНК, применимы к ДНК любого организма.
Таким образом, разворачивался уже второй этап молекулярно-биологической революции. На первом этапе мы стремились описать статус и функционал ДНК в клетке, затем, после получения рекомбинантной ДНК, появились реальные инструменты для вмешательства в работу ДНК и манипулирования ею. Был создан плацдарм для стремительного прогресса, а мы примерили на себя роли Творца. Полученные результаты пьянили: открывался огромный потенциал, позволявший глубоко погрузиться в тайны жизни и добиться успеха в борьбе с такими болезнями, как рак. Несмотря на то что работы Коэна и Бойера предоставили нам фантастические научные перспективы, – не открылся ли при этом ящик Пандоры? Не скрывалось ли в молекулярном клонировании какое-то неизвестное зло? Можно ли и далее беззаботно вшивать кусочки человеческой ДНК в E. coli, учитывая, какие огромные колонии этих бактерии обитают в «микробных джунглях» нашего кишечника? Что будет, если в кишечник проникнет видоизмененная бактерия? Короче говоря, можно ли, будучи в здравом уме, заткнуть уши и не слушать скептиков, заявляющих, что на глазах у всего научного мира мы творим бактерий-франкенштейнов?
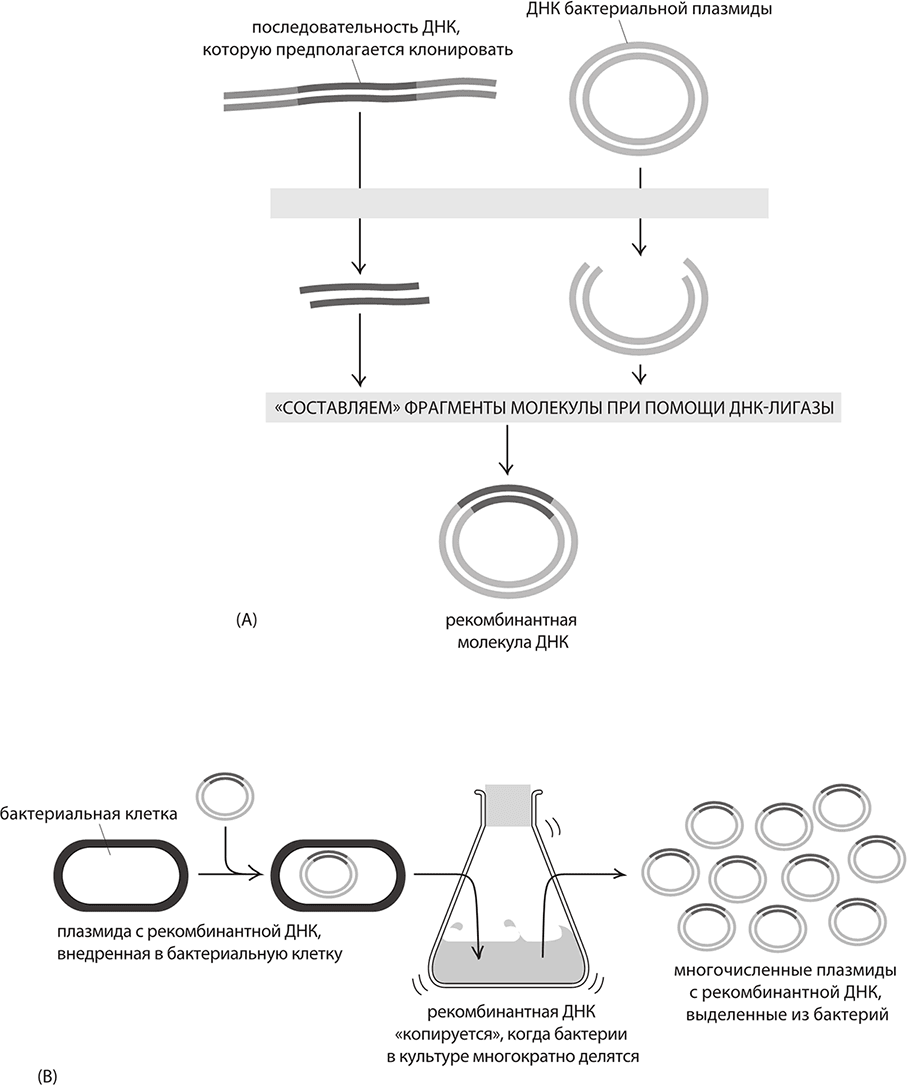
Рекомбинантная ДНК: суть клонирования гена. A. Бактериальные плазмиды оказались идеальным носителем для клонирования ДНК; разрезая одной и той же рестриктазой интересующую нас ДНК и плазмиду, можно вставлять интересующую нас ДНК в плазмиду, как деталь в пазл. B. Внедрив эту рекомбинантную плазмиду в бактерию, можно реплицировать в бактериальной культуре интересующий нас ген – на основе таких приемов развились генная инженерия, секвенирование ДНК и биотехнологическая индустрия

Кишечная палочка E. coli. Представляете, около 10 миллионов этих существ обитает в каждом грамме человеческого кала
В 1961 году удалось выделить обезьяний вирус SV40 (SV означает «обезьяний вирус») из почек макак вида резус, использовавшихся при разработке вакцины против полиомиелита. Хотя и считалось, что этот вирус никак не влияет на мартышек, в организме которых встречается, вскоре дальнейшие эксперименты показали, что вирус может вызывать рак у грызунов, а в определенных лабораторных условиях – даже в человеческих клетках. Поскольку кампания по вакцинации от полиомиелита проводилась с 1955 года и миллионы американских детей уже были заражены этим вирусом, новость действительно была тревожной. Что если, искореняя полиомиелит, мы случайно обрекли целое поколение на риск развития онкологических заболеваний? По-видимому, этого все-таки не случилось: эпидемия рака не разразилась, а SV40 в организме человека, судя по всему, не более патологичен, чем у мартышек. Тем не менее даже тогда, когда SV40 прочно обосновался в молекулярно-биологических лабораториях, оставались сомнения в его безопасности. Меня это особенно волновало, поскольку я к тому времени руководил лабораторией Колд-Спринг-Харбор, где появлялись все новые молодые ученые, работавшие с SV40 и зондировавшие с его помощью генетические основы рака.

Пол Берг и его вирусная «хонда»
Тем временем Пол Берг из Медицинской школы Стэнфордского университета усматривал в вирусе SV40 скорее возможности, чем опасности: он предвидел, что этот вирус удастся использовать для внедрения фрагментов ДНК в клетки млекопитающих. Вирус мог бы работать у млекопитающих как молекулярная система доставки лекарств, именно так, как действовали плазмиды у бактерий в опытах Стенли – Коэна. Однако если Коэн, в сущности, использовал бактерии как копировальные аппараты, позволяющие «увеличить» нужный фрагмент ДНК, то Берг рассматривал SV40 как средство для внедрения корректировочных генов людям, страдающим от генетических заболеваний. Берг опередил свое время. Он стал вдохновителем практики, которая сегодня именуется «генотерапия» и заключается во введении нового генетического материала в организм живого человека с целью сгладить унаследованные генетические расстройства.
Пол Берг был приглашен в Стэнфорд в 1959 году на должность младшего профессора; переход произошел в рамках программы научного обмена, которая также привела в Стэнфорд еще более знаменитого Артура Корнберга из Вашингтонского университета в Сент-Луисе. Действительно, связь между Бергом и Корнбергом прослеживается вплоть до того, что родились они оба в нью-йоркском районе Бруклин и там ходили в один и тот же естественнонаучный клуб для старшеклассников, которым руководила мисс Софи Вольфе. Берг вспоминал: «она умела сделать науку интересной, она помогала нам делиться идеями». На самом деле, это была скорее ее недооценка: из научного клуба мисс Вольфе при старшей школе им. Авраама Линкольна вышло трое нобелевских лауреатов: Артур Корнберг (1959), Пол Берг (1980) и кристаллограф Джером Карле (1985) – и все они отмечали, насколько существенное влияние на них оказала мисс Вольфе.
В то время как Коэн и Бойер, а вслед за ними и другие ученые уточняли детали, связанные с копированием и вставкой молекул ДНК, Берг планировал смелый эксперимент. Он решил проверить, можно ли использовать вирус SV40 с внедренным в него фрагментом чужеродной ДНК для доставки чужого гена в клетку животного. В качестве источника ДНК, не принадлежащей SV40, он воспользовался уже имевшимся под рукой бактериальным вирусом – бактериофагом. Требовалось выяснить, сможет ли молекулярная структура, состоящая из ДНК SV40 и бактериофага, успешно внедриться в животную клетку. Если же она внедрится, на что и надеялся Берг, появится возможность того, что такую систему со временем можно будет использовать для встраивания полезных генов в человеческие клетки.
Летом 1971 года в лаборатории Колд-Спринг-Харбор аспирант Берга выступил с презентацией планируемого эксперимента. Один из ученых, присутствовавших в аудитории, настолько встревожился, что сразу же позвонил Бергу. Он спросил: «А что, если этот механизм сработает прямо наоборот?» Иными словами, что, если вирус SV40 не станет принимать вирусную ДНК и затем встраивать ее в животную клетку, а сам попадет «под власть» ДНК бактериофага, в результате чего ДНК SV40 может внедриться, скажем, в ДНК бактериальной клетки E. coli? Такой сценарий не казался нереалистичным; в конце концов, именно на него и «запрограммированы» многие бактериофаги: они внедряют собственную ДНК в бактериальные клетки. Поскольку E. coli повсеместно распространена, а ее жизненный цикл тесно связан с жизнедеятельностью человека (ведь это один из основных микроорганизмов кишечной микробиоты человека), то благие намерения эксперимента Берга могли породить целые колонии опасных бактерий E. coli, несущих потенциально канцерогенный обезьяний вирус SV40. Берг прислушался к скептическим замечаниям коллеги, но не согласился с ними; он решил отложить эксперименты до тех пор, пока не будет подробно изучена потенциальная канцерогенность SV40 для человека.
Обеспокоенность по поводу смертельной угрозы, связанной с рекомбинантными технологиями, распространялась вслед за новостями об успешных опытах Бойера и Коэна. На научной конференции, посвященной нуклеиновым кислотам, состоявшейся в Нью-Гемпшире в 1973 году, большинством голосов была принята петиция к Национальной академии наук, в которой требовалось незамедлительно проанализировать угрозы, связанные с этой технологией. Через год комитет, назначенный Национальной академией и возглавляемый Полом Бергом, изложил свои выводы в письме, направленном в журнал Science. Я лично подписал это письмо, подписали его и многие другие, в том числе Коэн и Бойер, наиболее активно занимавшиеся соответствующими исследованиями. В этом документе, который позже стал известен под названием «Письмо о моратории», мы призывали «ученых всего мира» добровольно приостановить все исследования рекомбинантной ДНК «до более полной оценки потенциальных угроз, которые могут быть связаны с такими рекомбинантными молекулами ДНК, или до тех пор, пока не будут разработаны адекватные методы для предотвращения их распространения». В письме была важная оговорка о том, что «наши опасения базируются на суждениях о потенциальном, а не доказанном риске, поскольку экспериментальных данных об опасности рекомбинантных молекул ДНК пока недостаточно».
Уже очень скоро я глубоко разочаровался в том, что моя подпись оказалась под «Письмом о моратории». Ведь было очевидно, что молекулярное клонирование сулит миру фантастические блага, а теперь, проделав такую массу работы и оказавшись на заре биологической революции, мы капитулировали. Момент был неоднозначный. Как написал Майкл Роджерс в своем репортаже на эту тему для журнала Rolling Stone, «очевидно, что молекулярные биологи достигли таких экспериментальных рубежей, которые сравнимы лишь с рубежами физики в последние годы перед созданием атомной бомбы». Мы проявили благоразумие или малодушие? Сейчас не могу сказать с уверенностью, но поначалу я склонялся ко второму ответу.
«Конгресс по поводу ящика Пандоры» – так Майкл Роджерс назвал состоявшееся в феврале 1975 года заседание 140 ученых со всего мира, собравшихся в Асиломарском конференц-центре в городе Пасифик Гроув, штат Калифорния. Повестка дня заключалась в том, чтобы раз и навсегда определить, чем для нас является рекомбинантная ДНК: уникальной возможностью или угрозой. Стоит ли объявить наш мораторий бессрочным или двигаться вперед, невзирая на потенциальные риски? Есть еще вариант – дождаться, пока будут разработаны те или иные защитные меры. Пол Берг, возглавлявший оргкомитет, также был номинальным руководителем конференции и должен был справиться с практически неразрешимой задачей: подготовить по итогам встречи общее заключение.
Присутствовавшие журналисты недоуменно чесали в затылке, пока ученые общались на своем «птичьем языке». Там присутствовали и юристы, напоминавшие, что на конференции нужно решить и правовые вопросы: например, если я руковожу лабораторией, где ведутся исследования рекомбинантной ДНК, то несу ли я ответственность за то, что мой работник при этом заболеет раком? Что касается ученых, они по природе и профессиональному складу равнодушно относились к тревожным прогнозам, не подкрепленным доказательствами; поэтому были обоснованные подозрения, что никакого единодушного решения на конференции найти не удастся. Возможно, Берг сомневался не меньше других; так или иначе, он сделал ставку на свободный обмен мнениями, а не на жесткое давление с трибуны. Состоявшиеся дебаты напоминали всеобщую перепалку, и регламент зачастую срывал какой-нибудь оратор, намеренный лишь подолгу и не по делу разглагольствовать о том, какая важная работа ведется в его лаборатории. Мнения чрезвычайно варьировались – от робких («продлить мораторий») до агрессивных («будь проклят этот мораторий, руки прочь от науки»). Я, определенно, тяготел ко второму лагерю. В тот момент мне казалось, что будет просто безответственно тормозить исследования, перестраховываясь от каких-то неизвестных и неоцененных опасностей. В мире были безнадежно больные люди, страдавшие раком и муковисцидозом, – по какому праву можно отобрать у них, возможно, единственную надежду?
Сидней Бреннер, в ту пору обосновавшийся в Кембридже (Великобритания), предъявил кое-какие реальные данные, которых на этой конференции было очень мало. Он собрал колонии штамма E. coli, известные под названием K-12. Это излюбленная «рабочая лошадка» бактериологов, активно используемая в исследованиях, связанных с молекулярным клонированием. Некоторые редкие штаммы E. coli могут вызывать вспышки пищевого отравления. Однако на самом деле абсолютное большинство штаммов E. coli не патологичны, и Бреннер полагал, что и K-12 не исключение. Интересовало его прежде всего не собственное самочувствие, а здоровье K-12: выживут ли эти штаммы вне лаборатории? Он добавлял взвесь бактерий в стакан с молоком (получалось довольно невкусно), после чего залпом его осушал. Далее он наблюдал, что получается на выходе из пищеварительного тракта: удалось ли каким-то клеткам К-12 колонизировать его кишечник? Результат был отрицательным, и это означало, что K-12, процветавшие в чашке Петри, оказывались нежизнеспособны в «естественном мире». Однако ряд ученых скептически относились к этому выводу: даже если бактерии K-12 сами по себе не выживали, это еще не означало, что они не успевали обмениваться плазмидами – или другой генетической информацией – с бактериями-симбионтами кишечника. Следовательно, «генно-инженерный» материал все равно мог проникать в популяцию кишечной микрофлоры. Бреннер отстаивал идею о том, что следует вывести такой штамм К-12, который однозначно будет неспособен выжить вне лаборатории. Этого можно было бы достичь при помощи таких генетических мутаций, которые позволяли бы штамму делиться, лишь если он будет обеспечен конкретными питательными веществами. Естественно, мы подобрали бы такие питательные вещества, которые просто негде взять в естественной среде: полный набор таких веществ должен быть составлен лишь в лаборатории. Модифицированный таким образом штамм К-12 стал бы «безопасной» бактерией, выживающей в контролируемых экспериментах, но обреченной в естественном мире.

Дебаты о ДНК: Максин Сингер, Нортон Зиндер, Сидней Бреннер и Пол Берг за обсуждением проблем на Асиломарской конференции
Под напором Бреннера было принято компромиссное предложение. Естественно, оба радикальных крыла молекулярных биологов были им весьма недовольны, но конференция завершилась согласованным рекомендательным решением о том, что допускается продолжать исследования с использованием аттенуированных, непатогенных бактерий. Также вменялось в обязанность оборудовать дорогие карантинные комплексы для работы с ДНК млекопитающих, обеспечивающие биологическую безопасность. Эти рекомендации заложили основу для руководства, выпущенного год спустя Национальными институтами здравоохранения.
Я покидал конференцию опустошенным, чувствуя отчуждение своих коллег. Стэнли Коэна и Герба Бойера мероприятие также разочаровало; они разделяли мою точку зрения и были разочарованы тем, что многие наши коллеги поступились трезвым научным расчетом, просто чтобы собравшиеся сочли их «хорошими ребятами» (а не фанатами доктора Франкенштейна). В действительности же абсолютное большинство обвинителей никогда не работало с болезнетворными организмами и плохо понимало подоплеку принимаемых ограничений на наши исследования, а исследованиями занимались как раз те, кто отдавал себе отчет в том, что и как он делает. Меня коробило, с какой произвольностью принимались многие соглашения: так, сочли приемлемым работать с ДНК холоднокровных животных, но большинство ученых высказалось за решительный отказ от использования ДНК млекопитающих. По-видимому, они считали, что работать с лягушачьей ДНК безопасно, а с мышиной – опасно. Ошарашенный таким нонсенсом, я «вставил свои пять копеек» юмора: вы что, не знаете, что от прикосновения к лягушке вскакивают бородавки? Но никто не оценил моей шутки.
Благодаря принятому постановлению многие участники Асиломарской конференции решили, что можно будет без проблем вести исследования, связанные с клонированием «безопасных» бактерий. Однако все, взявшиеся за продолжение работы, вскоре попали на зыбкую почву. По логике, пропагандируемой в популярной прессе, если ученые обеспокоены некоторой научной проблемой, то общество тем более должно воспринимать эту проблему с тревогой. В конце концов, тогда все еще продолжалась эпоха американской контркультуры, которая, правда, постепенно шла на спад. Только-только улеглись страсти по поводу вьетнамской войны и политической карьеры Ричарда Никсона. Недоверчивая публика, практически неспособная понять чрезвычайно сложные вопросы естествознания, которые едва начинала постигать сама наука, легко велась на теории о заговоре, учиненном верхушкой власти. Мы, ученые, изрядно удивлялись, что нас причисляют к этой политичекой элите, в рядах которой мы себя и не мыслили. Даже Герб Бойер, образец «хиппующего ученого», обнаружил упоминание о себе в специальном хэллоуинском выпуске Berkeley Barb, андеграундной газеты, выходившей в районе Сан-Франциско, как об одном из десяти «жутчайших персонажей» региона: обычно «такая честь» отводилась лишь коррумпированным политиканам и капиталистам, притесняющим профсоюзы.
Больше всего я боялся, что такая махровая публичная паранойя по поводу вредоносных экспериментов в молекулярной биологии приведет к появлению драконовских законов. Если бы вдруг «что дозволено» и «что не дозволено» в рекомбинантных технологиях оказалось бы прописано в каком-нибудь нормативно-правовом акте, это только навредило бы науке. Планы новых экспериментов потребовалось бы подавать на рассмотрение и утверждение в политизированные экспертные советы, где царила беспросветная бюрократия, неистребимая, как моль в старом бабушкином шкафу. Тем временем, как бы мы ни старались оценить реальный потенциальный риск, присущий нашей работе, нам так и не удавалось справиться с недостатком информации и логическими сложностями «доказательств от противного». Никаких биологических катастроф с рекомбинантной ДНК никогда ранее не возникало, но журналисты пытались перещеголять друг друга, предлагая сценарии таких бедствий один другого мрачнее. Биохимик Леон Геппель, описывая свои впечатления от собрания в Вашингтоне (округ Колумбия) в 1977 году, так резюмировал всю абсурдность тех противоречий, с которыми приходилось иметь дело ученым.
Я чувствовал себя так, словно меня избрали в импровизированный комитет, собранный испанским двором для оценки потенциальных рисков, с которыми могла столкнуться экспедиция Христофора Колумба. Комитет должен был выработать регламент по поводу того, как следует действовать, если Земля окажется плоской, как экипажу безопасно заглянуть за край Земли и т. д.
Однако даже ирония ученых практически ничего не позволяла поделать с мракобесами, ополчившимися против мнимой «прометеевой гордыни» в науке. Одним из таких «крестоносцев» был Альфред Велуччи, мэр Кембриджа, штат Массачусетс. Велуччи заработал политические очки, отстаивая права «простого человека» в борьбе против элитных вузов, расположенных в городе, – речь о Массачусетском технологическом институте и Гарварде. Шумиха по поводу рекомбинантной ДНК стала для него настоящим политическим Эльдорадо. Вот характеристика современника, прекрасно описывающая сложившуюся тогда ситуацию:
Выходит он в своих клюквенно-красной двубортной куртке и черных штанах, под курткой – голубая рубашка в желтую полоску, из-под которой выпирает пивной живот. Карманы его набиты всякой всячиной, а зубы кривые. Таков Эл Велуччи, воплощение американского обывательского недовольства всеми этими учеными и технократами, этими хитрыми гарвардскими «ботанами», возомнившими, что весь мир у них на крючке и что они могут швырнуть его в грязную лужу. И кто же в результате оказывается в луже? Нет, не «яйцеголовые», а неизменно Велуччи и простые работяги, которым только и остается потом самим отмываться от грязи.
Отчего же разгорелся весь сыр-бор? Гарвардские ученые проголосовали за то, чтобы возвести прямо в кампусе карантинный объект, где можно было бы работать с рекомбинантной ДНК в строгом соответствии с регламентом Национальных институтов здравоохранения. Однако Велуччи, предвидя такое развитие событий, заручился поддержкой левой партийной группировки, члены которой, ополчившиеся на исследования ДНК, работали в Гарварде и Массачусетском технологическом институте, и всего за несколько месяцев сумел запретить в Кембридже любые исследования, связанные с рекомбинантной ДНК. В результате наступил короткий, но негативно отразившийся на науке период локальной утечки мозгов: ученые из Гарварда и Кембриджа потянулись в менее политизированную среду. Тем временем Велуччи привыкал к новообретенной славе бдительного защитника социума от науки. В 1977 году он написал президенту Национальной академии наук:
В сегодняшнем номере Boston Herald American (издательская корпорация «Хёрст») есть два репортажа, вызывающих у меня серьезное беспокойство. В Дувре, штат Массачусетс, заметили «странное существо с оранжевыми глазами», а в Холлисе, штат Нью-Гемпшир, мужчина и двое его сыновей повстречали «волосатую девятифутовую тварь».

Слушания в Кембридже, штат Массачусетс. В результате этого процесса в городе были полностью запрещены исследования рекомбинантной ДНК
Я с уважением обращаюсь в вашу авторитетную организацию с просьбой расследовать эти факты. Надеюсь, вы сможете проверить, могут ли эти «странные существа» (если, конечно, они существуют) быть каким-то образом связаны с экспериментами в сфере рекомбинантной ДНК, предпринимаемыми в Новой Англии.
К счастью, несмотря на активное обсуждение в обществе, государственные законы по ограничению исследования рекомбинантной ДНК так и не были приняты. Сенатор от штата Массачусетс Тед Кеннеди вступил в эту дискуссию на самом раннем этапе и организовал слушание по этому вопросу в Сенате всего через месяц после Асиломарской конференции. В 1976 году он обратился к президенту Форду, заявив, что федеральное правительство должно взять под контроль как промышленные, так и академические исследования ДНК. В марте 1977 года я давал объяснения в Законодательном собрании штата Калифорния. На заседании присутствовал губернатор Джерри Браун, и мне удалось пояснить ему лично, что было бы ошибочно принимать к рассмотрению законопроекты против таких исследований за исключением того случая, если ученых в Стэнфорде поразит какая-нибудь неизвестная болезнь. Если люди, непосредственно работающие с рекомбинантной ДНК, останутся совершенно здоровы, то специалистам по законотворчеству лучше сосредоточиться на более реальных общественных опасностях, таких, например, как езда по городу на велосипеде.
По мере того как проводились все новые и новые эксперименты (либо по регламенту Национальных институтов здравоохранения, либо в соответствии с правилами, принятыми в других государствах), становилось все очевиднее, что при экспериментах с рекомбинантной ДНК не возникает никаких франкенштейнов (а уж тем более – полно вам, мистер Велуччи! – «странных существ с оранжевыми глазами»). Уже в 1978 году я смог написать следующее: «Если сравнить ДНК со всеми прочими феноменами, названия которых начинаются на букву d, то она в самом деле совершенно безопасна. Гораздо уместнее поостеречься кинжалов, динамита, собак, дильдрина, диоксина или пьяных водителей (daggers, dynamite, dogs, dieldrin, dioxin, drunken drivers), нежели изображать схемы, достойные Руба Голдберга, измышляя, как наша лабораторная ДНК может привести к вымиранию человечества».
Позже в том же году в Вашингтоне, округ Колумбия, Надзорный комитет Национальных институтов здравоохранения по работе с рекомбинантной ДНК принял гораздо менее жесткий регламент, разрешавший развивать основной массив исследований, связанных с рекомбинантной ДНК, в частности изучать ДНК вирусных онкогенов. В 1979 году Джозеф Калифано, министр здравоохранения и социальных служб США, одобрил эти изменения, на чем и закончился период бессмысленной стагнации исследований рака у млекопитающих.
На практике Асиломарская конференция обернулась удручающе бессмысленным пятилетием, в течение которого тормозились важные исследования, а карьера многих молодых ученых оказалась загублена.
К концу 1970-х годов те проблемы, что были подняты в исходных экспериментах Коэна и Бойера, постепенно решились сами собой. Нам пришлось совершить досадный крюк, но биологи-молекулярщики как минимум продемонстрировали, что готовы нести социальную ответственность за результаты своих экспериментов.
Нельзя сказать, что во второй половине 1970-х годов молекулярная биология оказалась полностью сокрушена противостоянием с политикой; в эти годы были достигнуты некоторые важные успехи, и большинство полученных результатов базировалось на по-прежнему неоднозначной технологии молекулярного клонирования, изобретенной Коэном и Бойером. Важнейший прорыв в данном направлении был связан с открытием методов секвенирования ДНК. Для секвенирования нужно иметь множество образцов интересующего нас отрезка ДНК. Это было неосуществимо (если не считать образцов небольшой вирусной ДНК) до тех пор, пока небыли разработаны технологии молекулярного клонирования. Как мы уже убедились, клонирование, в сущности, заключается в следующем: вставляем интересующий нас фрагмент ДНК в плазмиду, а потом саму плазмиду внедряем в бактерию. Далее мы позволяем бактерии делиться и размножаться и в результате получаем множество копий искомого фрагмента ДНК. Затем этот фрагмент выделяется из бактерий – все, материал для секвенирования готов.
Две технологии секвенирования были разработаны одновременно. Автором одной из них был Уолли Гилберт из Кембриджа, штат Массачусетс (Гарвардский университет), автором другой – Фред Сенгер из британского Кембриджа. Уолли Гилберт заинтересовался секвенированием ДНК после того, как смог выделить репрессорный белок из регуляторной системы гена β-галактозидазы у бактерии E. coli. Как мы уже знаем, он продемонстрировал, что при встраивании нужного гена в хромосомную ДНК хозяина нужно позаботиться о том, чтобы сайт интеграции не находился внутри гена, кодирующего важную клеточную функцию. Кроме того, для обеспечения эффективной экспрессии его помещают под контроль регулируемого промотора.
Для интеграции в нужный сайт вводимый ген должен содержать нуклеотидную последовательность длиной не менее 50 нуклеотидов, сходную с таковой в хромосомной ДНК, в пределах которых и должен произойти физический обмен (рекомбинация) между двумя молекулами ДНК. Далее он решил выяснить, какова последовательность оснований на этом отрезке ДНК. Найти такой способ ему посчастливилось благодаря встрече с блестящим советским химиком Андреем Дарьевичем Мирзабековым. При помощи мощных химических реактивов Уолли Гилберту удалось разделить цепочки ДНК именно на нужных участках, специфичных к конкретным основаниям.
Уолли Гилберт оканчивал школу в Вашингтоне, округ Колумбия, и даже сбегал с уроков, чтобы почитать книги по физике в библиотеке Конгресса. На тот момент он боролся за приз в конкурсе по поиску молодых талантов под эгидой компании Вестингауз – это был настоящий Святой Грааль для всех одаренных старшеклассников. Как и следовало ожидать, он получил эту премию в 1949 году. (Много лет спустя, в 1980 году, получив приглашение в Стокгольм, в Шведскую академию наук, Гилберт лишний раз улучшил статистику, согласно которой премия Вестингауза – одна из наиболее серьезных заявок на получение Нобелевской премии в будущем.)


Уолли Гилберт (вверху) и Фред Сенгер (внизу) – короли секвенирования
В университете и аспирантуре Гилберт занимался физикой, а в 1956 году, через год после моего прибытия в Гарвард, стал работать на физическом факультете. Когда же я увлек его опытами с РНК, которыми занимался у себя в лаборатории, Гилберт забросил свою дисциплину ради моей. Вдумчивый и непреклонный Гилберт успел немало поработать на переднем крае молекулярной биологии.
Однако из двух методов секвенирования проверку временем выдержал вариант, предложенный Сенгером. Именно этот метод секвенирования был использован в проекте «Геном человека», а затем оказался востребованным и далее, пока не уступил место красивой химической технологии, изобретенной в британском Кембридже (об этом мы поговорим в главе 8). Некоторые химические соединения, расщепляющие ДНК и необходимые при секвенировании по методу Гилберта, сложны в обращении – чего доброго, начнут расщеплять ДНК самого исследователя. В свою очередь, при работе методом Сенгера используется тот же самый фермент, который обеспечивает естественное копирование ДНК в клетках, – ДНК-полимераза. Весь фокус в том, что при копировании пары оснований немного изменяются.
Сенгер использовал не только обычные дезокси-основания (А, Т, Г и Ц), которые встречаются в естественной ДНК, но и так называемые дидезокси-основания. Основания второй категории обладают замечательным свойством: ДНК-полимераза с готовностью внедряет их в цепочку ДНК (то есть копия собирается по образцу матричной цепи). Однако, после того как в цепочку попадет дидезокси-основание, другие основания в нее добавляться больше не могут. Иными словами, скопированная нить не может достраиваться после дидезокси-основания.
Допустим, у нас имеется матричная цепь с последовательностью ГГЦЦТАГТА. В эксперименте используется множество копий такой спирали. Теперь представьте себе, что эта цепь копируется при помощи ДНК-полимеразы, но в растворе, кроме А, Т, Г и Ц, присутствует еще и дидезокси-А. Фермент работает, сначала добавляя к цепи Ц (комплементарный исходному Г), затем еще Ц, затем еще Г и еще Г. Однако, когда фермент добирается до первого Т, открываются два варианта: либо он добавит к растущей цепочке обычный А, либо дидезокси-А. Если фермент подберет дидезокси-А, то цепь далее расти не сможет и получится короткой, с дидезокси A в конце: ЦЦГГддА. Но существует также возможность того, что цепь подхватит обычное A, и в этом случае ДНК-полимераза продолжит добавлять основания: Т, Ц и так далее. Дидезокси-основание в следующий раз сможет «закоротить» цепочку не раньше, чем фермент дойдет до следующего Т. Здесь, опять же, цепочка может подхватить либо нормальное А, либо дидезокси А (ддА). При присоединении ддА цепочка тоже получится обрубленной, но чуть более длинной, чем в первый раз: у этой цепочки будет последовательность ЦЦГГАТЦддА. Подобное происходит всякий раз, когда цепь дорастает до Т и далее к ней может присоединиться А. Если случится так, что цепочка подхватит обычное А, то она продолжит расти, а если подхватит ддА – то на этом завершится.
Что же в итоге? После эксперимента у нас имеется целый набор цепочек разной длины, скопированных с матричной ДНК. Что у них общего? Все они оканчиваются основанием ддА.
Теперь вообразите, что все происходит аналогично и с тремя оставшимися основаниями; в случае Т у нас в растворе будут обычные А, Т, Г, Ц плюс ддТ. В результате будут получаться молекулы ЦЦГГАддТ либо ЦЦГГАТЦАддТ.
Проведя реакцию всеми четырьмя способами – сначала с ддА, затем с ддТ, после этого с ддГ и с ддЦ, – получим четыре набора цепочек ДНК. В первой группе все цепочки заканчиваются на ддА, во второй – на ддТ и так далее. Как можно рассортировать эти слегка различающиеся цепочки в зависимости от слегка различающейся длины так, чтобы можно было логически вывести длину цепочки? Во-первых, можно организовать сортировку, уложив ДНК на пластинку, обработанную специальным гелем, а саму пластинку поместить в электрическое поле. Под действием электрического поля молекулы ДНК рассредоточатся по гелю. Скорость движения каждой цепочки есть функция ее длины – короткие цепочки движутся быстрее длинных. В течение фиксированного промежутка времени самый короткий фрагмент – в нашем случае ддЦ – уйдет дальше всех; чуть более длинный ЦддЦ уйдет не так далеко, а еще чуть более длинный ЦЦддГ пройдет еще меньший отрезок пути. Теперь вы догадываетесь, какой трюк применил Сенгер. Фиксируя относительные позиции всех этих мини-цепочек, движущихся сквозь гель, можно логически вывести, какова последовательность оснований в данном фрагменте ДНК: сначала идет Ц, затем еще Ц, затем Г и так далее.
В 1980 году Фред Сенгер получил Нобелевскую премию по химии совместно с Уолли Гилбертом и Полом Бергом, награжденным за вклад в разработку технологий, связанных с рекомбинантной ДНК (необъяснимо, почему такой чести не были удостоены ни Стэнли Коэн, ни Герб Бойер).
Для Сенгера это была вторая по счету Нобелевская премия. В 1958 году он получил премию по химии за изобретение метода секвенирования белков – он научился определять последовательность аминокислот в белковой молекуле и таким способом выяснил состав человеческого инсулина. Однако сенгеровские методы секвенирования белков и ДНК совершенно не связаны ни в техническом, ни в идейном отношении. Каждый из методов он разработал с нуля, и, пожалуй, Сенгер заслуживает звания величайшего технического гения в ранней истории молекулярной биологии.
Фред Сенгер, умерший в 2013 году, не походил на «типичного» дважды нобелевского лауреата. Он родился в квакерской семье, стал социалистом, а в годы Второй мировой войны отказался от военной службы по религиозным убеждениям. Еще невероятнее, что он нигде не распространялся о своих достижениях, а нобелевские регалии также не хранил на виду. «Получаете красивую золотую медаль и относите ее на хранение в банк. Есть еще сертификат, я храню его на чердаке». Он даже отказался от рыцарского титула: «Рыцарство выделяет вас среди окружающих. А я не хочу выделяться». После ухода на покой Сенгер с удовольствием садовничал у себя дома близ Кембриджа, хотя иногда и посещал Сенгеровский центр (ныне называется «Институт Сенгера») – геномную лабораторию в Кембридже, открытую в 1993 году.
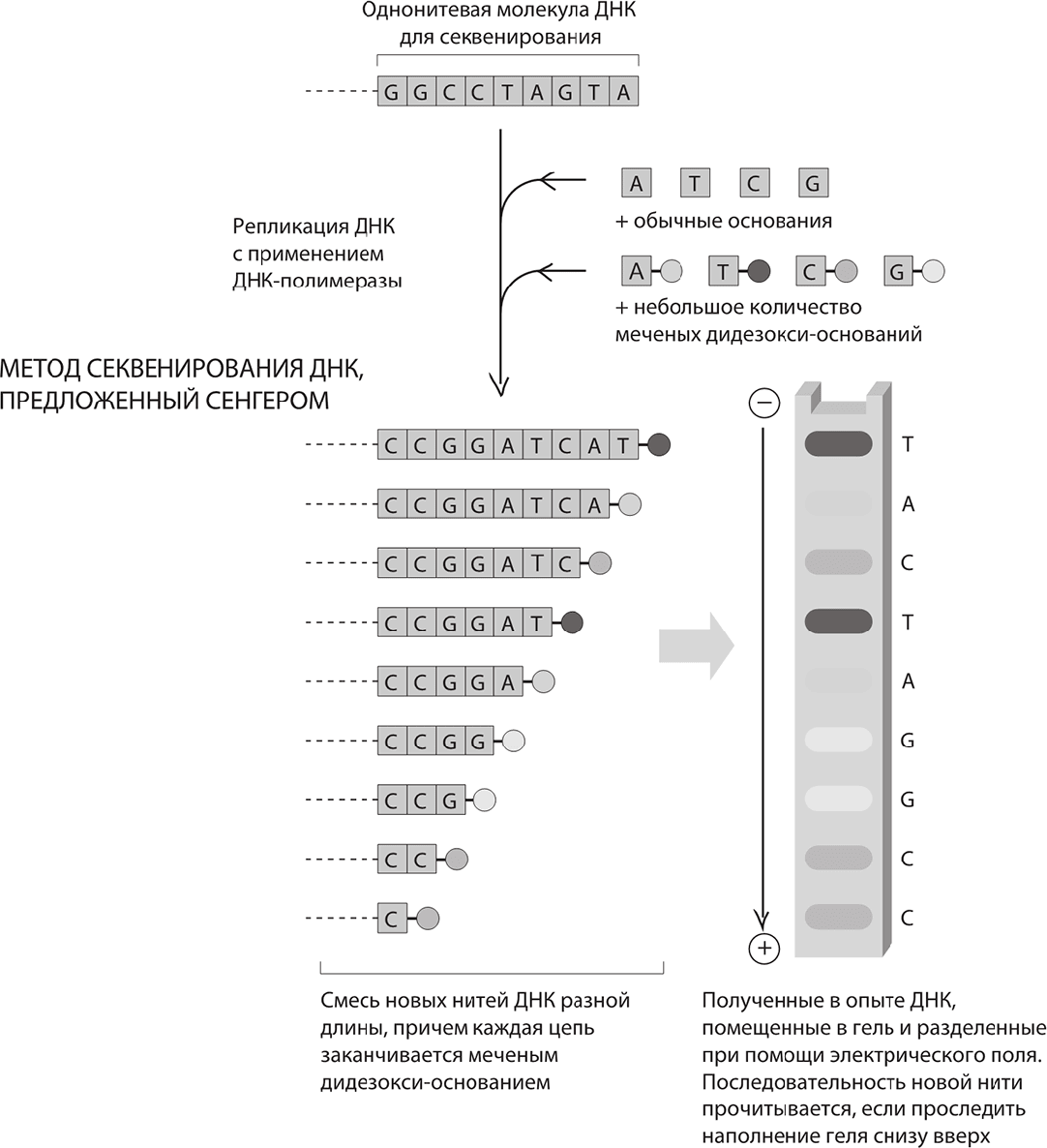
Метод секвенирования ДНК, предложенный Сенгером
Секвенирование подтвердило одно из наиболее замечательных открытий 1970-х годов. Уже было известно, что гены – это линейные цепочки, состоящие из оснований А, Т, Г и Ц, и что эти основания транслируются тройками, в соответствии с генетическим кодом. Из них собираются линейные цепочки аминокислот – такие молекулы называются «белками». Однако замечательные исследования, проведенные Ричардом Робертсом, Филом Шарпом и другими, показали, что у многих организмов гены образуют прерывистые участки и жизненно важные отрезки ДНК перемежаются с нерелевантными. Только после транскрипции матричной РНК эта путаница рассортировывается в процессе «редактирования», при котором ненужные участки удаляются. Это равноценно тому, как если бы в этой книге случайным образом встречались лишние абзацы, с виду перемешанные как попало, и в них рассказывалось бы то о бейсболе, то об истории Римской империи. Уолли Гилберт назвал такие вставные последовательности «интронами», а те участки, которые отвечают, собственно, за кодирование белков (то есть образующие функциональную часть гена), – «экзонами». Оказывается, что интроны встречаются в ДНК сравнительно сложноорганизованных существ; у бактерий их нет.
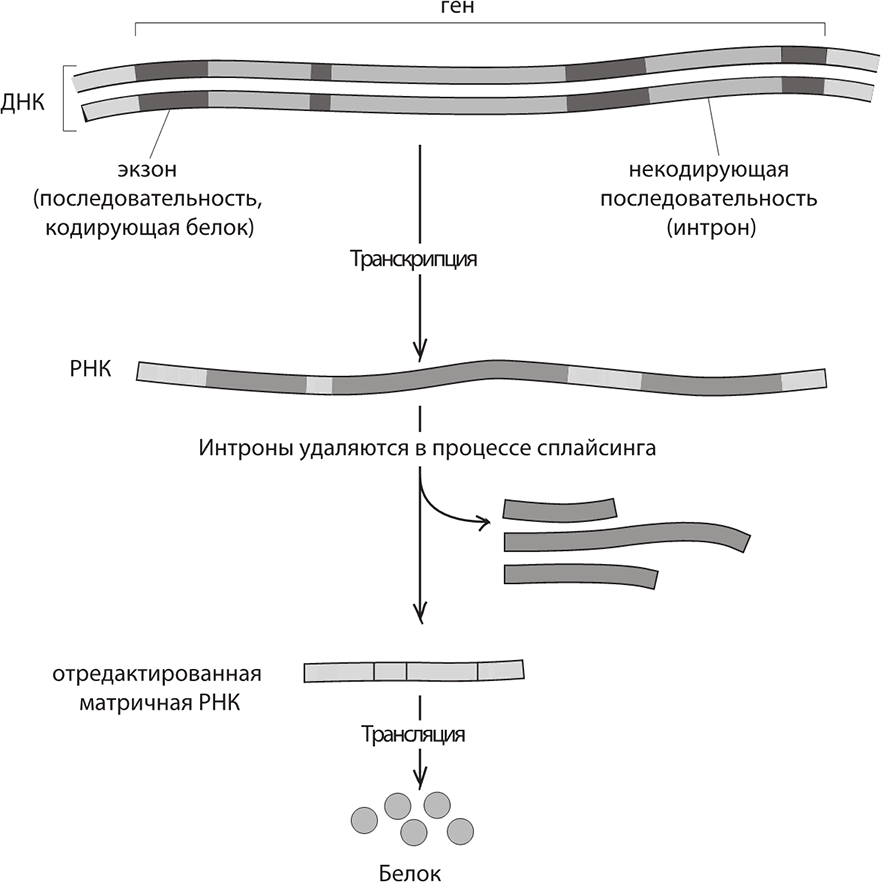
Интроны и экзоны. Некодирующие интроны вырезаются из матричной РНК перед синтезом белков
Некоторые гены особенно богаты интронами. Например, у человека есть ген фактора свертываемости крови VIII (он может мутировать у людей, страдающих гемофилией), который содержит двадцать пять интронов. Фактор VIII – большой белок, его длина составляет около двух тысяч аминокислот, но на кодирующие экзоны в нем приходится всего около 4 % общей длины гена. Оставшиеся 96 % – это интроны.
Каков функционал интронов? Ведь очевидно, что их наличие радикально усложняет все клеточные процессы, поскольку при формировании матричной РНК их всегда требуется вырезать, а это сложное дело, особенно с учетом того, что единственной ошибки при вырезании интрона при подготовке матричной РНК достаточно, чтобы, допустим, фактор свертываемости крови VIII приобрел мутацию сдвига рамки, которая «испортит» весь белок. Существует теория, что такие молекулярные вкрапления – попросту эволюционный рудимент, наследие, сохранившееся со времен зарождения жизни на Земле. Однако до сих пор активно обсуждается, как могли возникнуть интроны и есть ли от них какая-либо польза в великом коде жизни.
Когда в общих чертах стала понятна природа генов у эукариот (организмов, в клетках которых есть специальное хранилище для генетического материала – ядро; у прокариот, например у бактерий, ядра нет), в науке началась настоящая «золотая лихорадка». Группы мотивированных на открытия ученых, вооруженные новейшими технологиями, устроили настоящую гонку: кому первому удастся изолировать (клонировать) и охарактеризовать ключевые гены. Среди первых найденных «сокровищ» были гены, мутации которых вызывают рак у млекопитающих. Как только ученые завершили секвенирование ДНК нескольких хорошо изученных вирусных онкогенов, в частности SV40, удалось выявить конкретные гены, вызывающие рак. Эти гены способны превращать обычные клетки в клетки с онкологическими свойствами – например, в такие, которые бесконтрольно растут и делятся и поэтому образуют опухоли. Уже вскоре молекулярные биологи начали выделять гены из раковых клеток человека, и затем было найдено подтверждение тому, что рак у человека возникает из-за изменений на уровне ДНК, а не из-за обычных негенетических проблем роста, как предполагалось ранее. Были найдены гены, ускоряющие или стимулирующие рост опухолей, а также гены, замедляющие или ингибирующие его. По-видимому, для нормальной работы клетке, как и автомобилю, нужна педаль газа и педаль тормоза.
Генетическое кладоискательство захлестнуло всю молекулярную биологию. В 1981 году в лаборатории Колд-Спринг-Харбор стали читать продвинутый летний курс о приемах генетического клонирования. В ходе этого курса было разработано лабораторное пособие «Молекулярное клонирование», и за следующие три года эта книга разошлась тиражом более 80 тысяч экземпляров. Первый этап революции ДНК (1953–1972) – воодушевление, которое увенчалось открытием двойной спирали и привело нас к чтению генетического кода, – осуществился силами примерно трехсот ученых. На втором этапе, когда была получена рекомбинантная ДНК и разработаны технологии секвенирования ДНК, число революционеров менее чем за десятилетие возросло в сотни раз.
Такой взрывной рост отчасти связан с возникновением совершенно новой индустрии – биотехнологии. Теперь ДНК интересовала не только биологов, стремившихся понять молекулярные основы жизни. Молекула ДНК вышла из академических кулуаров, населенных людьми в белых халатах, в большой мир, где обитают в основном носители шелковых галстуков и строгих костюмов. Френсис Крик назвал свой дом в Кембридже «Золотая спираль», и вскоре это выражение приобрело совершенно новый смысл.
На фото: журнал Time сообщает о рождении биотехнологического бизнеса (а также анонсирует королевскую свадьбу)
Назад: Глава 3 Читаем код: воплощение ДНК
Дальше: Глава 5 Биотехнология: ДНК, доллары и биопрепараты

