Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 36 Зеленая эпитафия динозаврам
Дальше: Глава 38 Прощание с гигантами
Глава 37
Ход c3–c4
Покрытосеменные леса благодаря мощному микоризальному выветриванию усилили сток питательных веществ в океан, что обеспечило фитопланктон удобрением. Последовал буйный рост планктонных водорослей. Так как они являются главными поставщиками затравки для скучивания облаков, в чем водорослям помогал дождевой тропический лес, повысилось альбедо планеты. Одновременно упало содержание в атмосфере парникового углекислого газа, расходуемого на выветривание. (Далее падение только усиливалось: 12 млн лет назад доля CO2 составляла 0,04 %, 3 млн лет назад – 0,03 %.) Так совместными усилиями водорослевый планктон и наземные цветковые, начиная с олигоценового периода (30 млн лет назад), стали понижать температуру на планете. Однако по одним лишь скачкам изотопной подписи углерода рост продуктивности фитопланктона уловить невозможно, требуются дополнительные индикаторы. Одним из них является барит (BaSO4), которого образуется тем больше, чем выше темпы поступления в осадок бария, а попадает этот биогенный металл на дно вместе с остатками фитопланктона, который его накапливает. Начало похолодания как раз совпадает с повышенным содержанием бария в олигоценовых морских отложениях.
Постепенное похолодание выдвинуло на место теплолюбивых лесов совсем другую растительность – травянистые сообщества (степь, саванна, прерия, пампасы, финбош, отчасти тундра и пустыня). Они-то и стали «визитной карточкой» второй половины кайнозойской эры – неогенового и четвертичного периодов (последние 23 млн лет) (рис. 37.1).

Общие изменения во всей биосфере, произошедшие в это время, сопоставимы со среднемеловым ценотическим кризисом, когда цветковые сильно потеснили голосеменные растения. Американский палеонтолог Хелен Таппан образно сравнила наземную и морскую биоту со средневековыми сюзеренами и вассалами, которые трапезничают за одним столом, но при этом последние сидят далеко от солонки и до драгоценной по тем временам соли им не дотянуться. Фитопланктон и другие морские организмы точно так же ограничены в доступе к главным биогенным элементам – азоту, фосфору, кремнию, круговоротом которых полмиллиарда лет управляют растения на суше. Поэтому любые сильные изменения в составе наземной растительности не могут не отразиться на состоянии морской биоты.
В отличие от деревьев, травянистые цветковые растения почти избавились в листовой пластинке от клеток, не осуществляющих фотосинтез. Лист стал тоньше, но как орган, накапливающий энергию и экономящий воду, гораздо эффективнее. При этом травы поразительно устойчивы к выеданию: у потравленных злаков корневище быстро производит новые листья и стебли. При умеренном «выпасе» зеленая масса не уменьшается, а прибывает. Отказались травы и от мощного одеревеневшего ствола, а чтобы выдерживать выедание и вытаптывание травоядными животными, стали вырабатывать в клетках и межклеточном пространстве многочисленные кремневые тельца – фитолиты (от греч. φυτόν – растение и λίθος – камень) размером от тысячных долей до 0,15 мм. По сути, это мелкие острые осколки стекла, оставляющие царапины на зубах животных после пастьбы. По этим царапинам, а также по изотопной подписи азота и углерода в зубах и палеопочвах, собственно, узнали и о «выборе блюд» у древних травоядных, и о времени появления и распространения степных сообществ на разных континентах, а также об условиях, в которых они сложились (рис. 37.2б). (Травы, кстати, освоили малопродуктивные почвы и существенно расширили пригодные для жизни земли.)
У млекопитающих, существенную часть рациона которых составляют травы, на зубах образуется особенно много царапин, как, например, у шерстистого мамонта; у тех, кто больше налегает на веточный корм и плоды с косточками (подобно североамериканскому современнику мамонта – мастодонту), царапин меньше, но появляются ямки. Кроме того, у выедателей – тех, кто пасется, не поднимая головы (бизоны, лошади, носороги), в зубах заметнее присутствие тяжелых изотопов углерода и азота (15N). Зато в зубах скусывателей (таких, как олени) – млекопитающих, предпочитающих ощипывать листочки деревьев и кустарников, этих изотопов относительно мало. Поступают изотопы, конечно, вместе с кормом. Особенности изотопной подписи азота (δ15N) определяются, прежде всего, микоризальным симбиозом растений и влажностью климата: чем он суше, тем беднее почвы азотом и тем круче задирается изотопная подпись, а в ряду травы – кустарники – деревья содержание тяжелого изотопа ниже у последних, поскольку на их обмен веществ существенно влияет микориза, накапливающая легкий изотоп. Самые низкие показатели δ15N, конечно, у полугрибов-полуводорослей – лишайников и, как следствие, у предпочитающих этот корм северных оленей. Есть свой изотопный сигнал у бобовых (акации, дрок, клевер, люпин, люцерна, астрагал). В их тканях соотношение 15N/14N такое же, как в атмосфере, поскольку симбионты бобовых – азотфиксирующие бактерии (ризобии) – улавливают азот прямо из воздуха. Благодаря симбиозу бобовые не только себя обеспечили этим дефицитным в почве и минералах элементом, но и делятся им с другими членами травянистого сообщества, повышая продуктивность. Судя по ископаемым остаткам и данным молекулярного анализа, древовидные бобовые (акации и мимозы) были достаточно разнообразны уже в эоценовую эпоху, а травянистые, как и другая степная растительность, развернулись в миоценовую. Разница изотопных подписей «наследуется» и в тканях хищников, поэтому можно определить, кто на кого предпочитал охотиться. Причем, поскольку легкий 14N выводится с мочой, ткани животных с каждой последующей ступенью трофической пирамиды все больше «тяжелеют», примерно на 3,0–4,1‰.
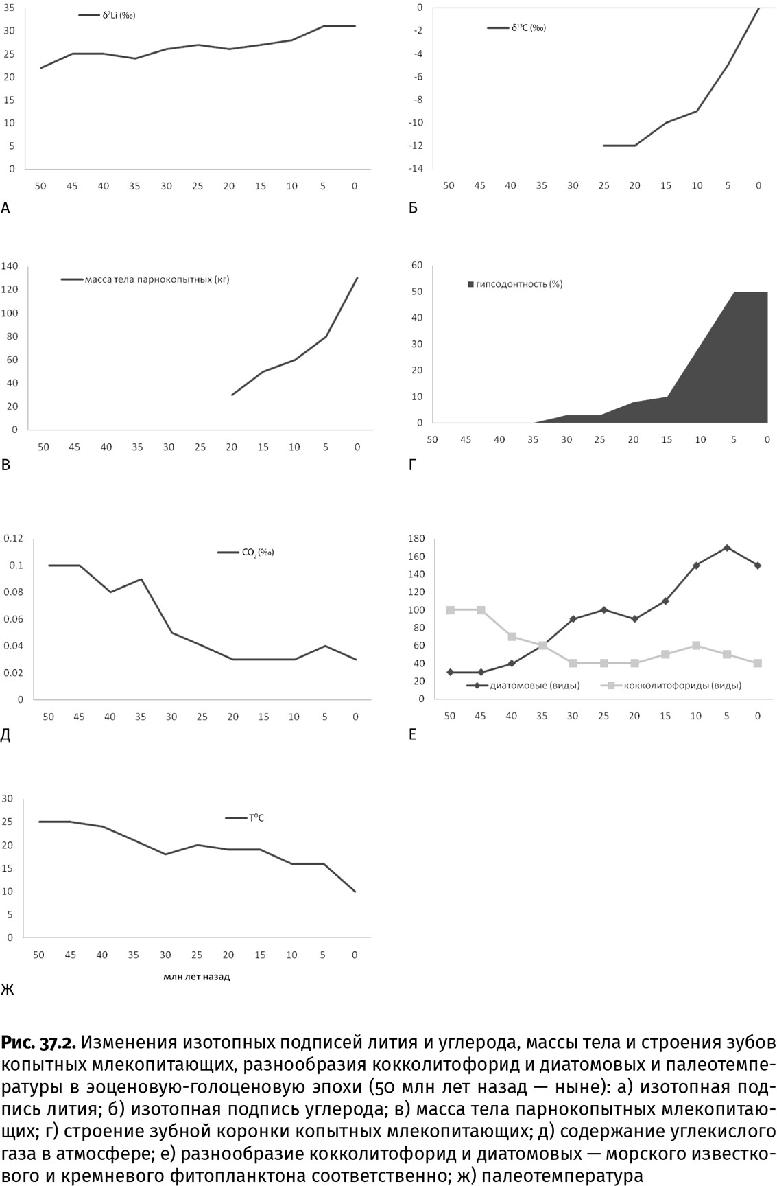
Изотопная подпись углерода в тканях цветковых позволяет узнать, как именно у них происходил круговорот этого элемента. Дело в том, что деревья, большинство кустарников, а также травы прохладного (18–24 °C) и влажного климата (большинство бамбуковых, мятликовых, рис и его родственники) нуждаются в легком изотопе углерода, поэтому их ткани обеднены тяжелым изотопом. Травы жаркого и сухого климата (32–35 °C) с резкими сезонными перепадами, высоким уровнем освещенности, а также растущие на засоленных почвах, довольствуются любыми углеродными изотопами. По заметной разнице в изотопной подписи углерода (около –26,5±3‰ – у первых и всего –12,5±3‰ – у вторых) можно определить, каким именно травам принадлежали растительные остатки. Учитывая поступательное снижение уровня углекислого газа в атмосфере, они приняли своевременные меры и, обходясь малым числом устьиц, меньше теряют влаги. А благодаря новой цепочке фотосинтетических реакций, идущих без фракционирования углеродных изотопов, такие травы не только получили прибавку органического вещества, но и выживают в засуху. К этим растениям, приспособленным к жизни при пониженном содержании двуокиси углерода в атмосфере, относятся многие злаковые, включая важные для человечества культуры – просо, сорго и кукурузу, а также амарант и сахарный тростник.
У большинства наземных растений в фотосинтезе ведущую роль играет цикл Кальвина, в котором углекислый газ включается в органическое вещество. В начале цикла молекула СО2 объединяется с рибулозо-1,5-дифосфатом и молекулой воды, и образуется две молекулы 3-фосфоглицерата с тремя атомами углерода каждая. Поэтому такие растения относят к группе С3. В клетках листовой мякоти реакция катализируется ферментом рибулозодифосфат-карбоксилазой/оксигеназой (рубиско), который при низком уровне двуокиси углерода или в жарком климате перестает распознавать, что за молекула перед ним – углекислый газ или кислород. Растения начинают дышать в дневное время – поглощать кислород вместо двуокиси углерода, при этом тратится энергия, теряются молекулы СО2, захваченные ранее, и в итоге продуктивность может упасть на 40 %.
Травы саванн и прерий пошли другим – С4-путем: у них СО2 сначала присоединяется к фосфоенолпирувату с образованием 4-углеродного оксалоацетата. При этом цикл Кальвина осуществляется в клетках обкладки водопроводящих пучков (плотность которых у этих трав выше), а С4-цикл (Хэтча – Стэка) – в клетках листовой мякоти. Иначе говоря, они отказались от специализированных покровных клеток, превратив их в фотосинтезирующие, и одновременно изолировали центр, где осуществляется цикл Кальвина, от устьиц. Такая анатомия, когда разные углеродные циклы протекают в разных клетках, препятствует потерям углекислого газа и замедляет прохождение (диффузию) его молекул (0,3–0,8 см/с против 28 см/с): больше молекул усваивается. В результате С4-растения запасают значительно больше углекислого газа (40–85 мг/дм3 в час против 1–50 мг/дм3 за то же время у С3-растений). Кроме того, их фермент (фосфоенолпируват-карбоксилаза) захватывает исключительно молекулы СО2, благодаря чему теряется вдвое меньше воды (300–400 г на 1 г сухого вещества против 700–1000 г). С этим всем и связаны повышенная урожайность, малая чувствительность к высокой температуре воздуха и засухостойкость этих цветковых. Хотя травы саванн и прерий представляют 3 % общего видового разнообразия наземных растений, они составляют пятую часть растительного покрова и на их долю приходится четверть продуктивности. Таким образом, С4-растения способны прокормить больше животных. (А ныне стали важнейшими культурами для производства пищи и биотоплива.)
Саванные травы – не единственные растения, сумевшие приспособиться к нынешней атмосфере. (И кстати, между собой они не родственные: С4-растения независимо возникли в шести центрах происхождения на всех континентах, кроме Антарктиды, и представляют собой более 60 самостоятельных эволюционных линий.) К другой группе принадлежат кактусы и еще некоторые суккуленты (толстянка, алоэ, агавы, «живые камни» Южной Африки), сумевшие наладить свой тип фотосинтеза – САМ (аббревиатура от англ. Crassulacea acid metabolism – кислотный обмен веществ по типу толстянковых) в очень засушливых условиях. Цикл Кальвина и С4-цикл у них осуществляются в одних и тех же клетках, но в разное время суток, и газообмен усиливается по ночам, благодаря чему сберегаются и влага, и углекислый газ. Именно потому эти растения и породили огромное разнообразие форм в самых, казалось бы, неподходящих условиях.
Но вернемся к фитолитам: эти кремневые иголки свидетельствуют о том, что основные группы травянистых покрытосеменных – бамбуковые, мятликовые (злаки), родственники риса – зародились в олигоценовую эпоху, но стали фоновым ископаемым материалом с распространением степных сообществ во второй половине миоценовой. Тогда же резко (до 50 %) возросла доля травоядных млекопитающих с гипсодонтными зубами (с высокой коронкой) по сравнению с обладателями мезо- и брахидонтных зубов (с низкой коронкой). («Лошадиные» зубы – это и есть гипсодонтные.) В гипсодонтных зубах и «застряли» изотопы, характерные для С4-трав (рис. 37.2б, г).
Гипсодонтные зубы, в которых, как в зубах жующих динозавров, существенную роль играет цемент, заполняющий впадины между бугорками, выдерживали сильное стачивание при жевании волокнистой пищи вперемешку с фитолитами и песком. Иначе такой наждачный порошок быстро истирал бы эмаль. На срезе подобный зуб напоминает складчатую структуру булата да и по твердости мало чем уступает этой, в несколько слоев прокованной, стали.
Подобно тому как в эоценовую эпоху началась тесная коэволюция костных рыб и шестилучевых кораллов, в миоценовую последовала коэволюция копытных, некоторых других групп млекопитающих, способных пережевывать и переваривать грубоволокнистую пищу, и трав (сначала С3, а потом и С4). Последние отличаются особенно высоким содержанием фитолитов. С распространением таких трав быстро стало нарастать разнообразие гну, ориксов, газелей, вилорогов, буйволов, быков, бизонов и родственных им полорогих, а также некоторых оленей, гиппопотамов, свинообразных, верблюдов, лошадей и тапиров. (В зубах всех этих млекопитающих больше тяжелого изотопа углерода, а их остатки приурочены к отложениям, сформировавшимся в ландшафтах, где могли преобладать С4-травы и, конечно, накопились их фитолиты.)
Многие из этих травоядных копытных – лошади, носороги, верблюды, полорогие, олени, различные группы антилоп, свиньи, жирафы, неполнозубые (броненосцы и ленивцы), а также полностью вымершие копытные Южной Америки – нотоунгуляты (Notoungulata, от греч. νότος – южный и лат. ungulatus – снабженный копытами) пережили сходные эволюционные перестройки: чтобы дать место длинным корням гипсодонтных щечных зубов и вместить расширенные резцы, челюсти и лицевая часть черепа у этих животных вытянулись, зубной ряд сместился вперед по отношению к глазницам и челюстному суставу, а мозговая коробка сдвинулась назад. Особенно усложнились щечные зубы у поздних хоботных – слонов и мамонтов, превратившись в терки с поперечными гребнями.
Поскольку травы богаты клетчаткой и волокнами, для переваривания потребовался вместительный и сложный кишечный тракт, который, как мы уже выяснили на примере позднепалеозойских рептилий, можно уместить только в большом теле. Крупные животные переваривают дольше и успевают извлечь все ценное даже из самой грубой пищи, которую помогает разлагать и сбраживать кишечная микрофлора. У многих непарнокопытных, хоботных и грызунов брожение происходит в толстой кишке, поэтому их называют заднекишечными. Большинство парнопалых, некоторые сумчатые, приматы и древесные ленивцы – переднекишечные, они переваривают пищу в огромных складчатых отделах желудка. У взрослой коровы емкость только одного из этих отделов – рубца – достигает 100–250 л, и в каждой тысячной доле литра рубцовой жидкости проживает 15 млрд и более бактерий. (По сути, корова, как и другие травоядные, потребляет не столько растительную пищу, сколько бактериальные белки.) Заднекишечные непарнокопытные переваривают обильный грубый корм быстро. Жвачные усваивают пищу медленнее, но, поскольку их «передние» ферменты разрушают клетчатку, извлекают больше энергии из того же объема пищи. При этом не обязательно отращивать тушу свыше тонны весом. (Сухопутные гиганты – носороги и хоботные – заднекишечные.) Кроме того, способность питаться худшими кормами позволяет парнопалым не слишком зависеть от сезонной продуктивности трав. Их избирательность в еде тоже имеет преимущество над неразборчивостью лошадей. Итог – больше видов на единицу площади, что стало особенно заметным в плейстоценовую эпоху с распространением трав С4-типа, когда разнообразие парнокопытных резко возросло, а лошадей – сократилось.
Увеличение размеров тела (рис. 37.2в) помогло копытным решить и другую задачу: травянистые угодья отличаются малой растительной массой на единицу площади, и в таких условиях приходилось полагаться не только на зубы, но и на ноги. При значительных увеличениях размеров миоценово-плейстоценовых травоядных по сравнению с лесными предками закономерно удлинились и их конечности, благодаря чему возросла ширина шага. А переход на более твердый грунт привел к сокращению пальцев в стопе и кисти до двух (у многих парнопалых и верблюдов) и даже до одного (у поздних лошадей, литоптернов среди нотоунгулят и некоторых кенгуру). В свою очередь, перераспределение нагрузки, обусловленное сокращением и слиянием скелетных элементов конечности (не только элементов кисти и стопы, но и предплечья и голени), позволило поднять темп ходьбы и бега.
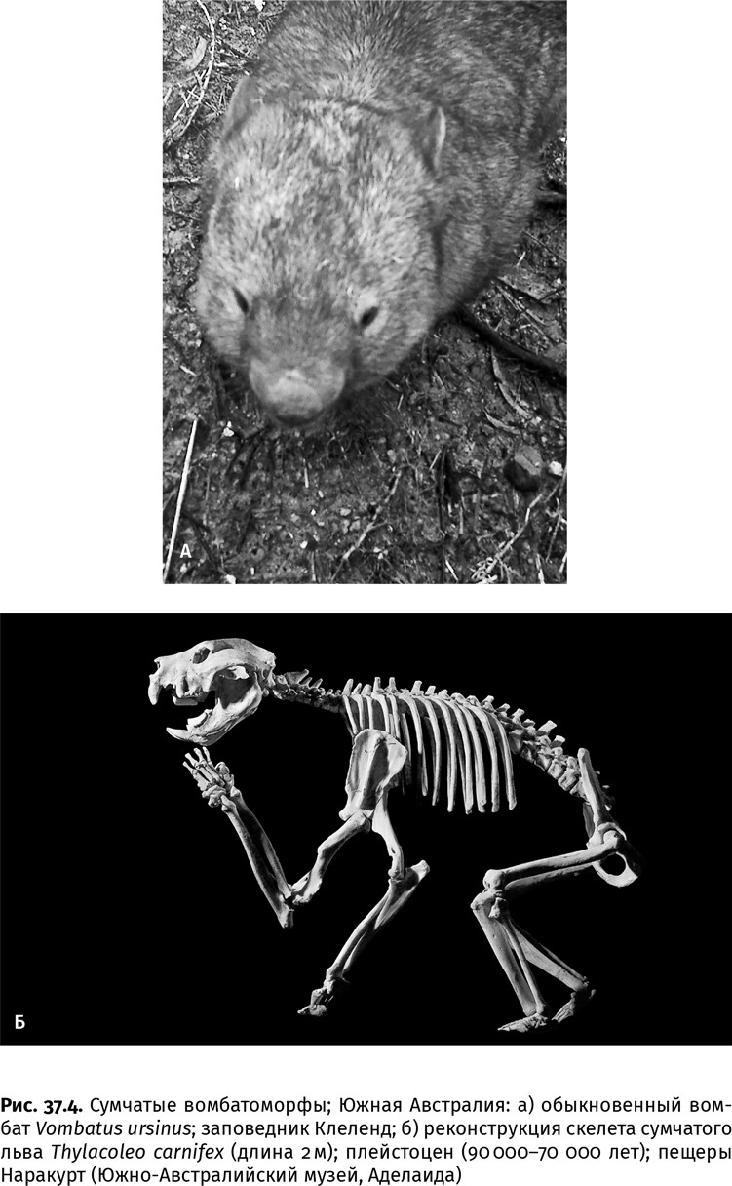
В травянистых сообществах большие стада копытных не выедают растительность подчистую. Этому препятствует разнообразие видов. Каждый из них выбирает только свои травы и «подстригает» их на определенной высоте, а кроме того, в кишечнике разносит семена на десятки километров. Всхожесть многих семян после такой ферментации только увеличивается. Крупные травоядные млекопитающие, такие как хоботные и носороги, повреждают сомкнутый древостой и густые заросли кустарников, не позволяя им восстанавливаться во всей полноте. Так они создают условия для злаков и разнотравья на местах кормления. Кроме того, копытные с большой массой спасают степи от пожаров: в забавном ботсвано-южноафриканском фильме «Боги, наверное, сошли с ума» (1980 г.) бушмен не советует белым африканцам разводить костер, поскольку носорог обязательно прибежит, чтобы его затоптать. И действительно, между пожарами в саванне и белым носорогом, как показали наблюдения, есть определенная связь: этот вид, специализирующийся на травянистой растительности, выедает ее таким образом, что для распространения пожара не остается достаточно горючего материала. В разносе семян помогают грызуны – хомяки, мыши, суслики и др. (рис. 37.3). Запасающие и норные виды особенно важны на открытых пространствах. Они создают в почве хорошо проветриваемые помещения, и по весне в их ходах прорастают надежно укрытые от холодов остатки семян. Причем даже 30 000 лет спустя, как это случилось со смолевкой узколистной, семена которой запасли в норках берингийские суслики. (Вырастить смолевку – современницу мамонта – смогли российские криобиологи под руководством Давида Абрамовича Гиличинского и Светланы Георгиевны Яшиной.) Однако норы важны не только для сохранения семян, но и для аэрации всего почвенного слоя, и норные грызуны сделались самыми мощными (по объемам вскапываемого субстрата) биотурбаторами на суше. Кроме них в этом деле участвуют некоторые насекомоядные (кроты – в Северной Евразии и Америке, златокроты – в Африке и сумчатые кроты – в Австралии), а также броненосцы в Новом Свете и вомбаты на Южном континенте. Современные вомбаты пробивают норы даже в твердой, пропитанной известковыми солями почве – калькрете; колония таких зверей занимает площадь до 1 км2, где отдельные особи роют со скоростью 4 м/ч на глубине до 3 м и оставляют туннели до 60 м длиной (рис. 37.4а). Прежний гигантский вомбат (Phascolonus), величиной и обликом похожий на небольшого мишку с длинными когтями на сильных лапах, при массе до 250 кг и длине до 2 м, вероятно, мог выполнять еще больший объем подземных работ и являлся самым крупным наземным биотурбатором. Лишь некоторые, тоже вымершие, броненосцы приближались к нему по размерам.

Наряду с копытными грызуны стали движущей силой степных сообществ и эволюционировали вместе с травами: в Северной Америке только род «серая полевка» образовал 217 видов и подвидов и всего за 1,5 млн лет, а в Южной за 3,5 млн лет возникло 180 видов хомяковых. Кроме них на Южно-Американском континенте с конца миоценовой эпохи развивалась своя группа кавиоморфных грызунов: американские дикобразы, морские свинки (от их местного имени – Cavia – и происходит название всей группы), агути, капибары, шиншиллы, хутии, нутрии и некоторые другие. (Одна из отличительных особенностей этих грызунов заключается в том, что часть их жевательной мышцы, крепящаяся к лицевому отделу черепа, проходит через большое отверстие в передней стенке глазницы.) У некоторых крупных кавиоморфных видов тоже сформировались гипсодонтные зубы, но у большинства зубы, кроме резцов, мелкие, что компенсируется двойным слоем эмали, а важные для жизни этих существ резцы растут постоянно, впрочем как и у других представителей этого отряда. Ныне 40 % видового разнообразия млекопитающих приходится на грызунов.
В распространении семян соперничать с грызунами могут лишь ткачиковые птицы и некоторые другие воробьиные, снимающие до половины урожая. За последние 30 млн лет воробьиные составили две трети от общего числа современных видов. Зерноядных птиц сдерживают зоркие крылатые хищники и стремительные змеи – ужеобразные (60 % видового разнообразия этих пресмыкающихся), а в Северной Америке – ямкоголовые гадюки (включая гремучих змей). Число и многообразие и тех и других возросло в миоценовую эпоху.
И конечно, в становлении травянистых сообществ большую роль сыграли насекомые. Пришло время пожирающих траву злаковых мушек, мелких цикадок и клопов, а также саранчовых. Саранча предварительно дробит стебли и листья челюстями, а далее в ее кишечнике бактерии высвобождают из растительной массы азотистые вещества и вносят витамин В: в почву опять же попадают готовые удобрения. («Грозой полей» саранча становится тогда, когда степное разнотравье превращается в монокультурные посадки.) Копрофаги – жуки-навозники и навозные мухи (точнее, их червеобразные личинки – опарыши) научились успешно перерабатывать груды навоза, также превращая этот неизбежный продукт в удобрения. Благодаря их деятельности до 80–90 % массы навоза изымается в первые недели, а потери азота снижаются с 80 до менее 15 %. Вместе с личинками мух и крылатыми падальщиками – стервятниками жуки-могильщики и трупоеды споро убирают падаль. Отмершую растительность перерабатывают термиты (в саваннах) и жуки-чернотелки (в степях), но появились обе эти группы в меловых лесах (рис. 37.5). Следствием работы «тружеников невидимого фронта» становятся высокоплодородные почвы, такие как чернозем. А одиночные пчелы-опылители способствовали дальнейшему преображению разнотравья.

На всех континентах, кроме рано (более 30 млн лет назад) обледеневшей Антарктиды, травоядные отращивали зубы, учились быстрее бегать, прибавляли в весе. Хотя сходные ниши заняли достаточно разные животные, они в чем-то уподобились друг другу, поскольку кто-то должен был выполнять работу выедателей (собственно, тех, кто пасется, не поднимая головы) и скусывателей, питающихся листьями, ветками и древесным подростом (рис. 37.6). Даже в плейстоценовую эпоху, несмотря на суровый холодный климат и ледяной щит, покрывший треть суши в Северном полушарии, степная мегафауна процветала. В то время (2,58 млн лет назад) травянистые сообщества покрывали значительную часть суши там, где сейчас широколиственные и тропические леса, тайга. По бескрайним степям бродили несметные стада копытных и хоботных, на которых охотились огромные хищники, включая саблезубых кошек, расплодились грызуны, змеи и воробьиные птицы.
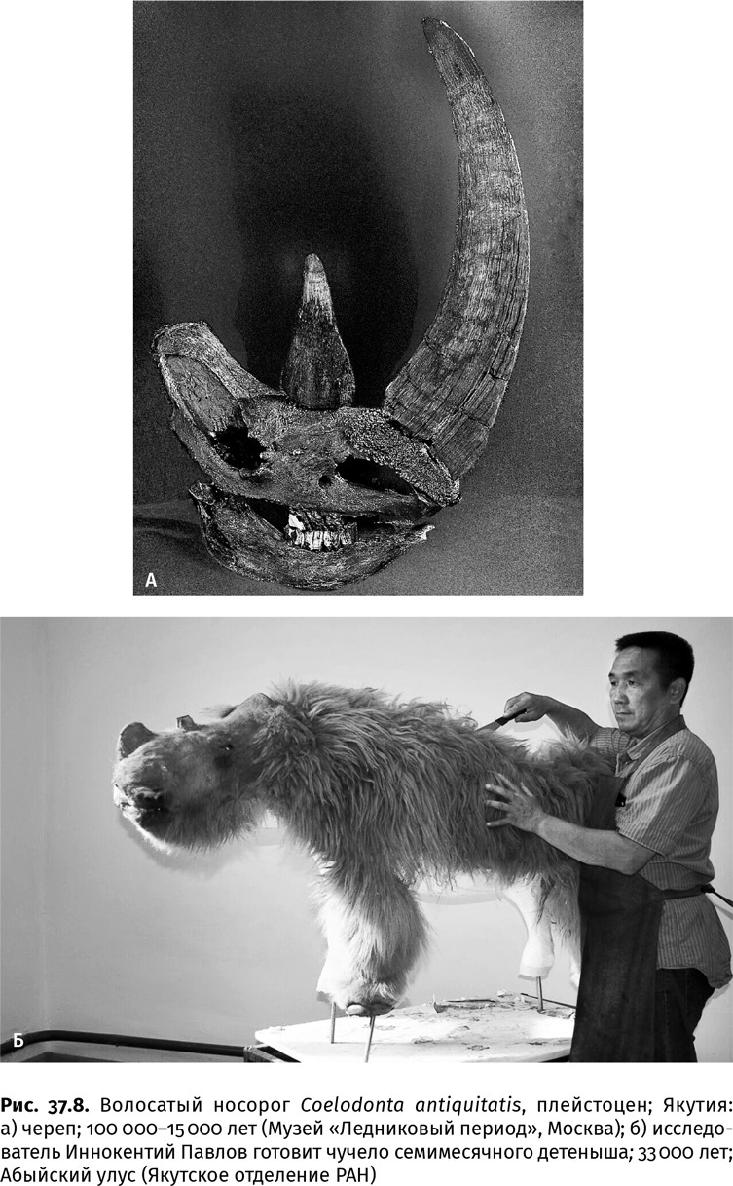
В Приполярье и Заполярье Евразии обитали шерстистые мамонты, волосатые носороги, бизоны, овцебыки, сайгаки, северные олени и лошади, а в Европе, кроме них, эласмотерии (огромные однорогие носороги), туры, большерогие олени и пещерные медведи (последние были скорее растительноядными, чем даже всеядными); охотились на них волки, пещерные гиены и пещерные львы (рис. 37.6а, 37.7–37.16). (Львы и гиены в пещерах, в отличие от впадавших в спячку медведей, не жили, просто в таких условиях лучше сохраняются их остатки.) Жизнь в суровом климате привела к тому, что у этих животных появились сходные приспособительные признаки, например сложный волосяной покров; бурый жир, служивший энергетической подпиткой в случае бескормицы; возможно, молекулярные антифризы, как у мамонтов, у которых гемоглобин имел большее сродство к кислороду, что помогало дышать на морозе. Северные млекопитающие активнее использовали для добычи пищи зимой бивни, рога и копыта (все эти органы носят следы серьезного износа), чем их собратья, обитавшие в благоприятных условиях.
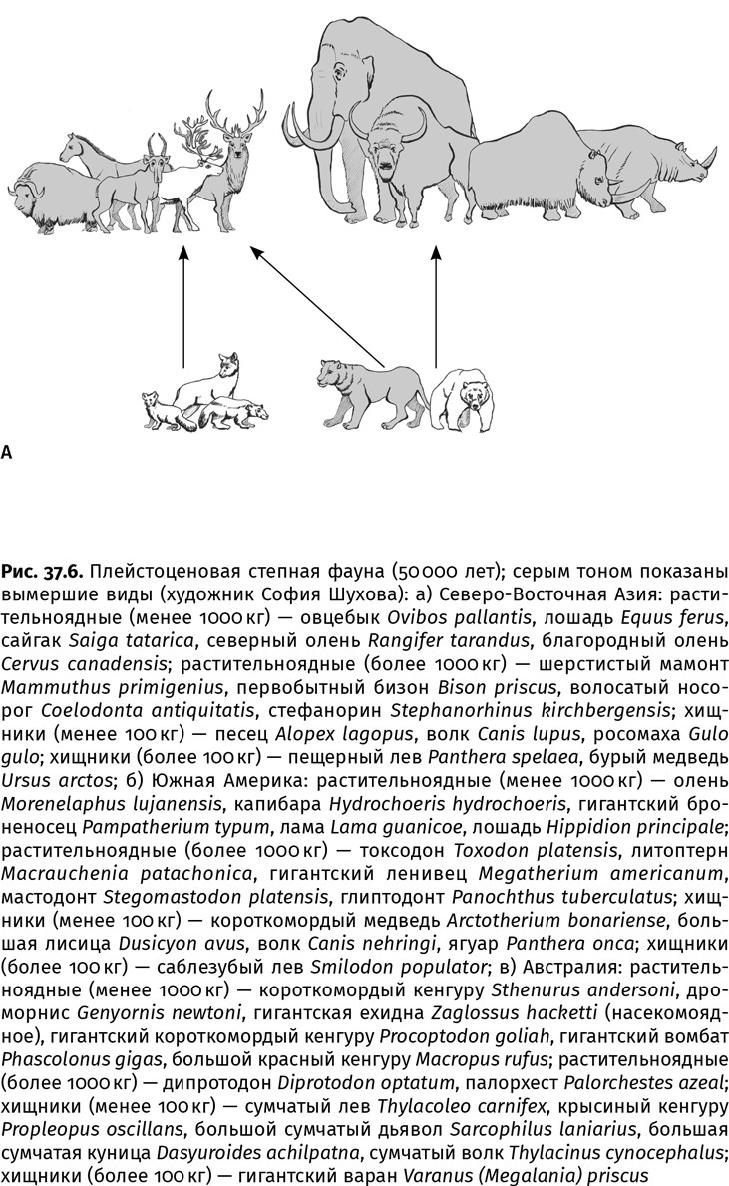
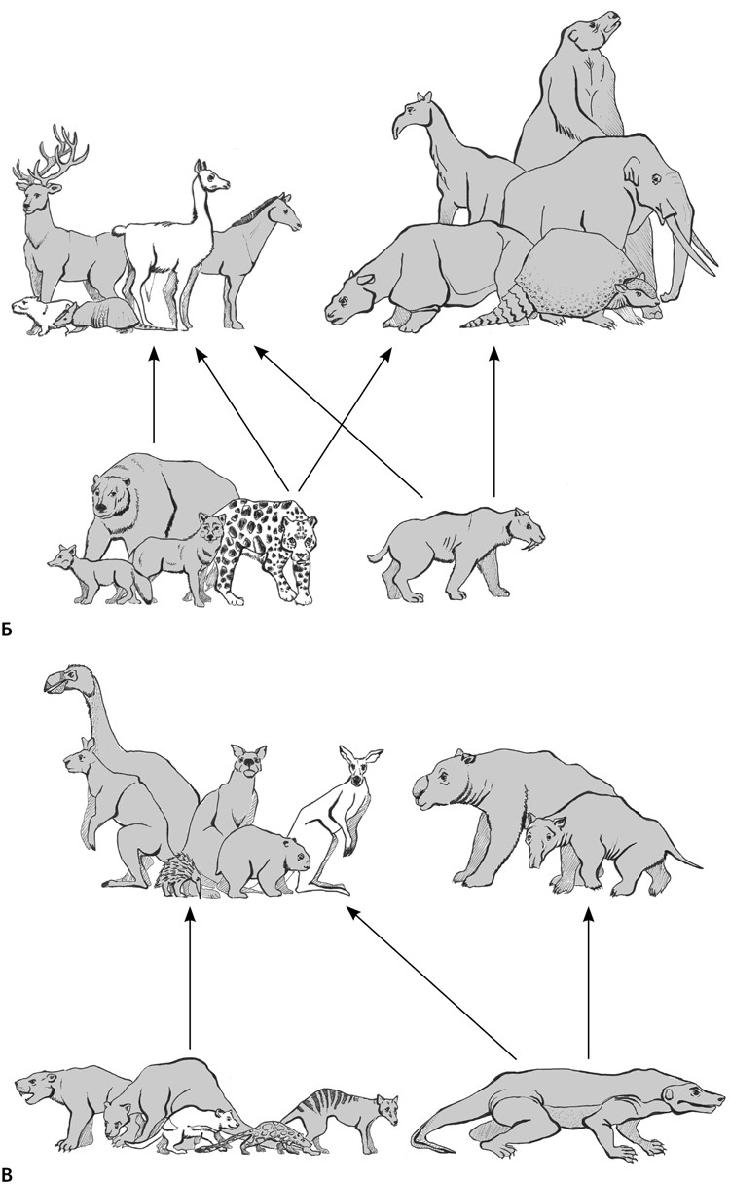
Северная Америка, хотя и сообщалась с Евразией, поскольку уровень моря упал из-за образования снежных шапок и возник широкий Берингийский мост, тоже отличалась своеобразием. Кроме мамонтов, бизонов, овцебыков, сайгаков и пещерных львов, пришедших в плейстоценовую эпоху из Азии, здесь сохранились реликтовые хоботные – мастодонты (Mastodontidae, от греч. μαστός – сосок и οδούς – зуб) и гомфотерии (Gomphotheridae, от греч. γόμφος – шкворень и θηρίον – зверь), больше питавшиеся веточным кормом. У древних мастодонтов щечные зубы несли не гребни, а отдельные похожие на сосочки бугорки; такие архаичные зубы сохранились у гомфотериев. Кроме того, у мастодонтов было две пары бивней: длинные – верхние и короткие – нижние. С юга по Панамскому перешейку добрались сюда южноамериканские мегазвери – гигантские ленивцы и глиптодонты (Glyptodontidae, греч. γλϋφή – резьба и οδούς – зуб), а также дикобразы и последние американские сумчатые – опоссумы. Глиптодонты при длине до 4 м и массе до 2000 кг оделись в тяжелый наружный панцирь, их голову прикрывал костный шлем, а хвост был задраен в костные кольца и нес на конце шипастую булаву. Гигантские ленивцы, или мегатерии (Megatheriidae, греч. μέγας – большой и θηρίον – зверь), массой до 4000 кг, могли вставать на задние конечности, чтобы объедать высокие деревья, пригибая ветки длинными передними лапами с кривыми когтями и обрывая листья длинным языком (рис. 37.17). У тех и других были огромные носовые полости с окостеневшей носовой перегородкой для крепления усиленной мускулатуры губ, которыми гиганты собирали траву. Еще здесь водились тапиры, верблюды и ламы, вилороги, пекари (свинообразные), а охотились на копытных и хоботных ягуар, пума, американский гепард (Miracinonyx), американский лев и саблезубая кошка – смилодон (Smilodon). Смилодоны вели очень нетипичный для кошачьих стайный образ жизни. Еще один местный вид – короткомордый медведь (Arctodus simus) – при массе около 1000 кг мог бы являться крупнейшим хищником среди когда-либо существовавших млекопитающих, и его размер был бы предельным для плотоядных животных с высокими темпами обмена веществ, но, судя по характеру стачивания зубов, это был всеядный зверь, активно потреблявший грубоволокнистую растительность.

Многие из этих млекопитающих проследовали навстречу южным выходцам и практически уничтожили самобытную фауну Южной Америки, 50 млн лет надежно отделенную океанами от прочих континентов. Более активный Север одержал полную победу над медленным Югом. Из былого богатства уцелели лишь неполнозубые (ленивцы, глиптодонты, броненосцы и родственные им пампатерии, муравьеды), кавиоморфные грызуны да несколько нотоунгулят – литоптерн макраухения (Macrauchenia), похожий на верблюда с хоботком, и немного напоминавший носорога токсодон (Toxodon). Несмотря на значительные утраты, в Южной Америке сохранилось высокое разнообразие именно мегафауны: более 80 видов массой свыше 100 кг, в том числе 20 «мегазверей» – более 1000 кг (рис. 37.6б). (Капибара Neohoerus достигала рекордного для грызунов веса – 110 кг.) Возможно, столь огромная биомасса здешних мегатравоядных была обусловлена относительно невысокими для млекопитающих темпами обмена веществ; жевали они тоже не лучшим образом: зубы имели, хотя и гипсодонтные, но с небольшой жевательной поверхностью.
Самое необычное сообщество травоядных сложилось в Австралии, Новой Гвинее и Тасмании, представлявших тогда единый материк – Сахул (рис. 37.6в). На этом естественном «Ноевом ковчеге», пребывающем 45 млн лет в одиночном дрейфе, даже среди растений местные виды составляют более 80 %. В сухих и иссушающих австралийских условиях в миоценовую эпоху сложились свои саванны. Частые пожары, периодически уничтожавшие всю растительность, вызвали к жизни весьма мудреные приспособления. Многие растения, например банксии, обзавелись плодами в грубой кожуре, которая лопается только в огне. Без нагрева они просто не размножаются и после пожара выглядят как обугленные головешки, разевающие многочисленные ярко-желтые или оранжевые рты, – это раскрылись плоды. Толстая кора надежно сохраняет сердцевину объятого пламенем ствола. А эвкалипты будто стремятся воспылать сами, накапливая в листьях горючее миртовое масло. Зола же, как известно, служит прекрасным удобрением. Австралийские сумчатые (вомбаты, коалы, кенгуру и валлаби) ведут свое начало от мелких всеядных древолазов вроде современных, похожих на крыс, соневидных опоссумов, вместе с предками утконоса и ехидны перебравшихся сюда в палеоценовую эпоху из Южной Америки через Антарктиду. В миоценовую эпоху, как только травянистые сообщества начали наступать на дождевой лес, с сумчатыми произошло то же самое, что и с млекопитающими других континентов. Зубы коал и поссумов приспособились к шершавым грубым листьям эвкалиптов, а кишечная «микрофлора» – к ослаблению воздействия ядовитых миртовых масел. (Интересно, что древнейшие, среднеэоценовые, остатки листьев и семенных коробочек эвкалиптов найдены там, откуда перебрались в Австралию сумчатые, – в Южной Америке. Может быть, и семена они распространили.) При переходе из лесов на пастбища возникли десятки видов кенгуру. Их кишечник содержит бактерии, переваривающие клетчатку. Кенгуру сбились в стада и подросли: плейстоценовые виды (Protemnodon, Sthenurus) – до 2,6 м и 230 кг. С удлинением и усилением задних конечностей и хвоста они перешли на быстрый рикошетирующий прыжок, а у некоторых вымерших форм на задних ногах осталось по одному пальцу, как у лошадей или литоптернов. Вместе с кенгуру паслись дроморнисы (Dromornithidae), или, на языке аборигенов, михиранги – огромные (до 3,6 м высотой) бегающие гусеобразные птицы с длинными ногами и копытоподобными когтевыми фалангами. (В данном случае термин «гусеобразные» означает систематическую принадлежность, клюв у них мало напоминал гусиный.) Гигантский вомбат по строению зубной системы (бескорневые, постоянно растущие, двулопастные щечные зубы и пара верхних и нижних резцов) уподобился грызунам. Древостой нарушали сумчатые размером с носорога (до 2800 кг), но с хоботком вместо рога на морде – сумчатые тапиры (Palorchestes) и дипротодоны (Diprotodon). Гипсипримнодонтиды (Hypsiprymnodontidae, от греч. ϋψος – верх, πρυμνός – крайний и οδούς – зуб), или крысиные кенгуру, а также сумчатые волки (Thylacinidae) и дьяволы (Dasyuridae) исполняли роль среднеразмерных насекомоядных, плотоядных и падальщиков. А вершину пищевой пирамиды венчал лев, тоже, конечно, сумчатый (Thylacoleo, от греч. θύλακος – мешок, сумка и λέων – зверь): полосатый, что известно по рисункам коренных австралийцев, и массивный (100–130 кг). В отличие от большинства плацентарных хищников, он разрывал добычу режущими предкоренными зубами, занимавшими четверть челюсти. Следы этих зубов найдены на костях ископаемых сумчатых. Но, притом, что «лев» не уступал в силе укуса смилодону, в строении его скелета наблюдается много странностей: передние конечности заметно длиннее задних, хвост, видимо усиленный, как у кенгуру, и в целом обликом он напоминает древолаза. (Впрочем, крупный хищник – леопард – тоже древолаз, хотя и не отличается столь экзотичной внешностью.) Примечательно, что в отсутствие специализированных плотоядных в Сахуле таковыми становились представители самых разных групп: сумчатый лев был родственником дипротодонтов, коал и вомбатов (рис. 37.4), а гипсипримнодонтиды являлись «озверевшими» кенгуру. Только появившиеся в олигоценовую эпоху тилациниды-дазиуриды были специализированными плотоядными, хотя и не очень большими. Самым же крупным хищником здесь оказался гигантский (до 6 м длиной и до 200 кг массой) родственник комодского варана – мегалания [Varanus (Megalania) priscus], тоже ядовитый.




Поскольку размеры растительноядных млекопитающих определяются площадью пастбища, на островах мамонты, слоны и бегемоты мельчали, и нуждавшиеся в крупной пище плацентарные хищники там выжить не могли. (Сотня зайцев не заменит одного быка: на их поимку нужно потратить в 100 раз больше усилий, на возмещение которых потребуется еще больше пищи.) Поэтому место последних занимали гигантские насекомоядные (ежи, землеройки), хищные птицы и пресмыкающиеся. Покрытая обширными пустынями Австралия в этом отношении скорее большой остров, чем маленький континент.






С повсеместным развитием травянистого покрова, содержащего фитолиты, усилился вынос в океан кремнезема и начался расцвет его потребителей – диатомовых, силикофлагеллят и фораминифер-силиколокулинин. Замерить усиление потока кремнезема напрямую пока сложно, но можно сделать это косвенно – по изотопной подписи лития (7Li/6Li, или δ7Li). Поскольку оба изотопа очень легкие, они сильно различаются по массе, что приводит к заметному фракционированию (до 40‰) при выветривании. При этом 6Li остается в новообразованных минералах, а 7Li выносится в океан. Так как суша – основной источник тяжелого изотопа (в гидротермальных поступлениях его в четыре раза меньше), изотопная подпись этого металла позволяет оценить изменения в темпах выветривания. Оказывается, что ускорение видообразования у диатомовых и время формирования самых мощных диатомитов приходится на резкий рост δ7Li (рис. 37.2а, е). Как говорится, что и требовалось доказать. (Можно использовать для тех же выводов изотопную подпись стронция, но ее разрешающая способность ниже из-за меньшей разницы изотопных масс.)
Так как в степных почвах содержится в 5–10 раз больше гумуса, чем в лесных, возрос и сток в океан биогенных веществ, где появились морские луга, на этот раз без кавычек, поскольку их создали освоившие морскую среду покрытосеменные: на мелководье, даже с карбонатным илом, образовались заросли талассии (Thalassia), или черепаховой травы, сирингодиума (Syringodium), или ламантиновой травы и др., где завелись свои крупные специализированные травоядные – морские черепахи и сирены (Sirenia, родственники хоботных), включая девятиметровую стеллерову корову, истребленную в XVIII в. Обкусывая и перекусывая листья и стебли этих растений, морские травоядные помогают им, перешедшим на бесполое размножение, распространяться. В свою очередь, стабилизируя ил, морские травы понижают количество взвеси в водной толще, и по соседству с лугами строятся коралловые рифы.
С умножением диатомовых и силикофлагеллят современный океан обеднел растворенным кремнеземом. Нуждавшимся в этом минерале радиоляриям пришлось облегчать собственный скелет, их разнообразие сократилось, а стеклянные губки отступили на глубину. Снизилось обилие и видовое разнообразие (в пять и более раз) кокколитофорид и динофлагеллят, которых диатомовые потеснили (рис. 37.2е). Усиление выедания диатомовых планктонными каляноидными веслоногими раками (Calanoida), которые обзавелись очень крепкими, тоже окремневшими, челюстями-мандибулами, привело к уменьшению размеров водорослей, но толщина раковины возросла, и потребление растворенного кремнезема только повысилось. Поскольку накопление на дне известковых кокколитов происходит с выделением двуокиси углерода, а осаждение отмерших диатомовых, на которых сейчас приходится 60 % потока органического вещества, – с поглощением этого газа, казалось бы, простое замещение одних «красных» водорослей другими привело к огромным последствиям. Уровень углекислого газа в атмосфере начал падать, что закономерно привело к похолоданию, известному нам как современный ледниковый период (рис. 37.18). Круг замкнулся: степные травы концентрировали и поставляли кремнезем в океан, способствуя процветанию диатомовых, а диатомовые понижали уровень углекислого газа, помогая травам расширить степные угодья (рис. 37.2б, д – ж).


Усилению похолодания могло поспособствовать расширение тундры, особенно сфагновых болот: в таких болотах подавляется деятельность бактерий и тем самым – выход СО2 и СН4; они экранируют солнечное излучение, не позволяя многолетнемерзлым грунтам протаивать на большую глубину даже в теплое время, и усиливают эвапотранспирацию, способствуя формированию туманов и облаков. В целом альбедо болотных угодий в два раза выше, чем у тайги, летом и в пять раз выше, чем зимой.
Со степными ландшафтами связано еще одно очень важное для планеты новшество – появление человекообразных приматов. Разделение ветвей шимпанзе и ранних австралопитеков (Australopithecus), произошедшее 6–7 млн лет назад, пришлось на один из максимумов распространения саванн и редколесий в Африке, а взрывное видообразование в роду собственно человека (Homo), а также парантропов (Paranthropus), случившееся 2,7–1,5 млн лет назад, приходится на развитие саванн с травами С4-типа.

