Книга: От атомов к древу. Введение в современную науку о жизни
Назад: 16. криптозой
Дальше: “Философский камень” биологии
17. фанерозой
И солнце парит топь в полдневном жаре,И в зарослях хвощей из затхлой мглыВозносятся гигантских сигиллярийУпругие и рыхлые стволы.
Косматые — с загнутыми клыками —Пасутся мамонты у мощных рек,И в сумраке пещер под ледникамиКремень тяжелый точит человек.Михаил Зенкевич. Темное родство
Конец эдиакарского периода является в то же время рубежом двух эонов — протерозоя и фанерозоя. И вот тут нужно небольшое пояснение.
“Фанерозой” буквально означает “явная жизнь”. Это эпоха, к которой относится подавляющее большинство изучаемых палеонтологами ископаемых остатков. Все предшествующие времена, включая протерозой, архей и катархей, собирательно называют криптозоем — “скрытой жизнью”. Фанерозой же, в свою очередь, делится на три эры, названия которых, скорее всего, знакомы большинству читателей: палеозой, мезозой и кайнозой. “Палеозой” буквально означает “древняя жизнь”, “мезозой” — “средняя жизнь” и “кайнозой” — “новая жизнь”. А каждая из этих эр делится на периоды. Период, с которого начинается палеозой (и тем самым весь фанерозой), называется кембрием. Как и многие другие геологические периоды, кембрий получил свое имя из географии: Кембрия — это римское название Уэльса, кельтской страны на западе Британии. Соответственно, очень распространенный синоним криптозоя — докембрий.
Чтобы правильно видеть перспективу, будем помнить следующее: весь фанерозой составляет (округленно) всего лишь примерно 1/9 как от времени существования Земли, так и от истории жизни на ней. Остальные 8/9 — это криптозой, или докембрий, которому посвящена предыдущая глава. Иное дело, что в фанерозое события очень сгущаются.
В 1845 году шотландский геолог Родерик Мурчисон предложил назвать все времена до начала кембрия азойской эрой, то есть — буквально — безжизненной. Долго это название не продержалось: уже палеонтологи XIX века показали, что в толще докембрийских пород есть следы жизни. А сейчас мы точно знаем, что жизнь существовала на Земле на протяжении большей части докембрия, и можем датировать многие ключевые события докембрийской истории — например, кислородную революцию или появление многоклеточности. И о том и о другом мы вкратце говорили в предыдущей главе.
“Большой взрыв жизни”
Главное отличие фанерозойской жизни от докембрийской — колоссальное обилие многоклеточных животных. Причем подавляющее большинство этих животных с самого начала относится уже к современным типам. В кембрии появляются губки, гребневики, стрекающие, круглые и кольчатые черви, членистоногие, моллюски, брахиоподы, иглокожие, полухордовые и хордовые. Внезапное появление всех этих животных в палеонтологической летописи принято называть кембрийским взрывом. В более древних слоях их остатков нет (по крайней мере, определяемых однозначно и бесспорно). Таким образом, кембрий — это время рождения фауны, близкой к современной. Кембрийский взрыв дал такой эффект и произошел так быстро, что его часто называют “эволюционным Большим взрывом” — по аналогии с тем Большим взрывом, в котором родилась Вселенная.
Еще кембрийский взрыв иногда называют скелетной революцией. Действительно, многие появившиеся в этот момент группы животных приобрели твердые скелеты, причем совершенно разные по структуре и на разной биохимической основе: например, между спикулами губок, раковинами моллюсков и хитиновыми панцирями членистоногих нет буквально ничего общего. Такая одновременность никак не может быть случайной. Тем не менее “кембрийский взрыв” и “скелетная революция” — не синонимы. Во-первых, твердые скелеты были далеко не у всех кембрийских животных (например, у первых хордовых их не было). Во-вторых, и в докембрии иногда попадаются явные скелетные структуры — например, непонятно кому принадлежавшие жилые трубки. В общем, понятие “кембрийский взрыв” куда более определенное, и неудивительно, что современные авторы чаще говорят именно о нем.
А был ли взрыв?
Но вот вопрос: произошел ли кембрийский взрыв на самом деле? Существует мнение, что многие современные группы животных появились еще в глубоком докембрии, но долгое время не оставляли ископаемых остатков, а потому были палеонтологически “невидимы”. Причины этого могли быть разными: мелкий размер животных, отсутствие у них твердых скелетов или просто неподходящие для захоронения физические условия. Гипотеза “долгой скрытой докембрийской эволюции” неплохо поддерживается молекулярной филогенетикой, основанной на сравнении аминокислотных и нуклеотидных последовательностей белков и генов разных животных (разумеется, современных — ведь со времен кембрия ни белков, ни ДНК не осталось). Реконструкции, сделанные исключительно по молекулярным данным, зачастую уводят корни современных типов животных даже не в эдиакарий, а в предыдущий период — криогений. Тогда выходит, что кембрийский взрыв — это не столько реальное эволюционное событие, сколько артефакт сохранности. На рубеже кембрия животные просто “проявились”, обретя твердые скелеты и начав захораниваться в осадочных толщах. А возникли они гораздо раньше.
Однако есть впечатление, что тщательной проверки, при которой молекулярно-биологические данные аккуратно шаг за шагом сопоставляются с палеонтологическими, гипотеза “долгой скрытой докембрийской эволюции” все же не выдерживает. В 2013 году группа исследователей под руководством австралийского палеонтолога Майкла Ли попыталась провести такую проверку, использовав материал по самому многочисленному типу животных — членистоногим. Были рассмотрены и морфологические признаки, изучаемые палеонтологами, и последовательности ядерных генов, изучаемые генетиками. Специальная компьютерная программа построила по всему этому массиву данных единое эволюционное древо (причем молекулярные последовательности она учитывала наравне с морфологией). Назовем его для краткости “древом Ли”. Программа, использованная для построения древа Ли, могла определить скорость эволюции на каждом отрезке этого древа и в результате вычислить наиболее вероятные даты расхождений эволюционных ветвей. Это и было целью исследователей.
Теперь давайте следить за цифрами. Считается, что кембрийский период начался 542 миллиона лет назад. Согласно древу Ли, расхождение двух главных эволюционных ветвей членистоногих — Chelicerata (мечехвосты и паукообразные) и Mandibulata (многоножки, ракообразные и насекомые) — произошло 543 миллиона лет назад. А расхождение двух подгрупп внутри Mandibulata, многоножек и ракообразных, произошло 538 миллионов лет назад. Как видим, граница кембрия попадает точно между этими датами: Chelicerata и Mandibulata разошлись перед самым началом кембрия, а многоножки и ракообразные — сразу после него. Понятно, что точность таких датировок не абсолютна, но уж случайным совпадением они не могут быть никак. Итак, мы видим, что рождение главных эволюционных ветвей членистоногих произошло в ближайшей временной окрестности нижней границы кембрия. На этот момент действительно пришелся пик изменений.
Получается, что кембрийский взрыв — никакой не артефакт. Вероятно, большинство крупнейших эволюционных ветвей животных действительно возникло в ближайшей временной окрестности границы кембрия (плюс-минус несколько миллионов лет). Есть и математические модели, подтверждающие, что погруженные в докембрий “стволы” эволюционных древес современных типов животных должны быть короткими. Время существования этих докембрийских “стволов” — считаные миллионы лет, может быть, первые десятки миллионов, но уж никак не сотни миллионов. В общем, на данный момент у нас достаточно оснований, чтобы считать гипотезу “долгой скрытой докембрийской эволюции” скорее неверной, а кембрийский взрыв — реальностью, как это, собственно говоря, и следует напрямую из палеонтологических данных.
Чтобы ослабить категоричность, добавим: вывод, который мы только что сделали, безусловно обладает свойством фальсифицируемости. Это означает, что можно сформулировать четкие условия, при которых он будет опровергнут. Например, для этого достаточно найти хотя бы одного достоверно определимого скорпиона (или сороконожку, или улитку) криогениевого возраста. Но пока такого не было, и вероятность, что это когда-нибудь случится, с каждым годом уменьшается.
Причины взрыва
Итак, в начале кембрия невероятно быстро возникло множество новых крупных эволюционных ветвей животных. Такого не случалось больше никогда, ни раньше, ни позже. Даже после катастрофических массовых вымираний (о которых речь впереди) животный мир восстанавливался за счет нарастания разнообразия уже существовавших больших групп, а не за счет появления новых. Вот почему кембрийский взрыв обязательно требует объяснения.
Правда, “невероятно быстро” не значит “мгновенно”. Новые группы животных отнюдь не появились в начале кембрия все разом в полном составе, как актеры после подъема занавеса. Кембрийский взрыв был хоть и сильно сжатым во времени, но все же постепенным. Скорость эволюционных процессов в нем вполне можно измерить, и такие исследования, как мы видели, есть. Кембрий длился примерно 57 миллионов лет (542–485 млн лет назад), при этом в самом его начале (первые шесть миллионов лет) морская фауна была еще довольно бедна. Новые группы животных появились там действительно быстро, по меркам истории Земли, но вовсе не одномоментно.
С чем же все-таки это было связано? За полтора века, прошедших с тех пор, как ученые (и в том числе Чарльз Дарвин) осознали загадку кембрийского взрыва, предлагались разнообразнейшие объяснения этого события, от генетических до космических. Одна современная обзорная статья на эту тему так и называется — “По ту сторону кембрийского взрыва: от галактики до генома”. Например, тенденция к массовому образованию минеральных скелетов — знаменитая “скелетная революция”, она же “биоминерализация”, — в начале кембрия охватила не только самых разных многоклеточных животных, но и одноклеточных эукариот, и некоторые водоросли. Напрашивается предположение, что это было связано с глобальным изменением химического состава внешней среды, то есть в данном случае морской воды. И в самом деле, показано, что в начале кембрия по каким-то чисто геологическим причинам примерно в три раза выросла концентрация в морской воде кальция (Ca2+) — иона, который нужен для создания твердых скелетов как никакой другой. Минеральная основа скелетов животных — чаще всего карбонат кальция (раковины моллюсков, иглы и чашечки коралловых полипов, спикулы губок), а иногда фосфат кальция (кости позвоночных). Уже из этих названий ясно, что без кальция тут так или иначе не обойтись.
Проблема в том, что объяснить скелетную революцию еще не значит объяснить сам кембрийский взрыв. Скелетная революция всего лишь снабдила твердыми минерализованными тканями животных, уже существовавших к моменту ее начала. Причем это коснулось далеко не всех из них. В тех кембрийских местонахождениях, тип сохранности которых допускает захоронение бесскелетных существ, сразу же обнаруживается, что значительная часть кембрийской фауны так и осталась “мягкотелой”. Так что дело не в скелетах. Явление, которое надо объяснить в первую очередь, — это уникальное ускорение эволюции многоклеточных животных, очень быстро (за конец эдиакария и начало кембрия) создавшее множество новых крупных групп, неважно, скелетных или нет.
В дальнейшем рассказе мы будем исходить из сценария, который кратко изложил еще в начале 1970-х годов американский палеонтолог Стивен Стэнли. Конечно, тут надо помнить, что палеонтология — очень быстро развивающаяся наука. Работы 40-летней давности в ней всегда требуют поправок, и мы эти поправки по ходу разговора внесем. Правда, в данном случае это будут скорее дополнения. Главная идея Стэнли исключительно хорошо выдержала проверку временем. Сумма фактов, известных на данный момент, вполне в нее вписывается.
Начнем с начала. На полях заметим: решить, что именно стоит принять за “начало”, при разборе любого исторического процесса — задача непростая, ведь причинно-следственные цепочки могут тянуться в прошлое едва ли не до бесконечности, сбивая неосторожного исследователя с толку. В нашем случае “началом” будет эдиакарская биота. Что же она собой представляла?
В экологии принято выделять организмы-средообразователи, активность которых определяет собой структуру целых сообществ. Такие организмы называются эдификаторами. Например, в дубраве эдификатором является дуб, в маленьком тихом пруду им вполне может быть ряска, и т.д. Так вот, в эдиакарских морях эдификаторами были покрывавшие дно “ковры” нитчатых водорослей — так называемые водорослевые маты. На этих “коврах” жили уже знакомые нам вендобионты. Большинство из них вело прикрепленный образ жизни. Как они питались — не совсем понятно, но, скорее всего, осмотрофно, всасывая из воды растворенные вещества всей поверхностью тела. Таким способом до сих пор питаются некоторые морские протисты, например крупные — до 20 сантиметров! — многоядерные ксенофиофоры, похожие на гигантских амеб (они относятся к супергруппе Rhizaria). Вендобионты вполне могли быть близки к ним по образу жизни.
Впрочем, на эту тему есть и другие идеи. В 1986 году палеонтолог Марк Макменамин предположил, что вендобионты были экологическими аналогами современных погонофор — глубоководных кольчатых червей, лишенных рта и кишечника. Погонофоры живут в океане на такой глубине, куда солнечный свет не проникает. Зато там встречаются горячие источники, выделяющие в воду сероводород (H2S). Тело погонофоры набито симбиотическими бактериями, которые окисляют сероводород до серы и полученную при этом энергию используют для фиксации углекислоты, как при фотосинтезе. За счет этого процесса питаются и сами бактерии, и червь, в котором они живут. Некоторые погонофоры питаются за счет других бактерий, окисляющих не сероводород, а метан, но тоже автотрофных. В любом случае погонофоры — это фактически автотрофные животные. Что касается вендобионтов, то им было еще проще: они часто жили на мелководье, где солнечного света хватало для фотосинтеза, и вполне могли бы питаться за счет наполнявших их тело симбиотических одноклеточных водорослей. Это тоже реально, есть современные черви и моллюски, которые именно так и делают. Правда, у них этот способ питания обычно является дополнительным, но почему бы ему не стать и основным? Мир вендобионтов, где никто никого не ел, Макменамин назвал “садом Эдиакары”, с явной шуточной аллюзией на сад Эдема. Большой недостаток этой гипотезы в том, что ее пока трудно проверить. К тому же она заведомо не может распространяться на всех вендобионтов без исключения: некоторые из них жили в море глубже уровня, куда проникает достаточно света для фотосинтеза. Но в конце концов, в разных условиях они могли питаться по-разному.
Парадокс в том, что концепция “сада Эдиакары” представляется близкой к истине при любом сколько-нибудь реалистичном предположении о способе питания вендобионтов. Тут уже и неважно, жили в них водоросли или нет. В эдиакарском мире действительно никто никого не ел (не считая одноклеточных организмов). Принципиально важно, что в эдиакарских сообществах до некоторого момента не было не только хищников (которые питались бы другими животными), но и “травоядных” (которые умели бы механически соскребать водоросли). Таким образом, водорослевым матам там никто особо не мешал расти.
Все изменилось, когда повышение концентрации кислорода в морской воде (судя по геологическим данным, оно шло постепенно в течение всего эдиакария) позволило некоторым многоклеточным существам ускорить обмен веществ настолько, чтобы начать вести по-настоящему активный образ жизни. Тогда появились “сборщики урожая” — крупные животные со сложной двигательной системой и ртом, которые перемещались по водорослевым матам и выедали их значительные участки. Одним из таких “сборщиков” была знакомая нам кимберелла. По образу жизни и по скорости передвижения первые эдиакарские животные-водорослееды, скорее всего, приблизительно напоминали современных улиток. Для нас это выглядит безобидно, но с точки зрения эдиакарских жителей появление таких чудищ было настоящей катастрофой. Водорослевые маты немедленно перестали быть сплошными. К тому же животные не только соскребали их сверху, но и объедали снизу, освоив для этого проникновение в грунт (у зоологов принято называть такое поведение “минированием”). Тут досталось заодно и вендобионтам, которые в конце эдиакария просто исчезли.
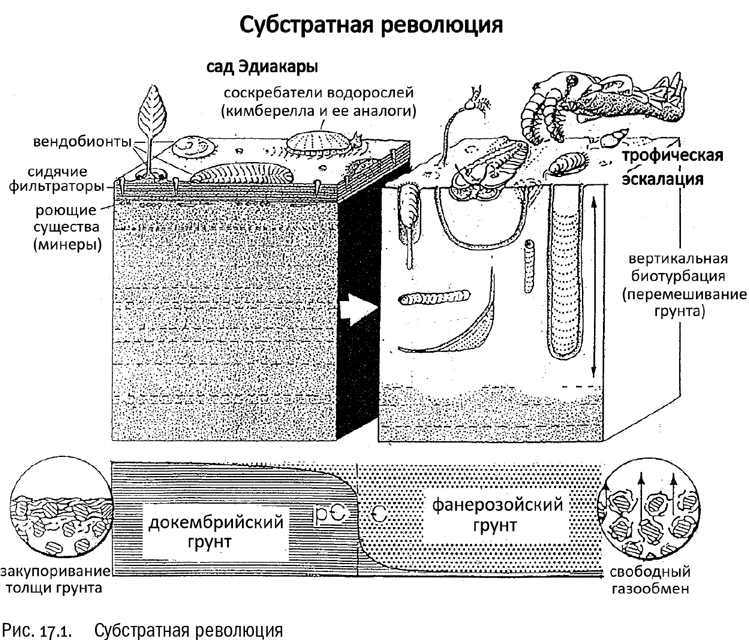
С этого момента начала работать общая закономерность, установленная экологами уже давно и проверенная разными способами вплоть до прямых экспериментов: в условиях давления хищника разнообразие его жертв повышается по сравнению с сообществом, где хищников нет вовсе. Если раньше донные сообщества заполнялись очень немногочисленными доминирующими видами водорослей, то теперь равновесие рухнуло и на этом фоне началась бурная эволюция. А между тем набор экологических ниш, доступных животным, тоже расширялся. Появились, например, активные грунтоеды, приспособленные к тому, чтобы постоянно жить в норах, пропуская донный грунт сквозь кишечник и извлекая из него питательные вещества. Так и до сих пор живут многие морские черви, вроде пескожила. Черви-грунтоеды впервые стали рыть в морском дне не только горизонтальные, но и вертикальные ходы, вызывая обогащение грунта кислородом и тем самым дополнительно облегчая его заселение другими животными (см. рис. 17.1). Эти события получили название субстратной революции. Таким образом, эволюционирующие животные не только занимали готовые экологические ниши, но и активно создавали новые, превращая процесс в автокаталитический (самоускоряющийся).
Некоторые жители поверхности дна стали расширять свои экологические ниши не в сторону грунта, а, наоборот, в сторону водной толщи. В результате возник зоопланктон — сообщество мелких животных, пребывающих взвешенными в воде и дрейфующих вместе с ней. Как правило, представители зоопланктона питаются, фильтруя воду и отцеживая из нее фитопланктон, то есть находящиеся в той же водной толще одноклеточные водоросли (а таких к моменту кембрийского взрыва было уже сколько угодно, на все вкусы). И действительно, в раннем кембрии в палеонтологической летописи появляются первые планктонные фильтраторы — жаброногие рачки. Жаброноги, как и все ракообразные, обладатели членистых конечностей, исходно предназначенных для хождения по грунту, то есть по дну. Поэтому нет никаких сомнений, что ранние этапы своей эволюции они прошли на дне, а к планктонному образу жизни обратились только после этого.
Последствия появления зоопланктона, в свою очередь, оказались вполне глобальными. Дело в том, что животные-планктеры отфильтровывают из воды не только водоросли, но и любую взвесь, в которой могут быть хоть какие-то питательные вещества. В основном это распыленные остатки мертвых организмов. Отфильтровав взвесь и всосав из нее полезные молекулы, планктеры (в первую очередь ракообразные) аккуратно “упаковывают” остальное содержимое кишечника в плотные комки — фекальные пеллеты, которые из-за своей компактности быстро тонут и отправляются на дно. А вода в результате очищается. Пеллетная транспортировка взвеси — важнейший фактор, понижающий мутность воды в океане.
Надо сказать, что переменивший облик Земли феномен пеллетной транспортировки непосредственно связан с одной чисто физиологической особенностью животных, образующих основную массу зоопланктона, — ракообразных. Дело в том, что находящаяся в кишечнике пища у них окружается перитрофической мембраной — тонкой белково-хитиновой пленкой, пропускающей мелкие молекулы, но отфильтровывающей крупные. Остатки переваренной пищи оформляются в компактные пеллеты именно благодаря ей. Перитрофическая мембрана есть и у насекомых, что неудивительно: ведь насекомые — это на самом-то деле всего лишь одна из многих эволюционных ветвей ракообразных . “В лице” насекомых ракообразные освоили сушу. Впрочем, тут мы забегаем вперед.
Итак, после появления планктонных фильтраторов вода в океане стала прозрачной, свет начал проникать в нее на большую глубину и повысилась концентрация кислорода (часть которого раньше расходовалась на окисление той же мертвой взвеси). Первый фактор сразу увеличил глубину зоны, в которой света достаточно для фотосинтеза, а второй улучшил условия для придонной фауны. По всем данным, прозрачный насыщенный кислородом фанерозойский океан резко отличается от мутного докембрийского океана. Заодно повысилась концентрация кислорода и в атмосфере. Естественно, что в новых благоприятных условиях разнообразие как растений, так и животных дополнительно выросло. Замкнулась еще одна автокаталитическая петля.
Пришествие хищника
Все животные, о которых мы до сих пор говорили, были “травоядными” — хотя бы в самом наиширочайшем смысле этого слова. Они питались или фотосинтезирующими организмами, или, на худой конец, чьими-то останками. При этом собственная биомасса “травоядных” представляла собой ценный (и до какого-то момента совершенно невостребованный) ресурс для животных, питающихся другими животными, то есть для хищников.
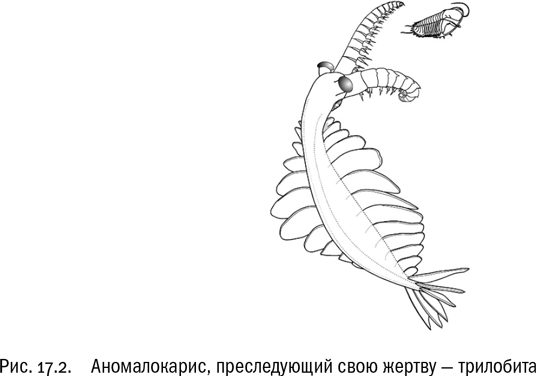
Поначалу никаких многоклеточных хищников просто не существовало. Но при наличии таких атрибутов активной жизни, как нервная система, мускулатура и ротовой аппарат, их появление было всего лишь вопросом времени. Первые крупные хищники, уже совершенно определенно специализированные на питании другими многоклеточными животными, появляются в летописи примерно 520 миллионов лет назад. Это динокариды — хорошо плавающие существа, родственные членистоногим. Самый известный представитель динокарид — аномалокарис, стройное сегментированное создание длиной около метра со сложными фасеточными глазами и мощными членистыми околоротовыми конечностями, явно служившими для захвата крупной подвижной добычи (см. рис. 17.2). В самом начале кембрия подобных хищников нет. Пресловутая “скелетная революция”, несомненно, в какой-то мере стала ответом на их появление — изменение химического состава морской воды только облегчило ее. А появление скелетов, в свою очередь, запустило освоение новых и новых экологических ниш.
Стивен Стэнли совершенно правильно писал, что для объяснения кембрийского взрыва вполне достаточно чисто биологических причин. Факторы, действующие на биосферу извне, могли повлиять на скорость того или иного процесса, но все главные события можно объяснить и без них. Вспышка разнообразия многоклеточных животных была естественным результатом серии автокаталитических процессов, запущенных появлением первых “травоядных” (вроде кимбереллы) и происходивших на уровне природных сообществ, то есть, иначе говоря, экосистем. Вне экологии понять кембрийский взрыв невозможно.
С появлением хищников процесс образования новых жизненных форм стал понемногу тормозиться. Репертуар экологических ниш сложился, почти все они уже были распределены и заняты. Разумеется, расширение сообществ продолжалось и дальше — просто медленнее. Например, только после окончания кембрийского периода появились лопатоногие моллюски, занявшие довольно-таки экзотическую нишу роющих хищников, преследующих своих жертв внутри грунта. Но такого размаха, как на рубеже эдиакария и кембрия, крупномасштабная эволюция животных больше никогда не достигала.
С точки зрения событийной истории началом кембрийского взрыва можно считать появление первых эффективных водорослеедов (кимберелла), а концом — появление первых эффективных хищников (аномалокарис). Кимберелла появилась 555 миллионов лет назад, аномалокарис — 520 миллионов лет назад, интервал между ними — 35 миллионов лет. Не так уж и быстро.
Первопроходцы суши
Выход жизни на сушу — событие, настолько сильно растянутое во времени, что его просто невозможно рассматривать как четкий порог. Более того, сама постановка вопроса о выходе на сушу на самом деле спорна. Утверждение “море — колыбель жизни” вовсе не настолько само собой разумеющееся, как может показаться.
Например, в последние годы стала популярной гипотеза, согласно которой жизнь возникла не в океане, а в мелких наземных водоемах. В пользу этой гипотезы есть несколько биохимических доводов, самый простой и наглядный из которых следующий. Известно, что цитоплазма практически всех живых клеток содержит гораздо больше ионов калия (K+), чем ионов натрия (Na+). Между тем почти во всех природных водоемах соотношение концентраций этих ионов в точности обратное. В морской воде в 40 раз больше натрия, чем калия, а в живой клетке, наоборот, в 10–20 раз больше калия, чем натрия. Внутриклеточный избыток калия важен для работы многих ферментов, и в том числе для системы синтеза белка. Причем анализ генных последовательностей показывает, что эти калий-зависимые ферменты очень древние. Скорее всего, они были уже у общего предка всех современных живых организмов. Значит, первые клетки формировались в среде, где калия было намного больше, чем натрия. Океан такой средой быть не мог. Гораздо вероятнее, что это были горячие источники, вода в которых как раз может иметь подходящий химический состав. И располагаться эти источники могли как в море, так и на суше (см. главу 11).
Но даже если живые клетки появились в неглубоких водоемах на суше, то после этого они быстро заселили океан. Дело в том, что по большинству параметров морская среда намного комфортнее наземной: в ней всегда хватает воды, и в нее почти не проникают опасные для клеток ультрафиолетовые лучи. Сейчас ультрафиолетовый компонент солнечного света экранируется слоем атмосферы, обогащенным газом озоном (O3), который постоянно образуется в небольшом количестве из атмосферного кислорода (O2). До кислородной революции на Земле не было никакого озонового слоя, поэтому защищать клетки от ультрафиолетовых лучей могла только вода. Получается, что океан был если и не колыбелью жизни, то уж точно ее убежищем на первые несколько миллиардов лет эволюции.
Первые признаки жизни на суше относятся еще к раннему докембрию. В основном это палеопочвы, обогащенные “легким” изотопом углерода 12C — как мы помним, именно по этому критерию биогенный углерод, прошедший по биохимическим путям живых клеток, отличается от абиогенного. Первые такие палеопочвы имеют возраст примерно 2,7 миллиарда лет, но не исключено, что они существовали и раньше. Это — архейская эпоха, когда на Земле жили одни прокариоты. Зная, насколько огромны экологические возможности прокариот, нетрудно допустить, что кто-то из них прижился и в таком негостеприимном месте, как архейская суша.
Первое прямое свидетельство наземной жизни — остатки нитчатых синезеленых водорослей в ископаемой пещере возрастом 1,2 миллиарда лет. Этот момент времени относится к эпохе “скучного миллиарда лет”, когда синезеленые водоросли, они же цианобактерии, уже не только были многочисленны, но и мало чем отличались от своих современных родичей. А о современных синезеленых водорослях известно, что они довольно часто живут вне воды, например на почве или на скалах. Так что никаких серьезных причин сомневаться в этой находке нет.
Несколько позже — один миллиард лет назад — появляются остатки наземных водорослей, предположительно относящихся уже к эукариотам. Они образуют пластинчатые колонии, а также цисты, то есть покоящиеся стадии с толстой сложно структурированной клеточной стенкой. Тут надо сказать, что проникновение эукариотных водорослей в наземную среду — явление, опять же, не столь и редкое. Например, современная зеленая водоросль трентеполия не просто выдерживает наземные условия, а специализирована к ним и ведет полностью наземный образ жизни. У нее есть целый набор приспособлений к жизни на воздухе — очень толстые клеточные стенки, обилие запасных веществ. Колонии трентеполии обычно выглядят как кирпично-красный или желтый налет на коре деревьев (зеленый цвет, присущий большинству растений, тут маскируется дополнительными красными пигментами, защищающими водоросль от “световых перегрузок” в слишком хорошо освещенной среде). В том, что водоросли с похожим образом жизни существовали миллиард лет назад, нет ровным счетом ничего невероятного.
Надо отметить, что оценки, которые мы сейчас обсуждаем, не самые смелые, а, наоборот, самые скептические. Например, древнейшая известная кора выветривания (понятие, частным случаем которого является почва) образовалась примерно 3,5 миллиарда лет назад, и есть исследователи, которые допускают существование наземной жизни уже тогда. Это не выглядит невозможным, но пока не имеет строгих доказательств. В чем мы можем быть уверены, так это в том, что наземная жизнь, причем многоклеточная, существовала еще до позднепротерозойской эпохи “Земли-снежка”. Неудивительно, что сразу после “Земли-снежка”, то есть в эдиакарском периоде, она возродилась.
И как возродилась! Есть гипотеза, что именно активностью наземной биоты объясняется “вторая кислородная революция”, то есть происходивший в эдиакарии рост содержания кислорода в атмосфере. Причинно-следственная цепочка тут вот какая. Важным фактором, влияющим на концентрацию кислорода, является присутствие в той же среде мертвого органического вещества. Если такого вещества много, то значительная часть кислорода тратится на его окисление, переходя в итоге в углекислоту и воду. Если любым способом вывести из оборота мертвую органику, свободного кислорода при прочих равных условиях станет больше. Именно это произошло, когда кембрийский зоопланктон освоил пеллетную транспортировку мелкой взвеси на дно. В эдиакарии механизм, приводивший к этому эффекту, тоже существовал, но был несколько другим. Мертвая органика связывалась частицами глины, которые поступали в океан с континентальным стоком, и оседала на дно вместе с этими частицами. Между тем геологи считают, что источником такого количества глины может быть только биогенная кора выветривания, то есть почва. Тогда получается, что эдиакарская наземная биота, предположительно состоявшая из водорослей и грибов, запустила уже достаточно мощное почвообразование. Кстати, есть данные, что в эдиакарии были и лишайники.
С другой стороны, в палеонтологии, как мы уже знаем, довольно широко распространена идея, что эволюция многоклеточных животных зависела от содержания кислорода в атмосфере (так называемая гипотеза кислородного контроля). Высокая концентрация кислорода, несомненно, облегчила переход к “настоящей” животной многоклеточности, требующей много энергии. Если же подъем этой концентрации произошел в конечном счете из-за активности наземных организмов — значит, выход жизни на сушу не только предшествовал появлению настоящих многоклеточных животных, но и был в какой-то мере его причиной.
Леса и насекомые
В первые четыре периода палеозойской эры — кембрий (542–485 млн лет назад), ордовик (485–443 млн лет назад), силур (443–419 млн лет назад) и девон (419–359 млн лет назад) — наземная биота постепенно, но неуклонно усложнялась. Высшие споровые растения, относящиеся примерно к тому же уровню организации, что и современные мхи, совершенно достоверно появились в ордовике. Наземные водоросли, от которых они, скорее всего, произошли, известны из кембрия. Ну а в конце ордовика уже существовали первые сосудистые растения, имеющие специальные проводящие ткани для транспорта воды и питательных веществ. К ним относятся все современные высшие растения, кроме мхов: папоротники, псилоты, хвощи, плауны, голосеменные и цветковые. Начиная с силура именно сосудистые растения преобладают в растительном покрове Земли.
Животные тоже вышли на сушу. В первую очередь это членистоногие — тип, который в наземных условиях достиг колоссального расцвета (см. рис. 17.3). В силуре появились многоножки и скорпионы, в девоне — сенокосцы, клещи, ложноскорпионы и насекомые. Причем на сушу некоторые из них выбрались довольно рано: например, ископаемые следы, наверняка принадлежащие наземным многоножкам, обнаруживаются в ордовике. А в начале девона появились хищные губоногие многоножки, которые могли питаться только другими членистоногими. У ископаемых экземпляров этих многоножек сохранились даже ногочелюсти с крючками, сквозь которые в тело жертвы впрыскивался яд из ядовитых желез — этот механизм у них одинаков с современными представителями группы. Это означает, что к началу девона сообщество наземных членистоногих было уже достаточно сложным, чтобы хищник всегда мог найти себе добычу.

Что касается позвоночных, то они вышли на сушу едва ли не последними. Наземные позвоночные появляются в палеонтологической летописи только во второй половине девона, причем их первые известные представители, скорее всего, еще вели в основном водный образ жизни. Расцвет их начался в следующем периоде — каменноугольном, или карбоне (359–299 млн лет назад). Освоившись на суше, наземные позвоночные заняли в сообществе место “суперхищников”, способных без особого труда съесть любое членистоногое, и приступили к собственной бурной эволюции.
В сложившемся “пазле” наземной фауны явно не хватает двух привычных нам фрагментов. Во-первых, там нет пауков. Их ископаемые родичи, еще не умевшие плести паутину, вышли на сушу в силуре, но настоящие пауки, и то очень примитивные, появились только в конце карбона. А во-вторых, мы пока еще не сказали ни слова о летающих насекомых. Упомянутые девонские насекомые были первичнобескрылыми, то есть относились к таким группам, у которых крыльев никогда не было. Первичнобескрылые насекомые есть и сейчас: например, встречающаяся в домах сахарная чешуйница. Но подавляющее большинство современных насекомых — крылатые (включая сюда и тех, кто по той или иной причине потерял крылья вторично). И вот этой группы в девоне еще нет. Первый бескрылый родственник крылатых насекомых, очень похожий на них строением ротового аппарата, обнаружен в конце девона. А первое насекомое с крыльями — в начале карбона. С этого момента в истории жизни на Земле началась новая эпоха, хотя поначалу, вероятно, это особенно заметно не было.
Дело в том, что крылатые насекомые во многих отношениях уникальны на фоне всей остальной земной фауны, вместе взятой. Достаточно сказать, что к этой группе относится примерно две трети всех современных видов животных. Кстати, наиболее богатый видами отряд крылатых насекомых — жесткокрылые, или жуки; с этим связано знаменитое (хотя и не имеющее четкого подтверждения в источниках) высказывание биолога Джона Холдейна: “Если Создатель существует, он, должно быть, необычайно любит жуков”. При всем этом крылатые насекомые обладают необычайно широким набором способов питания. Это наглядно выражено во многообразии их ротовых аппаратов: грызущий, лижущий, грызуще-лижущий, трубчато-сосущий, колюще-сосущий, режуще-сосущий и др., вплоть до таких необычных, как хватательная маска личинки стрекозы, гигантские жвалы жука-оленя или 20-сантиметровый спирально закрученный хоботок длиннохоботного бражника. Крылатые насекомые встречаются везде, где только могут жить многоклеточные существа, за исключением морских глубин. Они бывают хищниками, растительноядными, паразитами, поедателями грибов или разрушителями мертвой органики, и во всех этих ролях могут вырабатывать тончайшие пищевые специализации. Кроме того, они образуют еще и множество других, непищевых связей с соседями по сообществам: например, насекомые-опылители сильно повлияли на эволюцию цветковых растений. В общем, биосферную роль крылатых насекомых трудно переоценить.
Растительный мир на рубеже девона и карбона тоже изменился. В середине девона сразу несколько неродственных групп наземных растений выработали жизненную форму дерева с мощной корневой системой и вертикальным ветвящимся стволом высотой до восьми метров. А в позднем девоне появились первые леса, состоявшие в основном из древовидного папоротника археоптериса. И хотя археоптерисовые леса конца девона не пережили, другие древесные породы тут же пришли им на смену. Например, в карбоне появились те самые сигиллярии с “упругими и рыхлыми стволами”, которые упомянул в стихах поэт-акмеист Михаил Зенкевич. В общем, на рубеже девона и карбона началось великое “облесение суши” (afforestation of the land). Лес открыл множество новых экологических ниш, которые не замедлили занять животные, питавшиеся как растениями, так и друг другом. И тот же лес выбрасывал в атмосферу огромное количество кислорода, тем самым облегчая животным всевозможные “эксперименты” с формой и размером. В карбоне достигли своего расцвета амфибии и появились первые рептилии — несомненно, в связи с ростом разнообразия насекомых, которыми эти позвоночные могли питаться. Так пошло и дальше. Крылатые насекомые и леса — два фактора, вот уже примерно 350 миллионов лет определяющие облик земной суши.
Парк пермского периода
Последний период палеозойской эры — пермский — продолжался 46 миллионов лет (298–252 млн лет назад). Пермский мир был уже во многих отношениях близок к нынешнему. Сушу населяли многообразные наземные позвоночные, среди которых были и хищники, и растительноядные. Процветал мир насекомых; именно в перми появился отряд, оказавшийся самым эволюционно успешным в этом классе, — жуки. На континентах росли леса, состоявшие в Северном полушарии из древних хвойных, а в Южном — из лиственных голосеменных, отдаленно родственных современному гинкго. Водная жизнь тоже бурно развивалась: пермские моря были полны моллюсков, иглокожих, брахиопод, ракообразных, кораллов и других беспозвоночных, не говоря уж о рыбах. Современного наблюдателя (если бы он не слишком всматривался) в перми поразило бы, прежде всего, отсутствие птиц, млекопитающих и цветковых растений, а если бы он мог увидеть Землю из космоса — то еще и совершенно другое расположение материков.
В конце карбона — начале перми произошло так называемое гондванское оледенение, охватившее значительную часть суши Южного полушария. Очень вероятно, что его главной причиной было распространение лесов. Деревья карбонового периода связывали огромное количество углерода в биомассе своих стволов, разлагать которую было некому: эффективные разрушители древесины тогда еще не возникли. В результате деревья падали и захоранивались как есть, создавая залежи каменного угля. Переход углерода из атмосферного углекислого газа в эти залежи, где он оказался “заперт”, сильно ослабил парниковый эффект — вот этого, по-видимому, и хватило для запуска оледенения. Достоверно показано, что атмосферная концентрация CO2 в тот момент сильно упала. Гондванское оледенение создало холодный климат на части земной суши, но никаких биосферных переворотов, насколько можно судить, не вызвало. Все крупные группы организмов его спокойно пережили.
События конца перми были гораздо драматичнее. Но чтобы к ним перейти, нам понадобится небольшое вступление.
Пять великих рубежей
Еще палеонтологи XIX века прекрасно знали, что рубежи периодов, а тем более эр, обычно характеризуются спадами разнообразия живых организмов, то есть, попросту говоря, вымираниями. Это естественно: одни живые существа вымирают, другие приходят им на смену. Границы периодов как раз и проводятся по моментам, когда смены флор и фаун особенно заметны. А конец пермского периода одновременно является концом всей палеозойской эры — не приходится удивляться, что некоторая часть биоты на этом рубеже исчезла.
А что получится, если оценить это явление количественно? В 1979 году палеонтолог Дэвид Рауп, много занимавшийся статистическим анализом разнообразия древних фаун, опубликовал статью со следующим выводом: есть основания считать, что в конце пермского периода вымерло, не оставив потомков, примерно 96% всех видов морских животных на Земле. Получалось, что биота, по крайней мере морская, прошла в этот момент через настоящее “бутылочное горлышко”. Подсчеты Раупа были основаны на огромном материале. И хотя его результаты касались только морской фауны, они не оставляли сомнений, что в конце перми действительно случилось очень крупное вымирание.
Другой известный палеонтолог, Дуглас Эрвин, решил определить скорость этого процесса. Анализируя вместе с коллегами распределение остатков животных в осадочных толщах, он постепенно пришел к выводу, что пермское вымирание произошло за очень короткий срок — меньше одного миллиона лет. Коротким этот срок, конечно, можно назвать только по меркам истории Земли, но для событий такого масштаба он действительно мал.
Итак, на границе палеозоя и мезозоя произошло нечто большее, чем рядовая смена ископаемых фаун. Рауп и Эрвин столкнулись с явлением, относящимся к категории массовых вымираний (mass extinctions). Что же это, собственно, такое?
Массовые вымирания, они же биосферные кризисы, случались в истории Земли довольно регулярно. Принято считать, что в фанерозое крупнейших вымираний было пять. Пермское — одно из них. Из остальных четырех два произошли внутри палеозоя (в конце ордовика и в конце девона), одно — внутри мезозоя (на рубеже триасового и юрского периодов), и еще одно отделяет мезозой от кайнозоя (именно тогда исчезли динозавры). Некоторые из этих событий вполне могли иметь внешние причины. Например, вымирание на границе мезозоя и кайнозоя связывают с падением на Землю 10-километрового астероида, кратер от которого сейчас находится на побережье полуострова Юкатан. Но похоже, что если это и верно, то случай тут все же исключительный: большинство вымираний объяснить подобными единичными катастрофами не удается. Необходимо учитывать, что кризисов, лишь немного уступающих пяти “главным” вымираниям, а то и сравнимых с ними, в фанерозое на самом-то деле было еще больше десятка. Космических катастроф просто не хватит на них на всех.
Некоторые палеонтологи считают, что кризис, сопровождающийся массовым вымиранием, есть совершенно нормальная стадия развития природного сообщества, во всяком случае, сообщества размером с земную биосферу. Известный палеоботаник Валентин Абрамович Красилов так и писал: кризис — это естественный механизм эволюции сообществ. Под этим углом зрения становится понятно, почему некоторые кризисы не удается толком связать ни с какими внешними воздействиями на живую природу. Но столь же ясно, что само по себе признание кризиса нормальным явлением еще ничего не объясняет. В основе любого исторического события, и биосферного кризиса в том числе, лежат вполне реальные причинно-следственные связи. Наша задача — их раскрыть. Если нет внешних причин, значит, есть внутренние, и наоборот.
В этом плане вымирание на границе палеозоя и мезозоя особенно интересно. Чтобы понять почему, его можно бегло сравнить с вымиранием на границе мезозоя и кайнозоя, тем самым, которое погубило динозавров. Прежде всего уточним названия этих событий. Последний период мезозоя называется меловым, первый период кайнозоя — палеогеновым, поэтому “динозавровый” кризис мы будем называть мел-палеогеновым, как это обычно и делают. Кризис на рубеже палеозоя и мезозоя по тому же принципу называют пермо-триасовым.
Разница между этими кризисами следующая. Во-первых, пермо-триасовое вымирание было намного мощнее. Аккуратный подсчет по одной и той же методике показывает, что мел-палеогеновый кризис уничтожил примерно 15% от всех существовавших тогда биологических видов, а пермо-триасовый — примерно 60%. Эти цифры несколько меньше тех, которые обычно приводятся в научно-популярных публикациях, но зато уж в них можно не сомневаться. Во-вторых, в отличие от мел-палеогенового кризиса, пермо-триасовый кризис не был вызван никаким внешним ударом вроде астероидного импакта (гипотеза, что подобный импакт был и тут, высказывалась, но не подтвердилась). И в-третьих, после пермо-триасового кризиса разрушенная биосфера вступила в очень долгий — растянувшийся на десятки миллионов лет — период постепенного восстановления, чего после мел-палеогенового кризиса не было вообще: там сообщества восстановились сразу.
Недавно детальные геологические исследования показали, что пермское вымирание заняло гораздо более короткий срок, чем думали раньше, — всего 60 000 лет. Это 0,06 миллиона лет, срок по меркам планетной истории просто невероятно маленький. Что же там случилось?
Пермский кризис
Большинство ученых сейчас считает, что пермскую катастрофу вызвали вулканы. Показано, что точно в момент биосферного кризиса (то есть 252 миллиона лет назад) на территории, которая сейчас называется Западной и Средней Сибирью, образовалась трапповая провинция — система действующих вулканов, не ограниченных точечными кратерами, а занимающих сплошное протяженное поле. Выглядело это, должно быть, грандиозно. Расколотая во многих местах земная кора извергла буквально море расплавленного базальта, залившее 1,6 миллиона квадратных километров — это площадь современного Ирана. Текущий базальт затопил огромную территорию и постепенно застывал, создавая абсолютно плоскую безжизненную равнину, по которой текли новые базальтовые волны от новых извержений. А дым от них, надо полагать, затмил небо. Но все это было только прологом ко всепланетной катастрофе.
Трапповые вулканы неизбежно выбрасывали в атмосферу колоссальные объемы вулканических газов — углекислого газа (CO2), хлороводорода (HCl), сероводорода (H2S), сернистого ангидрида (SO2) и др. Это имело много последствий. Во-первых, углекислый газ усилил парниковый эффект, вызвав сильное глобальное потепление. Во-вторых, вулканические газы повысили кислотность морской воды, а это по чисто химическим причинам сильно затруднило жизнь кораллов и иглокожих: карбонаты, из которых состоят твердые скелеты этих животных, в слишком кислой воде просто-напросто растворяются. В-третьих, сероводород ядовит для большинства живых организмов, особенно при концентрации, в тысячи раз превышающей нынешнюю атмосферную, — а именно такой уровень сероводорода создавали трапповые вулканы.
Более того, показано, что существует концентрация сероводорода, которая еще не мешает росту фитопланктона (то есть одноклеточных водорослей), но уже мгновенно убивает зоопланктон (например, веслоногих рачков). Если такое случится в реальном водоеме, он тут же “зацветет”. Дальше последует цепочка событий, обычная для цветущих водоемов и в наше время: бесконтрольно размножающимся водорослям не хватит ресурсов, часть из них погибнет, погрузится на дно и там начнет разлагаться, связывая запас кислорода в нижних слоях воды, который в данном конкретном случае должен быть и без того невелик: ведь чем выше температура, тем хуже кислород растворяется в воде. Если этим водоемом будет Мировой океан, то его глубины в результате станут практически бескислородными, что, собственно, в момент пермского кризиса и наблюдалось. Называя вещи своими именами, там случился глобальный замор.
Между тем в бескислородной среде, богатой соединениями серы, прекрасно себя чувствуют анаэробные бактерии, в том числе и сульфатредуцирующие, выделяющие в качестве побочного продукта метаболизма опять же сероводород. Готово — петля положительной обратной связи замкнулась. И наверняка подобных петель было несколько. Вулканических газов могло и не хватить, чтобы убить все живое, но их хватило, чтобы запустить серию обратных связей, сделавших катастрофу саморазвивающейся.
Пустынная планета
Сразу после пермского кризиса Земля представляла собой довольно унылое место. Ничем не сдерживаемый парниковый эффект привел к разогреву Мирового океана до 38–40 ºС. Это близко к смертельной для большинства животных и растений температуре денатурации белков (примерно 45º). К тому же при высокой температуре снижается растворимость кислорода в воде, так что существование водной фауны практически исключается. Палеонтологические данные показывают, что рыбы в эту эпоху остались только в приполярных областях океана. Суша, скорее всего, прогревалась еще сильнее, поэтому тропические и субтропические широты были непригодны для жизни крупных наземных животных. Авторы, опубликовавшие эти данные, делают твердый вывод: в раннем триасе температура стала дополнительной причиной массового вымирания.
Не блестяще обстояло дело и с кислородом. В перми его концентрация в атмосфере поднялась почти до 30%, а после пермского кризиса она упала до 10–15% и оставалась такой большую часть триаса. Это сильно ограничивало физиологические возможности уцелевших позвоночных, не давая им достичь крупных размеров. Полностью исчезли леса — деревьев, способных их образовать, просто не осталось нигде на Земле. Соответственно, вымерли все животные, занимавшие специфически лесные экологические ниши. Самым крупным и многочисленным наземным позвоночным этой эпохи был листрозавр — всеядная зверообразная рептилия размером со среднюю собаку. Впрочем, еще чаще листрозавра сравнивают со свиньей, подразумевая, что он вел похожий образ жизни и питался чем попало. Участвуя в подготовке телепередачи о листрозавре, английский палеонтолог Майкл Бентон назвал ее “Когда свиньи правили Землей”.
В целом можно утверждать, что больше всего от пермо-триасового кризиса пострадали морские беспозвоночные, а меньше всего — насекомые. У последних вообще все “провалы” разнообразия, вызванные массовыми вымираниями, сглажены по сравнению с другими группами животных. Частично это связано с их удачной анатомией и физиологией (дыхательная система насекомых состоит из микроскопических трубочек — трахей, которые пронизывают тело, доставляя воздух прямо к клеткам без всякого переноса кровью; как бы мало кислорода в этом воздухе ни было, животное его получит), а частично — с тем, что наземная среда, к которой они приспособлены, гораздо более разнообразна и расчленена, чем водная. Если вы — жук, то уж какое-нибудь приемлемое для себя микроместообитание всегда найдете.
Триасовое возрождение
На протяжении триасового периода структура сообществ постепенно восстанавливается. Более того, в ней возникают принципиально новые жизненные формы — например, морские рептилии. В середине триаса появляются двукрылые насекомые (то есть комары и мухи), а в конце — черепахи, крокодилы, динозавры, летающие ящеры (птерозавры) и млекопитающие. Это уже почти современная биота. Из групп, представителей которых мы привыкли каждый день видеть, не хватает только птиц (появляются в юре) и цветковых растений (появляются в мелу). Никаких переворотов, сравнимых с кембрийским взрывом или пермским кризисом, в истории Земли отныне больше не будет, по крайней мере до появления человека.
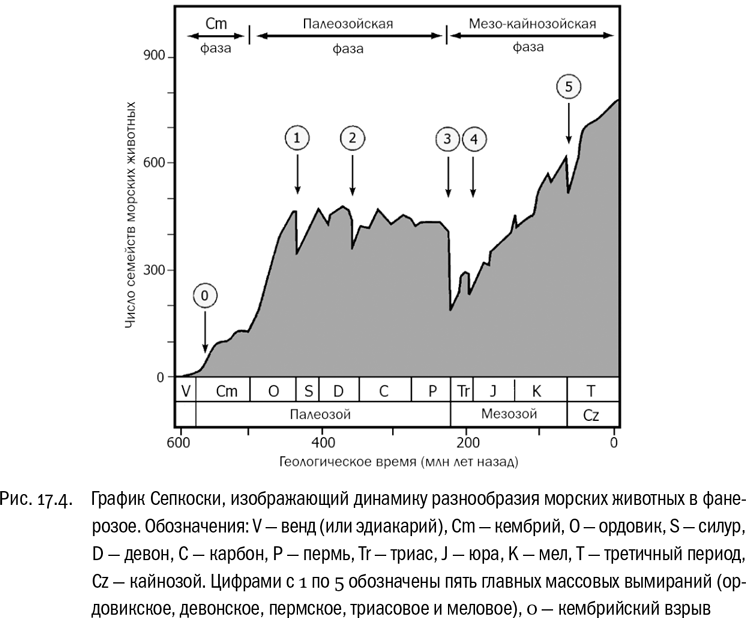
Тут уместно привести интереснейший чисто эмпирический результат, который получил американский палеонтолог Джон Сепкоски. Он собрал данные по морским животным за все времена — от кембрия до современности — и построил график, где по горизонтали было отложено время, а по вертикали — разнообразие, измеренное числом родов и семейств (см. рис. 17.4). Оказалось, что в течение всего палеозоя, не считая начального периода роста сразу после кембрийского взрыва, разнообразие остается в целом стабильным. После кризисов оно просто возвращается к прежнему уровню. На границе перми и триаса, естественно, виден глубокий провал. И что же дальше? За пермо-триасовым провалом следует не возвращение к прежнему устойчивому уровню, а непрерывный подъем, который иногда притормаживается очередными кризисами, но тем не менее неуклонно длится вот уже почти 250 миллионов лет. На графике Сепкоски очень наглядно видно, что вся эволюция как бы делится на две части: до пермского кризиса, когда разнообразие было по большому счету стабильным, и после него, когда оно непрерывно растет. Никакого общепринятого объяснения этому до сих пор нет, но сам факт установлен твердо. И получается, что пермский кризис изменил весь режим развития жизни на Земле. Перефразируя знаменитого английского историка Арнольда Тойнби, вполне можно сказать: эволюция — это ответ на вызов.
От клетки к социуму
Сумма доступных нам сейчас научных знаний, в общем, не оставляет сомнений, что одним из самых важных процессов в эволюции эукариот было слияние организмов. Оно шло на разных уровнях. Уже самая первая эукариотная клетка, скорее всего, возникла в результате симбиоза археи с бактерией. На следующем эволюционном этапе, который наступил довольно быстро, эукариотные клетки начали всевозможными способами объединяться друг с другом. Интересно, что генетическая близость клеток, вступающих в союз, была при этом моментом хоть и важным, но не решающим. В общем случае клетки, составляющие единый организм, могут быть как строго генетически одинаковыми (в типичном многоклеточном теле), так и вовсе не родственными друг другу (например, в лишайнике, где клетки гриба самым натуральным образом порабощают клетки зеленой водоросли). Преобладание в нынешней биоте организмов “первого типа”, каждый из которых возникает из одной-единственной яйцеклетки, — не сам собой разумеющийся факт, а поворот эволюции, требующий объяснения. Но так или иначе, не позже чем к началу палеозойской эры Земля стала миром многоклеточных эукариот — растений и животных.
На новом эволюционном витке началось объединение целых организмов в системы следующего уровня — социальные. Довольно точные эквиваленты слова “социальность” — “общественность”, или, в устаревшей лексике, “общежительность”. Любая социальность основана на обмене сигналами между особями, в результате которого у них формируется некоторое общее поведение. Например, социальные амебы, относящиеся к так называемым клеточным слизевикам (из супергруппы Amoebozoa), могут обмениваться химическими сигналами в виде молекул циклического аденозинмонофосфата (цАМФ, см. главу 8). Под действием этого сигнального вещества они сползаются вместе и образуют грибообразное плодовое тело. Это один из простейших случаев социальности, часто рассматриваемый как ее элементарная модель. Но ясно, что канал для обмена сигналами у слизевиков очень узкий. Примерно так же обстоит дело у растений, сигнальные системы которых почти исключительно химические и передают мало информации. Тут, однако, вспоминается знаменитый роман Джона Уиндема “День триффидов”, в котором хищные растения-мутанты научились общаться звуковыми сигналами, тем самым резко расширив канал передачи информации и создав сложное, организованное поведение. Нетрудно догадаться, какая группа организмов получила подобные возможности не в фантастике, а в реальности. Это — животные.
Качественно новым эволюционным достижением была не столько социальность животных как таковая, сколько ее высшая форма — эусоциальность, при которой “представители вида живут группами, состоящими из нескольких поколений, и члены группы действуют альтруистично по отношению друг к другу в соответствии с регулярным разделением труда”. Животные, ставшие эусоциальными, фактически объединяются в суперорганизм, члены которого больше не являются самодостаточными (не могут существовать без поддержки товарищей по виду и сами вынуждены тратить ресурсы на такую поддержку), но обретают взамен новые возможности, принципиально недоступные одиночной особи. Согласно процитированному определению, человек разумный — эусоциальный вид. “В этом смысле люди вполне сравнимы с муравьями, термитами и другими эусоциальными насекомыми” (там же). И есть некоторые основания полагать, что именно эусоциальность создала человека разумного — во всяком случае, в большей степени, чем любая другая его биологическая особенность.
Рассмотренное определение эусоциальности взято из относительно недавней книги Эдварда Уилсона, крупнейшего в мире специалиста по муравьям и глубокого мыслителя, в свое время фактически создавшего науку социобиологию. Тут возникает загвоздка, которую мы не можем обойти вниманием. Дело в том, что традиционно биологи называли эусоциальными только тех животных, у которых есть разделение на размножающуюся касту и стерильных рабочих. Идеал эусоциальности — это муравейник, где яйца откладывает только матка. Причем раньше так считал и сам Эдвард Уилсон. В своей обзорной работе 1975 года он перечисляет три признака эусоциальности: (1) общественная забота о потомстве, (2) разделение особей на размножающиеся и стерильные касты и (3) перекрывание как минимум двух поколений. Очевидно, что в случае с человеком камнем преткновения служит второй признак. Общественная забота о потомстве и перекрывание поколений у него есть, а вот разделения особей на размножающихся и стерильных — нет (если не считать бездетных элит вроде духовенства некоторых религий и китайских чиновников-евнухов, но наличие таких элит никак нельзя считать всеобщей чертой нашего вида).
Спустя 30 лет, в 2005 году, Уилсон с соавтором выпустили обзорную статью о муравьях, в которой дали следующее определение: “Эусоциальность — это эволюционно достигнутый уровень колониального существования, при котором взрослые члены колонии относятся к двум или более перекрывающимся поколениям, сотрудничают в уходе за молодью и делятся на размножающуюся и неразмножающуюся (или, по крайней мере, менее размножающуюся) касты”. Как видим, тут уже появилась оговорка. Кроме того, в этом же обзоре подчеркивается, что главное в явлении эусоциальности — это “общие принципы, существующие на организменном и сверхорганизменном уровне”, “параллели, ясно прочерченные между построением организма из молекул и тканей и суперорганизма — из взаимодействующих целостных организмов”. А в конце статьи упоминается и человек. Авторы признают, что путь его эволюции имеет много общего с эусоциальными животными, но прямо назвать человека эусоциальным существом еще пока не решаются.
Однако в том же 2005 году вышла статья, озаглавленная “Новое эусоциальное позвоночное?”. Ее авторы, английские биологи Кевин Фостер и Фрэнсис Рэтникс, обратили внимание на одну всем известную особенность человека — менопаузу. Это генетически запрограммированная физиологическая перемена, имеющая отчетливо пороговый характер (в отличие от старения, которое идет медленно и постепенно). И она приводит к тому, что все особи одного из двух полов выключаются из размножения примерно в середине жизни, при этом сохраняя свою социальную функцию — участие в выращивании потомства. Вероятнее всего, менопауза — это адаптация, появление которой было как-то связано с социальной структурой наших предков. Здесь Фостер и Рэтникс отмечают, что свойственная человеку полная и необратимая стерильность значительной части особей, ухаживающих за потомством, — это признак, вообще уникальный среди позвоночных. У голых землекопов — грызунов, которых часто считают образцом эусоциальности, — такого нет, стерильность рабочих особей у них физиологически обратима. Таким образом, если можно считать эусоциальным видом голого землекопа, то можно и человека разумного.
При всей спорности этой аргументации она, очевидно, дала нужную подсказку Эдварду Уилсону, мысли которого и без того много лет развивались в сходном направлении. В статье, опубликованной им вместе с соавторами в 2010 году, уже прямо сказано, что существует широкое определение эусоциальности (наличие “частично неразмножающихся каст”), под которое человек подходит. К этому времени Уилсон стал убежденным сторонником расширенной интерпретации эусоциальности: главное в ней — не распределение репродуктивных функций (как считали когда-то первые социобиологи), а возникновение сверхорганизма, “часто причудливого по устройству, но представляющего особый уровень биологической организации” (оттуда же). И вот в такое понимание эусоциальности человек уже вписывается. В 2012 году Уилсон выпустил замечательную книгу “Социальное завоевание Земли” (в русском переводе, вышедшем в 2014 году, — “Хозяева Земли”), целиком посвященную тому, какую роль в эволюции жизни на Земле сыграли эусоциальные существа, включая человека.
Муравьиными тропами
Именно на работах Эдварда Уилсона и будет преимущественно основан наш краткий рассказ про эусоциальность. Уилсон — идеальный проводник по этой теме, независимо от того, согласны ли мы с его общефилософскими взглядами (которые он решительно высказывает в нескольких своих последних книгах). И действительно, кто может разбираться во всем этом лучше, чем человек, посвятивший 60 лет своей жизни изучению муравьев?
Итак, первое, что можно констатировать: эусоциальность часто дает ее обладателям огромный эволюционный успех. Во многих тропических лесах две трети насекомых — это муравьи. Скоординированная активность множества муравьев или термитов позволяет построить огромное защищенное поселение с собственным микроклиматом, организовать походы на фуражировку и даже создать аналог сельского хозяйства (и среди муравьев, и среди термитов есть виды, разводящие в гнездах строго определенные грибы по достаточно сложным технологиям). Ясно, что одиночным насекомым такое не под силу.
Тем удивительнее, что эусоциальность появилась в эволюции относительно поздно. В палеозое неизвестны никакие следы активности эусоциальных животных — судя по всему, такого феномена тогда просто не существовало. Насколько мы знаем, первыми на Земле эусоциальными существами стали термиты. Самые ранние известные остатки термитов относятся к началу мелового периода. При этом меловые термиты очень похожи на современных — настолько, что неспециалист не смог бы их отличить, и, что еще важнее, они обнаруживаются на нескольких разных континентах, которые в меловом периоде уже отделены друг от друга океаном. Единым целым эти континенты были только в триасе. Так что, скорее всего, первые термиты произошли от своих тараканообразных предков не в мелу, а значительно раньше. Специалисты помещают вероятное время их возникновения где-то между средним триасом и ранней юрой, в интервале 237–174 миллиона лет назад. Получается, что первые общественные насекомые заняли (или создали) одну из принципиально новых экологических ниш, открывшихся после пермо-триасовой “перезагрузки” биосферы.
Несколько позже, но тоже в мезозойскую эру, появляются общественные перепончатокрылые насекомые — муравьи, пчелы, шмели, некоторые осы. На их развитие сильно повлияла так называемая “цветковая революция”, когда голосеменные растения в большинстве сообществ массово сменились недавно возникшими цветковыми — родственниками привычных нам магнолий, буков, роз и других деревьев, трав и кустарников. Это произошло примерно в середине мелового периода, и как раз тогда же началась очень бурная эволюция муравьев, в итоге сделавшая их одними из самых заметных членов наземных экосистем. Конечно, это не случайное совпадение. “Цветковые” леса обладают более сложной структурой местообитаний, в которую разным насекомым, в том числе и общественным, оказалось легче встроиться.
В масштабе эволюционного древа эусоциальность — в общем-то нечастое явление. Эдвард Уилсон и его соавторы пока насчитывают всего 18 независимых событий возникновения эусоциальности у животных (не считая человека), 16 из которых относятся к членистоногим. Эусоциальность возникла один раз у термитов, девять раз у перепончатокрылых (ос, пчел, муравьев), три раза у других насекомых и три раза у ракообразных. При этом во всех без исключения известных случаях необходимым предварительным условием для эусоциальности было защищенное гнездо, в котором постоянно живет и размножается группа особей одного вида. Эусоциальные морские ракообразные — десятиногие раки-щелкуны — умудряются создавать такое гнездо даже внутри тела другого животного, а именно губки. Только на основе постоянного гнезда может начаться разделение функций, при котором одни особи размножаются, а другие занимаются исключительно добычей пищи, уходом за личинками или защитой от внешнего мира. И тогда группа превращается в сверхорганизм.
У позвоночных животных эусоциальность возникает очень редко. Самый известный пример эусоциального позвоночного — африканский грызун под названием голый землекоп (Heterocephalus glaber). Голые землекопы ведут роющий образ жизни. Они создают подземное защищенное гнездо, в котором живет группа, представляющая собой расширенную семью с “маткой” (размножающейся самкой), ее двумя-тремя “мужьями” и несколькими десятками “рабочих” обоих полов, которые не размножаются, пока это делает “матка”. Такой уровень эусоциальности не уступает тому, которого достигли термиты.
У голых землекопов есть еще два необычных признака. Во-первых, они не стареют (смертность не растет с возрастом) и могут жить до 30 лет и больше — для млекопитающих такого размера это уникальный случай. Во-вторых, они не способны самостоятельно поддерживать постоянную температуру тела, то есть фактически не являются теплокровными животными. Последнее очевидным образом вызвано колониальной жизнью землекопов: температура в хорошо устроенном подземном поселении достаточно стабильна, чтобы физиологические механизмы терморегуляции можно было и отключить. Причины “отключения” старения гораздо более загадочны (одно из самых простых объяснений, хотя и не обязательно самое верное, таково: у землекопов любая рабочая самка в принципе может дождаться своего шанса занять место царицы, но для этого ей нужно как можно дольше прожить, сохранив силы, — отсюда естественный отбор против старения), но в любом случае они наверняка так или иначе связаны с необычной социальной структурой этих зверей.
Второй вид стопроцентно эусоциальных млекопитающих — дамарский землекоп (Fukomys damarensis). Если голый землекоп живет в Восточной Африке, то дамарский — в Юго-Западной. Эусоциальность сложилась у этих двух близких видов, скорее всего, независимо, так же, как она несколько раз независимо возникала у близких видов ос и пчел. Вот, по сути, и все эусоциальные позвоночные, не считая человека. Правда, к эусоциальности приближаются некоторые грызуны (прерийные полевки) и хищники (гиеновые собаки), но с голыми землекопами их по глубине приспособлений нельзя и сравнивать. В любом случае подавляющее большинство обладателей эусоциальности — это насекомые.
Эусоциальный мозг
Чем же отличается эусоциальность человека от эусоциальности любых других животных? Начнем с того, что в ней общего. Это склонность объединяться в устойчивые группы на стоянках. Тут стоит привести длинную цитату из Эдварда Уилсона, она лучше любого пересказа своими словами:
“Стоянки a priori были важнейшей адаптацией на пути к эусоциальности: по сути, стоянки — это человеческие гнезда. Все без исключения животные, достигшие эусоциальности, начинали со строительства гнезда и защиты его от врагов. Они выводили в нем потомство, покидали его, уходя на поиски пищи, и возвращались в него с добычей, которую делили с остальными обитателями... Почему защищенное гнездо играет такую важную роль? Потому что в нем члены группы вынуждены собираться вместе. Им приходится уходить на разведку и поиски пищи, но в конце концов они всегда возвращаются”.
Именно плотная “упаковка” разновозрастных и разнополых особей в компактном гнезде, которое нельзя покинуть без риска для жизни, вынуждает животных совершенствовать систему взаимодействий друг с другом. В истории человечества таким “гнездом” мог быть и костер, вокруг которого собиралась группа степных охотников, и пещера, и рыцарский замок, и даже советская коммунальная квартира. Этот фактор действовал на человека с самого начала, так же как он действует, например, на каких-нибудь социальных ос (кстати, именно от ос произошли муравьи).
И вот тут мы упираемся в принципиальные отличия человеческой эусоциальности от той, которая свойственна насекомым. Их, по мнению Уилсона, всего два: ограниченная способность к расселению и слишком крупный размер особей.
На расселение общественных насекомых очень сильно влияет их главное преимущество — крылья. Оплодотворенная муравьиная матка может легко пролететь расстояние, которое для человека (с поправкой на размер тела) было бы эквивалентно нескольким сотням километров. После этого она обламывает крылья и основывает новую колонию на новом месте, где, если повезет, может не оказаться никаких конкурентов. Следует добавить, что эта возможность стремительного расселения реализуется не всегда: например, у рыжих лесных муравьев гнезда обычно образуются “почкованием”, рядом со старыми, соединяясь с ними поначалу обменными дорогами. Но так или иначе расселение путем дальних перелетов муравьям в принципе доступно. А млекопитающим, во всяком случае наземным, — нет. Они могут расселяться только очень постепенно, не имея “предохранительного клапана”, позволяющего за один шаг разделить родственные колонии в пространстве и тем самым снизить напряженность конкуренции между ними. Это касается как голых землекопов, так и людей, и имеет у них одни и те же последствия, а именно высокую внутривидовую агрессивность. Показано, что голые землекопы отличаются “ксенофобией” и часто нападают на незнакомых — а значит, заведомо принадлежащих другому гнезду — особей своего вида. В отношении человека разумного тут можно обойтись без пояснений. В фильме Мамору Осии “Небесные скитальцы”, который уже упоминался совсем по другому поводу в главе 14, недаром звучат слова: “Нельзя уничтожить войну, не уничтожив человека”. В самом этом утверждении можно усомниться, но вот в том, что склонность к войне запечатлена в нашей чисто биологической природе, особых сомнений нет.
Не менее важен размер тех особей, которые образуют колонию. Млекопитающие — гиганты животного мира. Типичное современное млекопитающее превосходит массой типичное современное насекомое примерно на три порядка, то есть в тысячу раз. Это не абсолютный закон (некоторые млекопитающие бывают меньше некоторых насекомых), но — повторимся — типичное соотношение именно таково. Что же касается человека, то его размер очень велик даже по стандартам большинства млекопитающих: среди них на самом деле не так уж и много животных, вес которых превосходит килограмм, а тем более измеряется десятками килограммов. В эпоху динозавров, то есть в мезозое, таких зверей не существовало вовсе, но и в кайнозое их доля никогда не была особенно велика. Достаточно сказать, что две трети всех современных видов млекопитающих — это грызуны и летучие мыши.
Почему это важно? Очевидно, что чем крупнее животное, тем крупнее будут и его отдельные органы, в том числе мозг. Любой мозг состоит из более-менее однотипных нервных клеток — нейронов, размер которых у всех позвоночных в первом приближении одинаков. Чем мозг больше, тем большее число нейронов он вместит. А чем больше будет нейронов, тем больше между ними установится связей и тем сложнее при прочих равных условиях может стать поведение. Правда, оговорка насчет “прочих равных” тут очень сильна (в мозге слона втрое больше нейронов, чем в мозге человека, но 97% из них находятся в мозжечке — отделе мозга, который управляет в первую очередь мышечным движением). И тем не менее, если усреднить данные по крупным группам животных, мы, скорее всего, увидим, что абсолютные цифры тут важнее относительных. В мозге муравья примерно 250 000 нервных клеток, в мозге пчелы — около миллиона. При таком количестве нейронов поведение относительно жестко программируется инстинктами (это видно хотя бы из классических книг Жана Анри Фабра, детально описывавшего сложные формы поведения перепончатокрылых). У серой крысы 200 миллионов нейронов, и это уже другое дело: все мы знаем, как разнообразно поведение этих зверьков и как хорошо развита у них способность к обучению. А у современного человека нормальное число нейронов составляет 86 миллиардов. Эусоциальное существо с мозгом такого размера просто обречено стать разумным.
Опять же — почему? Есть серьезные основания считать, что самая ресурсоемкая область применения головного мозга крупных животных — это социальные контакты, то есть выстраивание системы взаимодействий с сородичами по виду. Основанное на этом представление, связанное в основном с именем английского антрополога Робина Данбара, получило название гипотезы социального мозга. Наблюдения над разными видами обезьян показывают, что с ростом абсолютного размера мозга взаимодействия между особями очень быстро усложняются. А в сложной социальной системе, в свою очередь, индивиды с более крупным мозгом могут получить за счет этого преимущество при естественном отборе, то есть заработает положительная обратная связь. Размер мозга голого землекопа, существа размером с мышь, по всей видимости, не достиг порога, за которым эта положительная обратная связь может запуститься. Ну а размер мозга первых представителей рода Homo, появившихся около двух миллионов лет назад, — достиг.
Примем во внимание, что и крупный размер, и эусоциальность — качества в целом довольно редкие (например, из всех современных насекомых эусоциальностью обладает только 2% видов). А их сочетание, судя по всему, реализовалось за всю историю Земли только один раз. Вот результатом этого сочетания и стала наша цивилизация. Результат неизбежный, зато само сочетание крайне маловероятное.
Люди и селениты
Огромный мозг человека позволил ему реализовать в высшей степени неэкономичную эволюционную стратегию, сочетающую эусоциальность с универсальностью каждой особи. Очевидно, что любой здоровый человек потенциально способен не только размножаться, но и более-менее успешно освоить любой предлагаемый наличным социумом род деятельности (разве что с небольшими ограничениями, вытекающими из случайной индивидуальной изменчивости по таким, например, признакам, как музыкальный слух). Причем эта способность сохраняется на протяжении большей части активной жизни. Кроме того, человек способен десятилетиями накапливать знания, формируя в итоге совершенно неповторимую структуру личности с уникальными навыками. Поддержание социальной системы, состоящей из особей с такими свойствами, — дело достаточно затратное. Не могла ли природа выбрать какой-то другой путь?
Здесь помогут мысленные эксперименты, поставленные научными фантастами. Например, в романе Герберта Уэллса “Первые люди на Луне” описана гуманоидная цивилизация, устроенная совершенно по иному принципу, чем наша. Уэллсовские селениты (жители Луны) готовят каждого члена общества к строго определенной профессии с самого рождения, используя для этого не только сложную систему воспитания, но и “смелые хирургические операции”. Например, селенит-математик чисто физиологически не способен заниматься чем-либо, кроме математики: у него сильно развит мозг (и даже определенные области мозга), конечности и внутренние органы уменьшены, а сильные и яркие переживания могут быть связаны только с математикой, и ни с чем другим. Селенит-пастух, наоборот, хорошо физически тренирован, имеет глаза, защищенные “твердой и угловатой роговой оболочкой”, но не знает ничего, кроме технических понятий своего ремесла, и счастье может испытывать, только занимаясь пастушеской работой. “И так обстоит дело с селенитами всех сословий — каждый представляет собой в совершенстве законченную составную часть общей машины”.
Уэллс прекрасно знал биологию и не мог не понимать, что с точки зрения этой науки он рисует довольно реалистичную картину. Эусоциальность почти всегда приводит к тому, что отдельные особи начинают специализироваться, приобретая физиологические и даже анатомические различия между собой. Чтобы оценить последнее, достаточно сравнить, например, рабочего термита с термитом-солдатом. Уэллсовские селениты вписываются в это правило. Примерно такого уровня специализации особей и стоило бы “из общих соображений” ожидать от эусоциальных существ с крупным мозгом.
С этой точки зрения социум Homo sapiens, в котором каждая особь одновременно универсальна и уникальна, выглядит чудом. А ведь он существует уже несколько десятков тысячелетий. По эволюционным меркам это мало, но по меркам истории культуры — очень много. Более того, внутри человеческих обществ время от времени возникают очаги дополнительного “сброса специализации”. Например, после так называемой катастрофы бронзового века (XII в. до н.э.) начались события, которые Михаил Константинович Петров, известный философ, специалист по истории античной культуры, в свое время проанализировал в статье с замечательным названием: “Пентеконтера. В первом классе европейской школы мысли”. Гибель крупных империй Восточного Средиземноморья разрушила специализацию людей, основанную на кастовом делении, и породила мир, где каждый мужчина был универсальным матросом на борту столь же универсального небольшого корабля — пентеконтеры (см. рис. 17.5). По мнению Петрова, именно с этого и началось знаменитое “греческое чудо”, создавшее в конце концов европейскую цивилизацию.
Космическая рулетка
Насколько закономерны в нашей Вселенной такие явления, как жизнь и разум?
Кратко на этот вопрос не ответить. Современные научные знания не дают оснований поддерживать связанную с именем маркиза Пьера Симона де Лапласа позицию полного детерминизма. В реальности существует не только необходимое. Существование жизни не противоречит никаким положениям физики или химии, но оно и не вытекает непосредственно из этих положений. Обратим внимание, что такое утверждение верно далеко не для всех природных явлений: например, образование атомов, молекул, звезд при данных физических законах было неизбежно. О возникновении жизни этого сказать нельзя. Звездные системы не обязательно порождают жизнь, как и жизнь (судя по всему) не обязательно порождает разум. Во всяком случае, у нас пока нет никаких серьезных оснований утверждать обратное.
Можно мысленно прочертить траекторию, соединяющую в одно целое все ключевые моменты химической и биологической эволюции, начиная от формирования планетной системы и заканчивая становлением цивилизации. Такие события, как возникновение первых клеток, многоклеточности или мозга, будут на этой траектории промежуточными точками. Как же оценить вероятность того, что жизнь на данной планете пройдет по ней от начала до конца?
Думается, что тут уместна вот какая аналогия. Представим себе игрока в рулетку, который все время ставит на красное с одним дополнительным условием: серия выпадений красного должна быть обязательно непрерывной. Пока раз за разом выпадает красное, человек остается в игре. Если хотя бы один раз выпало черное, игра прекращается и он выбывает. Какие шансы будут у такого игрока?
Это легко подсчитать. При одном испытании игрой в рулетку вероятность выпадения красного составляет ½. Но уже при десятке испытаний вероятность постоянного выпадения красного станет меньше 1/1000, а при сотне испытаний в знаменателе окажется вполне астрономическое 32-значное число. Вот оно-то и будет характеризовать соотношение тех, кто выиграл, и тех, кто проиграл.
Похоже, что жизнь в космосе сталкивается примерно с такой же игрой вероятностей. Для каждого отдельного фактора или события вероятность помешать развитию жизни может быть невысока. Настоящая проблема в том, что этих факторов и событий очень много. Планета не должна оказаться слишком близко или слишком далеко от звезды, не должна быть разрушена ударом другого небесного тела, не должна проявлять слишком высокую или слишком низкую тектоническую активность — продолжать в таком роде можно до бесконечности. Между тем никакого суммирования, приводящего к взаимной компенсации, здесь нет. Единичного события, хотя бы на короткое время выводящего условия на планете за пределы пригодных для жизни, может быть достаточно, чтобы закрыть вопрос навсегда, даже если значения всех остальных переменных при этом остаются в пределах допустимого.
Более того, биосфера может не только погибнуть от внешних по отношению к ней факторов (это можно назвать угрозой первого рода), но и буквально совершить самоубийство (угроза второго рода). Причем для этого не обязательно нужен разумный агент, который устроит термоядерную войну. У известного геолога Джозефа Киршвинка есть интересная статья под названием “Красная Земля, белая Земля, зеленая Земля, черная Земля”. Под “красной Землей” тут понимается Земля в начале кислородной эпохи: тогдашние осадки богаты окисленными соединениями железа, часто имеющими красноватый оттенок. “Белая Земля” — это Земля эпохи “Земли-снежка”. “Зеленая Земля” — это современная Земля с ее почти сплошным растительным покровом. А вот “черная Земля” — это ситуация, которая, по мнению Киршвинка, в принципе могла бы реализоваться в далеком будущем. Современные системы фотосинтеза поглощают свет разных цветов (синий, красный), но вот зеленый свет они большей частью отражают. Именно поэтому современные растения и выглядят зелеными. Между тем в солнечном свете зеленый компонент очень силен. А что, если эволюция создаст фотосинтезирующий организм с такими пигментами, которые будут преимущественно поглощать зеленый свет? Это вполне может повысить эффективность фотосинтеза и дать такому организму преимущество в выживании: эволюция — слепой скульптор, она никогда не заглядывает вперед дальше чем на один шаг. Земля, покрытая растениями, поглощающими зеленый свет, будет из космоса выглядеть черной. Отражательная способность планеты при этом грозит снизиться настолько, что биосфера не сможет справиться с перегревом и большинство живых организмов погибнет от высокой температуры (как это чуть было не произошло на рубеже перми и триаса). При всей внешней фантастичности этого сценария никаким законам природы он не противоречит.
Говоря о жизни в космосе, мы сразу сталкиваемся с еще одной проблемой, связанной скорее с человеческим восприятием реальности. Рассуждая о том, почему мы до сих пор не встретили инопланетные цивилизации, люди очень часто основываются (больше подсознательно, чем сознательно) на старинных представлениях, согласно которым Вселенная практически вечна. Философы, убежденные, что мир никогда не рождался и никогда не умрет, были даже в Средневековье. В Новое время близкую к этому позицию выразил, например, шотландский геолог Джеймс Геттон, писавший: “В истории Земли мы не видим никаких следов начала и никаких признаков конца”. Но сейчас-то мы знаем, что это совсем не так! Любая планетная система имеет конечный срок существования, в который развивающаяся там жизнь должна уложиться. И в сравнении с темпами эволюции самой жизни этот срок не так уж велик. Например, часть “жизненного цикла” Солнца, охватывающая промежуток от протозвезды до красного гиганта, должна занять около 10 миллиардов лет. Это всего-навсего вдвое больше, чем уже длится история Солнечной системы.
Кроме того, возникновение жизни было невозможно в первые миллиарды лет после Большого взрыва, пока звезды первых поколений не достигли стадии сверхновых и не взорвались, разбросав по Галактике пригодные для сборки планетных систем и живых тел тяжелые химические элементы. Считается, что наше Солнце — звезда даже не второго, а третьего поколения. В древней Вселенной, включавшей только звезды первого поколения и состоявшей почти исключительно из водорода и гелия, никакой жизни быть не могло. Это дополнительно ограничивает максимальный срок, отпущенный на биологическую эволюцию.
С другой стороны, эта эволюция обычно идет очень неравномерно. Например, первые эукариоты появились только после “кислородной революции”, и, скорее всего, вследствие нее. До этого биосфера Земли в течение двух миллиардов лет была чисто прокариотной. Но и в эукариотной биосфере эволюция отнюдь не всегда шла быстро: за “эукариотным Большим взрывом” (если он был) последовал “скучный миллиард лет”, в течение которого в живой природе Земли не появилось никаких качественных новшеств. Вполне возможно, что без них обходилось бы и дальше, если бы не катастрофическое оледенение, вызванное особенностями дрейфа континентов и приведшее к вынужденной перестройке всей биосферы (эпоха “Земли-снежка”). Сложись геологическая история Земли несколько иначе, самыми сложными живыми организмами на ней до сих пор могли бы быть строматолиты — или, в лучшем случае, красные водоросли. Более того, не исключен сценарий, когда целая биосфера доживает до гибели в огне своего солнца (ставшего красным гигантом), так и не успев породить ни многоклеточных животных, ни высших растений.
Отсюда следует ясный вывод. Обычно (и чаще всего неявно) принимаемое “господами ксенологами” допущение, что время, нужное для развития цивилизации, пренебрежимо мало по сравнению с временем существования Вселенной, совершенно неверно. На самом деле эти времена сравнимы друг с другом. Первое, конечно, меньше, но не на порядки, а всего лишь в разы. На Земле от возникновения жизни до возникновения разума прошло время, приблизительно равное трети времени существования Вселенной, и ничего не стоит вообразить сценарий, где это время было бы еще больше.
Эта логика сразу подсказывает возможное решение пресловутой загадки “молчания Вселенной” (Silentium Universi), или основного парадокса ксенологии, как предпочитали выражаться Стругацкие. Весьма вероятно, что наша цивилизация — просто первая в Галактике. Другие биосферы, если они где-то и существуют, еще не успели дойти до этой стадии. Только и всего. Может быть, успеют в будущем — а может быть, и нет.
“Человек — не статический центр мира, как он долго полагал, а ось и вершина эволюции, что много прекраснее”, — писал в середине XX века Пьер Тейяр де Шарден. Что ж, современные научные данные не исключают, что Тейяр был в каком-то смысле прав. Удивительный сверхорганизм человечества — острие эволюции жизни во Вселенной.
Назад: 16. криптозой
Дальше: “Философский камень” биологии

