Книга: От атомов к древу. Введение в современную науку о жизни
Назад: ЧАСТЬ IV ИСТОРИЯ ЖИЗНИ
Дальше: 17. фанерозой
16. криптозой
И вновь, прорезав плотные туманы,На теплые архейские моря,Где отбивают тяжкий пульс вулканы,Льет бледный свет пустынная заря.
И, размножая легких инфузорий,Выращивая изумрудный сад,Все радостней и золотистей зориИз облачного пурпура сквозят.Михаил Зенкевич. Темное родство
Зададимся следующим вопросом: можно ли рассматривать историю Вселенной как единый процесс, начавшийся Большим взрывом, охватывающий наше время и продолжающийся в неведомом космическом будущем? Средневекового книжника такой подход, вероятно, совсем не удивил бы. Написанная в XII веке хроника Оттона Фрейзингенского — “самая всеобщая из всех всеобщих историй”, по выражению великого историка-медиевиста Марка Блока, — начинается с сотворения мира и заканчивается Страшным судом. В конце XX века такое отношение к вещам возродилось в учебной дисциплине, которая получила название “большой истории” (Big History). Курсы “большой истории” выстраивают в единую последовательность события любого рода — от зарождения первых звезд до промышленной революции. Все развитие мира там представлено единой шкалой, охватывающей 13,8 миллиарда лет: примерно столько времени, по расчетам, прошло от Большого взрыва до современности.
Ясно, что “большая история” ставит себе грандиозную задачу. Требуемый для нее объем сведений (еще и стремительно растущий) очень трудно уложить в сколько-нибудь компактный рассказ. В этом отношении Оттону Фрейзингенскому приходилось куда легче, чем авторам, работающим с современными научными данными. Поэтому важнейшим понятием в “большой истории” является порог (threshold) — момент качественного изменения системы, дающий историку точку отсчета. В любом историческом процессе число по-настоящему значительных порогов относительно невелико. А привязав к ним обзор, можно “с птичьего полета” увидеть важные вещи, которые иначе могли бы и ускользнуть от внимания.
Попробуем, не вдаваясь ни в какие вселенские обобщения, применить этот подход к биологическому и палеонтологическому материалу. Мы сразу увидим, что за несколько миллиардов лет земная жизнь не раз миновала качественные пороги, любой из которых можно было бы и не пройти. Причем иногда это выглядело весьма драматично, напоминая пропасть, через которую перекинут узкий мост. И после каждого такого порога не только жизнь на Земле, но и вся Земля как планета серьезно менялась. Пущенное в оборот австрийским геологом Эдуардом Зюссом выражение “лик Земли” как нельзя лучше характеризует объект этих изменений.
Для удобства разговора введем два термина: биосфера — целостная живая оболочка Земли и биота — совокупность видов живых организмов. Биота отличается от биосферы тем, что связи между организмами в нее не включены: грубо говоря, это просто список видов. Термин “биосфера” предложил сам Эдуард Зюсс, а термин “биота” — румынский зоолог Эмиль Раковицэ. Каждый пройденный глобальный порог навсегда менял состояние и биосферы, и биоты.
Ясно, что любая “пороговая” схема эволюции биосферы будет в чем-то несовершенна просто из-за того, что частью событий совершенно неизбежно придется пренебречь. Тем не менее попытки создания подобных схем иногда удаются. Замечательным образцом этого подхода можно назвать книгу Кирилла Еськова “История Земли и жизни на ней”, основанную на учебном курсе и выдержавшую несколько переизданий. К сожалению, со времени создания этой книги прошло уже около 20 лет, а для такой бурно развивающейся науки, как палеонтология, это огромный срок. Нисколько не намереваясь критиковать великолепную работу Кирилла Юрьевича, попробуем предложить что-то вроде краткого комментария к ней.
Здесь мы рассмотрим семь порогов, соответствующих важным событиям в истории Земли: (1) появление жизни как таковой, (2) кислородная революция, (3) эпоха “Земли-снежка”, (4) кембрийский взрыв, (5) появление лесов и насекомых, (6) пермский кризис и (7) приобретение некоторыми животными эусоциальности. Поговорим о них по порядку (см. рис. 16.1).
Логика случая
Насколько закономерным или случайным событием по меркам Вселенной было появление земной жизни? Тут будет интересно сравнить мнения двух крупных биологов, работавших примерно в одно и то же время. Это француз Жак Моно и бельгиец Кристиан де Дюв. Оба они получили Нобелевскую премию за открытия в области клеточной биологии: Моно открыл важные механизмы регуляции работы генов, а де Дюв обнаружил несколько новых клеточных органелл. В том, как работает живая клетка, и Моно, и де Дюв разбирались превосходно. К тому же они поддерживали дружеские отношения, благо оба принадлежали к франкоязычному миру. Но вот их взгляды на происхождение жизни оказались совершенно разными.
Жак Моно считал, что раз жизнь (насколько мы пока знаем) уникальна, то нет никакой необходимости объяснять ее появление чем-то иным, кроме игры слепого случая. В конце концов, Вселенная настолько огромна, что где-нибудь на ее просторах вполне может один раз произойти любое сколь угодно маловероятное событие. Достаточно, чтобы вероятность такого события не была строго нулевой. Это не дает повода принимать случайность за закономерность.
“Вселенная не была чревата жизнью, как и биосфера не была чревата человеком, — писал Моно. — Нам просто выпал счастливый шанс в рулетке, как тому, кто только что выиграл миллион в казино”.
Де Дюв, наоборот, считал, что появление жизни — это закономерный процесс, готовый реализоваться на любой планете с подходящими физико-химическими условиями. Возражая Моно, он говорил, что Вселенная как раз “чревата жизнью”, и приводил два довода, которые с тех пор стали только сильнее.
Во-первых, химические “строительные блоки”, пригодные для создания живых систем, легко синтезируются в космосе. Хорошо известно, что они входят в состав метеоритов и комет. Де Дюв пользуется здесь труднопереводимым выражением “vital dust” — “жизнетворная пыль”. По его словам, “жизнетворная пыль” пронизывает всю Вселенную и образует своего рода семена жизни, готовые взойти на любом подходящем небесном теле. И в самом деле, сейчас мы точно знаем, что в метеоритах есть аминокислоты, сахара, азотистые основания, спирты, жирные кислоты и другие углеродные соединения, причем все они там довольно разнообразны. Похоже, что их синтез не требует никаких особенно редких условий.
Во-вторых, возникновение жизни — процесс по своей сути химический. Все информационные аспекты здесь вторичны. Истинные действующие лица — это нуклеиновые кислоты, углеводы, липиды и другие молекулы. А уж химию-то мы знаем хорошо — и можем уверенно сказать, что все химические процессы в большой степени детерминистичны, то есть дают при одних и тех же условиях один и тот же результат. Конечно, здесь участвует статистика (поскольку молекул очень много), но в итоге на волю случая почти ничего не остается. Применительно к нашей задаче это должно означать, что если на какой-нибудь планете сложатся такие же условия, какие были на Земле примерно четыре миллиарда лет назад, то вероятность возникновения жизни там будет близка к единице.
Сейчас мы понимаем, что многие свойства живых объектов на самом деле предопределены чистейшей химией. Например, каталитическая активность РНК, то есть ее способность ускорять химические реакции, появляется автоматически, как только начинают синтезироваться (каким угодно способом) цепочки РНК длиной хотя бы в десятки нуклеотидов. Сборка жирных кислот и других липидов в мембраны, подобные клеточным, тоже происходит сама собой, стоит только им попасть в водный раствор. Для всего этого достаточно подходящих внешних условий. Другое дело, что такие условия далеко не повсеместны, и — что еще важнее — даже там, где они готовы сложиться, системе ничего не стоит выйти за их пределы по каким-нибудь случайным причинам; пользуясь английской идиомой, это проще, чем упасть с бревна (as easy as falling off a log). Например, на современной Венере зарождение жизни земного типа совершенно невозможно, хотя почти по всем физическим параметрам эта планета очень близка к Земле и изначально они, скорее всего, были гораздо более похожи друг на друга, чем сейчас. Судьба Венеры наглядно показывает, насколько Земле повезло.
Первая жизнь
Когда на Земле появилась жизнь? Самый распространенный до недавнего времени ответ на этот вопрос таков: древнейшие предполагаемые остатки живых организмов найдены в Гренландии, в горных породах зеленокаменной формации Исуа, имеющих возраст 3,8 миллиарда лет. Значит, к этому времени жизнь уже точно существовала. Правда, неизвестно, какая именно. И вот тут таится первая проблема. Дело в том, что найденные в Исуа остатки не сохранили никаких следов структуры живых клеток. Это зерна чистого углерода, и вывод о том, что они когда-то были живыми существами, сделан исключительно по составу этого углерода.
Чтобы понять, в чем тут дело, вспомним, как устроены атомы (см. главу 1). Главный параметр любого атома — число протонов, или атомный номер (Z). Только от него зависит, к какому химическому элементу атом относится. Однако в атомном ядре есть не только протоны, но и нейтроны. Суммарное количество протонов и нейтронов в ядре данного атома называется его массовым числом (A). И вот оно у атомов одного и того же элемента может различаться. Например, любой атом, в ядре которого шесть протонов, будет атомом углерода. Но есть несколько разных типов атомов углерода, например с шестью нейтронами в ядре (12C) или с семью нейтронами в ядре (13C). Атомы, имеющие одинаковый атомный номер, но разное массовое число, называются изотопами.
Углекислый газ (CO2) может включать в себя как атом 12C, так и атом 13C. Но вот белок, связывающий углекислый газ для фотосинтеза, гораздо охотнее захватывает молекулы CO2 с углеродом 12C просто потому, что они более легкие. В результате происходит разделение изотопов. На самом деле оно идет не только при фотосинтезе, но и при других способах биологической фиксации CO2, но фотосинтез — самый распространенный из них на современной Земле. Однако в любом случае отсюда следует, что живые организмы, прямо или опосредованно питающиеся продуктами фиксации углекислоты — то есть практически все живые организмы на свете, — имеют смещенное соотношение “легкого” и “тяжелого” изотопов углерода: доля “тяжелого” изотопа там заметно понижена по сравнению с атмосферным углекислым газом. А это означает, что, найдя чистый углерод, можно по соотношению 12C/13C определить, является ли этот углерод биогенным, то есть входил ли он когда-нибудь в состав живого тела.
Но что, если при переплавлении горных пород заработал какой-нибудь другой, чисто физический механизм разделения изотопов углерода? Это теоретически возможно, и некоторые ученые считают, что в случае с породами Исуа именно так оно и было. Тогда “следы самой древней жизни” исчезают. Нельзя сказать, что эта тема закрыта, но статус пород Исуа сейчас определенно под сомнением. Печальнее всего, что биологи тут и сделать ничего не могут — решающее слово принадлежит геологии и изотопной химии. Биогенное происхождение углерода из Исуа не исключено, оно просто спорно. С другой же стороны, в самое последнее время появились данные, что в формации Исуа есть остатки строматолитов — слоистых бактериальных колоний, но это еще требует проверки.
В любом случае зеленокаменная формация Исуа, судя по всему, не предел. Недавно было объявлено, что найден предположительно биогенный углерод возрастом 4,1 миллиарда лет . Это совершенно поразительно, потому что для настолько древних времен почти неизвестны полноценные горные породы. Главные сохранившиеся с тех пор твердые объекты — это мелкие зерна чрезвычайно устойчивого минерала циркона, захороненные где-нибудь в более поздних осадках. Вот в составе этих цирконовых зерен геологи и нашли углерод со смещенным изотопным соотношением, типичным для живых организмов. По оценке авторов исследования, другие пути разделения изотопов в данном случае маловероятны, так что это могут быть следы жизни — невообразимо древней жизни! Какой она была, остается загадкой, ведь в изученных образцах налицо только химический сигнал.
Между тем первые живые организмы могли очень сильно отличаться от современных — причем под “современностью” в данном случае приходится подразумевать не более и не менее как последние три с лишним миллиарда лет. Например, мы знаем, что в современной цепочке передачи генетической информации есть три главных звена: копирование ДНК (репликация), синтез РНК (транскрипция) и синтез белка (трансляция). При этом молекулярные данные свидетельствуют, что у общего предка всех клеточных организмов системы транскрипции и трансляции были намного более простыми, чем у современных клеток, а системы репликации ДНК не было совсем. Еще Карл Вёзе показал, что белки репликации бактерий не имеют ничего общего с белками репликации архей и эукариот. Скорее всего, это означает, что весь механизм копирования ДНК возник минимум дважды — у бактерий и у архей (эукариоты унаследовали его от последних). Тогда получается, что у общего предка всех их, вместе взятых, генетическая информация хранилась в основном на РНК, как и предполагает популярная в наше время теория “РНК-мира”.
Кроме того, этот общий предок вполне мог еще не достичь так называемого дарвиновского порога — момента, когда интенсивность привычной нам вертикальной передачи генов (от предков к потомкам) начала существенно превышать интенсивность горизонтального переноса генов (между соседними геномами независимо от родства). Понятие “дарвиновский порог” (Darwinian Threshold) ввел тот же Карл Вёзе — он вообще много занимался ранними этапами эволюции. Нам сейчас трудно вообразить, как выглядела жизнь по ту сторону дарвиновского порога, но ясно, что тогдашние организмы были предельно изменчивы: никаких устойчивых видов в тех условиях существовать не могло. Нетрудно догадаться, что устойчивость биологических видов определяется именно надежной передачей генетической информации от предков к потомкам. Когда этот механизм еще не сложился, мир был совершенно другим. Вёзе потому и назвал порог дарвиновским, что его переход означал происхождение видов в самом что ни на есть буквальном смысле слова “вид” (а “Происхождением видов”, как известно, называется главная книга Дарвина). Живые системы, не достигшие дарвиновского порога, просто не могли распадаться на биологические виды таким же образом, как распадается на них жизнь сейчас.
Самые древние более-менее достоверно определимые остатки живых клеток имеют возраст 3,4 миллиарда лет. Это уже типичные прокариоты, скорее всего входящие в дожившую до современности группу сульфатредуцирующих бактерий. На этой отметке заканчивается туманная повесть о происхождении жизни и начинается ее собственная история.
Обоюдоострый меч О2
Переходя ко второму порогу — кислородной революции, надо прежде всего сказать, что биологические свойства молекулярного кислорода (O2) как минимум двуедины. Кислород — мощный окислитель, с помощью которого можно получить много полезной энергии, и в то же время сильный яд, свободно проходящий сквозь клеточные мембраны и разрушающий структуру клеток. Иногда говорят, что кислород — это обоюдоострый меч. Древнейшим жителям Земли существа, дышащие кислородом, скорее всего, показались бы кем-то вроде людей с фторной планеты из повести “Сердце Змеи” (см. главу 2). У всех организмов, имеющих дело с кислородом, обязательно есть специальные ферментные системы, гасящие его агрессивное химическое воздействие. А те, у кого таких ферментных систем нет, обречены быть строгими анаэробами, выживающими только в бескислородной среде. На современной Земле это некоторые бактерии и археи.
Практически весь молекулярный кислород на Земле имеет биогенное происхождение, то есть выделяется живыми существами. Главный источник O2 — это кислородный фотосинтез. Других процессов, способных давать его в сравнимых количествах, просто нет. Как мы уже знаем, фотосинтезом называется синтез глюкозы (С6H12O6) из углекислоты (CO2) и воды (H2O), происходящий с помощью энергии света. Главным “действующим лицом” тут служит углекислый газ, который восстанавливается водой. Кислород же в этой реакции — не что иное, как побочный продукт, отход. Фотосинтез может вообще не приводить к выделению кислорода, если вместо воды в нем используется в качестве восстановителя какое-нибудь другое вещество — например, сероводород, свободный водород или соединения железа. Такой фотосинтез называется бескислородным, и есть несколько его разных вариантов.
Бескислородный фотосинтез наверняка появился раньше кислородного. Поэтому в первый миллиард лет существования жизни (а скорее всего, дольше) фотосинтез хотя и шел, но никакого насыщения атмосферы Земли кислородом не вызывал. Содержание кислорода в атмосфере в те времена составляло не больше 0,001% от современного — попросту говоря, это значит, что его там не было.
Все изменилось, когда на сцену вышли синезеленые водоросли, или цианобактерии. Впоследствии именно эти существа стали предками хлоропластов, фотосинтезирующих элементов клеток эукариот. Цианобактерии — очень древняя эволюционная ветвь. По меркам земной истории, они удивительно неизменны. Например, широко распространенная в современных водоемах синезеленая водоросль осциллятория имеет ископаемых родственников, живших 800 миллионов лет назад, причем от современных осцилляторий они практически неотличимы. Таким образом, осциллятория — чрезвычайно впечатляющий пример живого ископаемого. Но самые первые цианобактерии появились намного раньше нее, это надежно подтверждено палеонтологическими данными.
Поначалу цианобактерии не были многочисленны, потому что освоенный ими кислородный фотосинтез не давал никаких серьезных преимуществ по сравнению с бескислородным, которым владели другие группы микробов. Но химическое окружение этих микробов постепенно менялось. Наступил момент, когда “сырья” для бескислородного фотосинтеза просто перестало хватать. И вот тогда час цианобактерий пробил. Кислородный фотосинтез имеет одно большое преимущество и один большой недостаток. Преимущество — это совершенно неограниченный запас исходного реагента-восстановителя (воды), а недостаток — высокая токсичность побочного продукта (кислорода). Неудивительно, что поначалу этот тип обмена не был “популярен”. Зато при малейшем дефиците других субстратов, кроме воды, обладатели кислородного фотосинтеза должны были сразу получить конкурентное преимущество, что, по всей видимости, и произошло. После этого наступила эпоха длиной примерно в миллиард лет, на протяжении которой облик Земли определяли в первую очередь цианобактерии. Недавно эту эпоху даже предложили неофициально назвать в их честь “цианозоем”.
Именно из-за цианобактерий 2,4 миллиарда лет назад началась кислородная революция, она же кислородная катастрофа, или Великое окислительное событие (Great Oxidation Event, GOE). Строго говоря, это событие не было ни мгновенным, ни абсолютно уникальным. Краткие всплески концентрации кислорода, так называемые “кислородные дуновения”, случались и раньше — это палеонтологически зафиксировано. И все же 2,4 миллиарда лет назад произошло нечто новое. За короткое по меркам земной истории время (считаные десятки миллионов лет) концентрация кислорода в атмосфере выросла примерно в 1000 раз и осталась на этом уровне (см. рис. 16.2). До прежних ничтожных величин она не опустилась больше никогда. Биосфера необратимо стала кислородной.
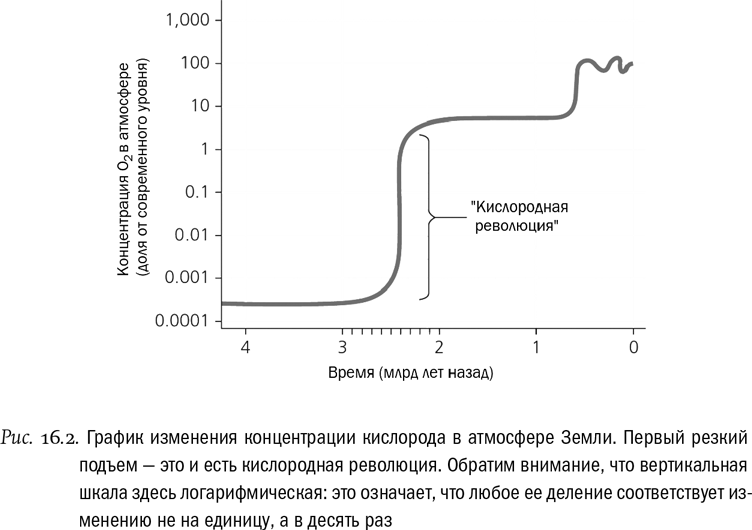
Для подавляющего большинства древних прокариот такой уровень кислорода был смертельно опасен. Неудивительно, что первым результатом кислородной революции, скорее всего, стало вымирание (хотя, возможно, и не столь катастрофическое, как думали ученые лет 30 назад, когда появился термин “кислородный холокост”). Выжили в первую очередь те, кто успел создать защищающие от кислорода ферментные системы. Есть основания полагать, что в первые 100–200 миллионов лет “нового кислородного мира” кислород был для живых организмов только ядом, и ничем больше. А вот потом ситуация поменялась. Ответом биоты на кислородный вызов стало появление бактерий, которые включили кислород в цепочку реакций, разлагающих глюкозу, и таким образом начали использовать его для получения энергии.
Сразу оказалось, что кислородное окисление глюкозы (дыхание) намного энергетически эффективнее бескислородного (брожения). В расчете на одну молекулу глюкозы оно дает в несколько раз больше свободной энергии, чем любой сколь угодно “навороченный” вариант бескислородного обмена. При этом все начальные этапы распада глюкозы у пользователей дыхания и брожения остались общими: кислородное окисление послужило всего лишь надстройкой над уже имевшимся древним биохимическим механизмом, который сам по себе в кислороде не нуждался.
Группа микробов, которая освоила рискованное, но эффективное получение энергии с помощью кислорода, называется протеобактериями. Согласно общепринятой сейчас теории, именно от одной из их подгрупп произошли дыхательные органеллы эукариотных клеток — митохондрии. Судя по генетическим данным, ближайший современный родственник митохондрий — это пурпурная спиральная альфа-протеобактерия Rhodospirillum rubrum. Родоспириллум обладает и дыханием, и брожением, и бескислородным фотосинтезом (в котором вместо воды используется сероводород), и может переключаться между этими тремя типами метаболизма в зависимости от внешних условий. Без сомнения, такой симбионт был предку эукариот очень полезен.
Более того, многие современные ученые считают, что симбиоз древних архей с альфа-протеобактериями был толчком к образованию эукариотной клетки. Это “раннемитохондриальная” гипотеза, которую мы уже обсуждали в главе 10. Как мы помним, она предполагает, что разделение будущей эукариотной клетки на цитоплазму и ядро произошло только после того, как в нее внедрился симбионт — протеобактерия. Более старый “позднемитохондриальный” сценарий, согласно которому протеобактерия была попросту проглочена готовой эукариотной клеткой, сейчас выглядит несколько менее вероятным. Но надо учитывать, что в любом случае обе клетки — и архейная, и протеобактериальная — были в процессе объединения серьезно “пересобраны”, породив своего рода химеру с новыми свойствами. Эта химера и стала эукариотной клеткой. Молекулярные компоненты архейного и протеобактериального происхождения в ней сильно перемешались, разделив между собой функции. Так или иначе без протеобактерий эукариоты не возникли бы. А это означает, что их появление было прямым следствием кислородной революции. В свете сказанного почти не выглядят преувеличением слова двух современных крупных ученых, палеонтолога и геолога: “Все согласны с тем, что эволюция синезеленых водорослей была самым значительным биологическим событием на нашей планете (даже более значительным, чем развитие эукариотических клеток и появление многоклеточных организмов)”. Действительно, знакомый нам мир животных и растений не существовал бы, если бы не цианобактерии и вызванный ими кризис. При всей малопривлекательности марксизма приходится признать, что известный тезис Маркса “революции — локомотивы истории” применительно к биосферным революциям иногда все-таки подтверждается.
Эпохи жизни
Вся история Земли делится на четыре огромных промежутка, именуемых эонами. Названия эонов следующие: катархей или гадей (4,6–4,0 млрд лет назад), архей (4,0–2,5 млрд лет назад), протерозой (2,5–0,54 млрд лет назад) и фанерозой (начался 0,54 млрд лет назад и продолжается сейчас). Первые три эона (то есть все, кроме фанерозоя) принято объединять понятием “криптозой”, что буквально значит “скрытая жизнь”. Это деление будет нам постоянно помогать. Сделаем оговорку, что почти во всех подобных случаях запоминать стоит не точные датировки, а саму последовательность эпох и относящихся к ним событий: она гораздо важнее (см. рис. 16.1). Исключение можно сделать разве что для двух-трех ключевых дат вроде возраста Земли.
Катархей — это так называемая догеологическая эпоха, от которой почти не осталось “нормальных” горных пород, расположенных послойно. Классические геологические и палеонтологические методы, основанные на сравнении последовательных слоев отложений, там принципиально неприменимы. Оставшиеся от катархея твердые объекты — это в первую очередь мелкие зерна циркона, те самые, в которых недавно нашли предположительно биогенный углерод. О катархейской жизни (если она была) известно крайне мало.
В архее Земля принадлежит прокариотам — бактериям и археям (совпадение корней в названии геологической эры “архей” и группы микробов “археи” случайно). Граница архея и протерозоя приходится примерно на момент одного из сильных “кислородных дуновений”, предшествовавших кислородной революции. Сама кислородная революция произошла уже внутри протерозоя, хотя и в самом его начале.
Протерозой — это эпоха кислорода и эукариот. О кислороде мы уже говорили, а с датировкой происхождения эукариот связан интересный парадокс. Дело в том, что более-менее надежно определимые многоклеточные эукариоты появляются в палеонтологической летописи гораздо раньше, чем столь же надежно определимые одноклеточные. Нитчатая водоросль Grypania spiralis, которую обычно считают эукариотом, появилась 2,1 миллиарда лет назад. Справедливости ради нужно сказать, что главным доводом за эукариотную природу грипании служит ее крупный размер — все остальные признаки на самом деле не дают уверенности, что это не гигантская колония цианобактерий. Но дело в том, что эта находка далеко не единственная. Самым древним известным эукариотом сейчас считается грибообразный организм Diskagma buttonii возрастом 2,2 миллиарда лет. А еще есть загадочные крупные спиралевидные существа — скорее всего, водоросли, возраст остатков которых не меньше 2,1 миллиарда лет, как и у грипании. Зато самые ранние одноклеточные, однозначно определяемые как эукариоты, имеют возраст всего 1,6 миллиарда лет. Это, разумеется, не значит, что многоклеточные эукариоты возникли раньше одноклеточных — такое предположение противоречит и молекулярным данным, и здравому смыслу. Одноклеточные просто хуже сохраняются, да и признаков, по которым можно понять, что это за организм, у них меньше.
Из приведенных цифр следуют важные общебиологические выводы. Вспомним, что дата кислородной революции — 2,4 миллиарда лет назад. Следовательно, мы теперь знаем, что всего через 200 миллионов лет после кислородной революции в палеонтологической летописи появляются не просто эукариоты, а многоклеточные эукариоты. Это означает, что первые этапы эволюции эукариот были пройдены, по меркам глобальной истории, очень быстро. Безусловно, эукариотной клетке потребовалось некоторое время, чтобы оформить симбиоз с предками митохондрий, создать ядро, усложнить цитоскелет. Но после того, как эти процессы завершились, первые многоклеточные организмы возникли буквально сразу же. На уровне клетки это не потребовало никаких дополнительных приспособлений. Любая эукариотная клетка уже имеет в наличии полный набор молекулярных структур, необходимых, чтобы построить из таких клеток многоклеточное тело (во всяком случае, относительно простое). Разумеется, все эти структуры не менее полезны и для жизни одиночной клетки, иначе они просто не появились бы. Общий предок эукариот, без сомнений, был одноклеточным, и очень многим его потомкам многоклеточность так никогда и не пригодилась. Насколько разнообразны современные одноклеточные эукариоты, мы отчасти знаем из главы 15.
Добавим, что совсем недавно палеонтологи описали нитчатые структуры, очень похожие по деталям строения на настоящую грибницу, в горных породах, возраст которых оценивается в 2,2–2,4 миллиарда лет (причем 2,2 миллиарда лет — это минимальная оценка). Кстати, имеющая возраст 2,2 миллиарда лет Discagma во многом напоминает относительно примитивные современные грибы — зигомицеты (см. рис. 16.3). Если предположение о таком огромном возрасте грибов подтвердится, это заодно послужит прекрасным подтверждением гипотезы “эукариотного Большого взрыва”, которую мы обсуждали в главе 15. Тогда волей-неволей получится, что все главные эволюционные ветви эукариот возникли очень быстро, за считаные десятки миллионов лет, — типичная взрывная эволюция. Альтернативное объяснение могло бы состоять в том, что эукариоты появились на несколько сотен миллионов лет раньше, чем мы сейчас думаем. Такая гипотеза тоже существует, но она пока более чем спорная.
Кислородная революция имела еще одно важное последствие, коснувшееся состава атмосферы. В архейской атмосфере было много азота (как и сейчас), а также углекислого газа и метана (гораздо больше, чем сейчас). Углекислый газ и метан очень хорошо поглощают инфракрасное излучение и тем самым удерживают в атмосфере Земли тепло, мешая ему уходить в космос. Это называется парниковым эффектом. Причем считается, что от метана парниковый эффект как минимум раз в 20–30 сильнее, чем от углекислого газа. А в архейские времена метана в атмосфере Земли было примерно в 1000 раз больше, чем сейчас, и это обеспечивало довольно теплый климат.
Тут вмешивается еще и астрономия. Согласно общепринятой теории эволюции звезд, светимость Солнца медленно, но непрерывно растет. В архее она составляла всего 70–80% от современной — понятно, почему парниковый эффект был важен для поддержания планеты в тепле. Но после кислородной революции атмосфера стала окислительной и почти весь метан (CH4) превратился в углекислый газ (CO2), эффективность которого как парникового газа гораздо ниже. Это вызвало катастрофическое гуронское оледенение, длившееся около 100 миллионов лет и в некоторые моменты охватывавшее практически всю Землю: на участках суши, которые тогда находились всего в нескольких градусах широты от экватора, найдены следы ледников. Пик гуронского оледенения наступил 2,3 миллиарда лет назад. К счастью, оледенение не могло остановить тектоническую активность земной мантии. Вулканы продолжали выбрасывать в атмосферу углекислый газ, и со временем его накопилось достаточно, чтобы восстановить парниковый эффект и растопить льды.
Надо сразу сказать, что такое климатическое испытание было в истории Земли не последним. Самое широко известное глобальное оледенение — позднепротерозойское, о котором речь впереди. Оно оставило больше следов, чем раннепротерозойское гуронское оледенение, и было раньше открыто палеонтологами, став в свое время настоящей сенсацией. Видимо, при определенных обстоятельствах Земле было не так уж и трудно превратиться в ледяной шар.
“Скучный миллиард лет” и его конец
За бурными событиями начала протерозоя последовал так называемый “скучный миллиард лет” (Boring Billion). В это время не происходило никаких оледенений, никаких резких перемен в составе атмосферы, никаких биосферных переворотов. Эукариотные водоросли жили в океанах, понемногу выделяя кислород. Их мир был по-своему разнообразным и сложным. Например, из эпохи “скучного миллиарда” известны многоклеточные красные и желто-зеленые водоросли, поразительно похожие на своих современных родственников. Известны оттуда и грибы. А вот многоклеточные животные на просторах “скучного миллиарда лет” отсутствуют. Будем аккуратны: на нынешний момент никто не может с полной уверенностью утверждать, что многоклеточных животных тогда не было, но аргументы в пользу того, что они были, в лучшем случае очень сомнительны.
В чем тут дело? Напрашивается мысль, что многоклеточность как таковая гораздо более совместима с образом жизни растения, чем с образом жизни животного. Любая клетка растения заключена в жесткую клеточную стенку, и нет сомнений, что это сильно облегчает регуляцию взаимного расположения клеток в сложном теле. Наоборот, клетки животных вообще лишены клеточной стенки, их форма неустойчива да еще и постоянно меняется при актах движения или фагоцитоза. Собрать из таких клеток целый организм — сложная задача. Если бы никаких многоклеточных животных не появилось вовсе, а биологами стали представители растений либо грибов, они, скорее всего, после изучения этой проблемы пришли бы к выводу, что сочетание многоклеточности с отсутствием клеточной стенки просто невозможно. Во всяком случае, это объясняет, почему многоклеточность много раз возникала в разных группах водорослей, но только один раз — у животных.
Есть и другая идея. В 1959 году канадский зоолог Джон Ральф Нерселл связал внезапное (как тогда считалось) появление животных в палеонтологической летописи с ростом концентрации кислорода в атмосфере. Животные, как правило, обладают активной подвижностью, которая требует столько энергии, что без кислородного дыхания им никак не обойтись. И кислорода им нужно много. А в эпоху “скучного миллиарда” содержание О2 в атмосфере почти наверняка не достигало 10% от современного уровня — минимума, который принято считать необходимым для поддержания животной жизни. Правда, эта подозрительно круглая цифра, скорее всего, завышена: физиологические исследования показывают, что многие животные могут обойтись и меньшим количеством кислорода. Подобные оговорки, однако, не мешают признать, что старая идея Нерселла как минимум не противоречит современным данным: предполагаемое начало эволюции многоклеточных животных весьма приближенно, но все-таки совпадает по времени с новым ростом концентрации атмосферного кислорода в конце протерозоя. Это просто не могло не стать фактором, облегчившим появление животных: в конце-то концов ясно ведь, что чем больше кислорода, тем для них лучше. Не надо только считать кислородный фактор строго единственным. Будем помнить, что и во времена, когда кислорода стало сколько угодно, никаких многократных попыток создания многоклеточности животного типа не отмечается. Этот эксперимент удался природе только один раз.
Уютная эпоха “скучного миллиарда лет” могла бы длиться еще долго, если бы в биологию не вмешалась география. Драматические события, героем которых стала сама планета, привлекали внимание ученых на протяжении полувека, но только лет 15 назад информацию о них удалось сложить в более-менее цельную картинку. Бросим на эту картинку беглый взгляд, начав, как и положено, с начала.
“Земля-снежок”: пролог
В 1964 году английский геолог Брайан Харленд опубликовал статью, в которой констатировал, что абсолютно на всех континентах есть следы древнего оледенения, относящегося к одному и тому же времени — позднему протерозою. Как раз в начале 1960-х геологи научились определять прошлое положение континентов с помощью данных о намагниченности горных пород. Харленд собрал эти данные и увидел, что убедительно объяснить их можно только одним способом: предположив, что позднепротерозойское оледенение охватило сразу все широты Земли, то есть было всепланетным. Любые другие гипотезы выглядели еще менее правдоподобными (например, пришлось бы предполагать немыслимо быстрое перемещение полюсов, чтобы все континенты по очереди накрывались полярной шапкой). Как сказал Шерлок Холмс во время поисков Джонатана Смолла, “отбросьте все невозможное, то, что останется, и будет ответом, каким бы невероятным он ни казался”. Именно так Харленд и поступил. Выпущенная им с соавтором обстоятельная, написанная прекрасным языком статья не претендует ни на какие сенсации — там просто-напросто честно изложены факты и выводы. И все же гипотеза о всепланетном оледенении была для большинства ученых слишком смелой. Поэтому популярность она получила далеко не сразу.
Буквально в те же годы теорией оледенений занялся известный геофизик, ленинградец Михаил Иванович Будыко. Он обратил внимание на то, что оледенение может саморазвиваться. Ледяной покров имеет высокую отражательную способность (альбедо), поэтому чем больше суммарная площадь ледников, тем большая доля солнечного излучения отражается обратно в космос, унося с собой тепло. А чем меньше Земля получает тепла, тем на ней становится холоднее, и площадь ледяного покрова в результате растет, повышая альбедо еще сильнее. Получается, что оледенение — это процесс с положительной обратной связью, то есть способный усиливать сам себя. А в таком случае должен существовать некоторый критический уровень оледенения, после которого оно будет нарастать, пока волны льда с Северного и Южного полюсов не схлопнутся на экваторе, полностью заключив планету в ледяной покров и понизив ее температуру на несколько десятков градусов. Будыко математически показал, что такое развитие событий возможно. Но он и понятия не имел, что в истории Земли несколько раз это действительно происходило! Потому что на тот момент Будыко и Харленд еще не читали друг друга.
“Земля-снежок”: основное действие
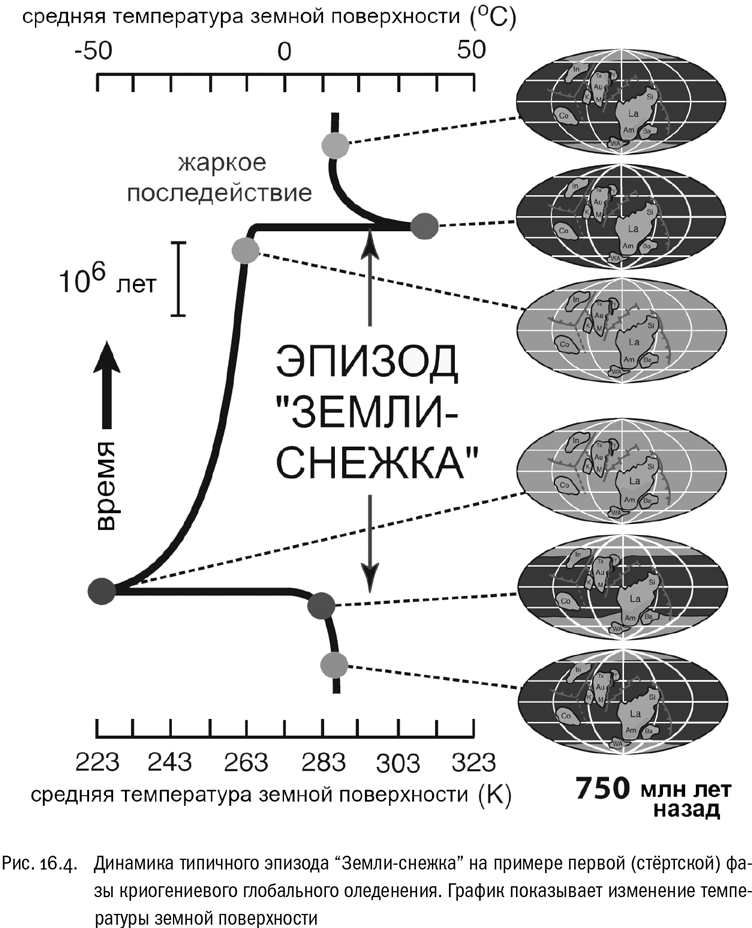
Сейчас оледенение, которое открыл Харленд, принято называть эпохой “Земли-снежка” (Snowball Earth). Судя по всему, оно действительно было всепланетным. А главной его причиной считается резкое ослабление парникового эффекта из-за падения концентрации углекислого газа (который стал главным парниковым газом после того, как кислород “съел” почти весь метан). Фотосинтез и дыхание тут, скорее всего, ни при чем. Если кислородную революцию биота Земли устроила себе сама, то теперь она оказалась жертвой внешнего фактора, совершенно небиологического по своей природе.
Дело в том, что оборот углекислого газа в целом гораздо меньше зависит от живых существ, чем оборот кислорода. Основным источником атмосферного CO2 на Земле до сих пор служат извержения вулканов, а основным стоком — процесс, который называется химическим выветриванием. Углекислый газ взаимодействует с горными породами, разрушая их, а сам при этом превращается в карбонаты (ионы HCO3– или CO32–). Последние отлично растворяются в воде, зато в состав атмосферы больше уже не входят. И получается предельно простая зависимость. Если интенсивность работы вулканов превосходит интенсивность химического выветривания, атмосферная концентрация CO2 растет. Если наоборот — падает.
На исходе “скучного миллиарда”, 800 миллионов лет назад, почти вся земная суша входила в состав единственного суперконтинента под названием Родиния. По словам известных геологов, гигантские суперконтиненты, как и крупные империи в социальной истории Земли, всегда оказывались неустойчивыми. Поэтому неудивительно, что Родиния быстро начала раскалываться. По краям разломов застывал извергнутый базальт, который сразу же становился объектом химического выветривания. А почвы тогда не было, и продукты выветривания легко сносились в океан. В конце концов Родиния распалась на семь или восемь небольших — размером примерно с Австралию — континентов, которые стали дрейфовать в стороны друг от друга.
Расход CO2 на выветривание базальта привел к падению его уровня в атмосфере. Вулканизм, которым распад суперконтинента неизбежно сопровождался, мог бы это скомпенсировать, если бы не одно случайное обстоятельство. В силу каких-то причуд дрейфа континентов и Родиния, и ее обломки находились у экватора, в теплом поясе, где химическое выветривание шло особенно быстро. Математические модели показывают, что именно по этой причине концентрация CO2 опустилась ниже порога, за которым начинается оледенение. А когда оно началось, останавливать выветривание было уже поздно.
Надо признать, что положение континентов в позднем протерозое оказалось настолько неудачным (с точки зрения обитателей планеты), насколько это вообще возможно. Дрейф континентов направляется потоками вещества земной мантии, динамика которых, по сути, неведома. Но мы знаем, что в данном случае эти потоки собрали всю земную сушу в единый континент, находившийся точно на экваторе и вытянутый по широте. Если бы он оказался на одном из полюсов или был вытянут с севера на юг, начавшееся оледенение закрыло бы часть пород от выветривания и тем самым приостановило уход углекислого газа из атмосферы — тогда процесс мог бы притормозиться. Как раз такую ситуацию мы наблюдаем сейчас, когда есть ледяные щиты Антарктиды и Гренландии. А в конце протерозоя почти все крупные участки суши находились близко к экватору — и оставались обнаженными до того самого момента, когда северный и южный ледяные покровы наконец сомкнулись. Земля стала ледяным шаром.
На самом деле эпизодов “Земли-снежка” было не меньше трех. Первый из них относится еще к гуронскому оледенению (которое, как мы помним, произошло не из-за углекислого газа, а из-за метана). Потом в течение более чем миллиарда лет никаких оледенений не было совсем. А затем последовало еще два разделенных небольшим перерывом всепланетных оледенения, одно из которых длилось примерно 60 миллионов лет, другое — примерно 15 миллионов лет. Именно их открыл Брайан Харленд. Геологический период, охватывающий эти оледенения, называют криогением (он является частью протерозойской эры).
О живой природе криогения известно относительно мало. Климат тогда на всей Земле был, по нынешним меркам, антарктическим. Большую часть Мирового океана покрывал километровый слой льда, так что интенсивность фотосинтеза не могла быть высокой. Свет, неожиданно ставший ценнейшим ресурсом, попадал в океан только местами, сквозь трещины, полыньи или небольшие участки тонкого льда. Удивительно, что некоторые многоклеточные организмы сумели пережить криогений, совершенно не изменившись, — например, красные водоросли. Они и сейчас приспособлены к тому, чтобы использовать очень слабый свет, проникающий на такую глубину, где уже не могут жить никакие другие фотосинтезирующие существа. Никуда не исчез и одноклеточный планктон. Содержание кислорода в криогениевом океане сильно упало, поэтому жизнь на его дне, скорее всего, была в основном анаэробной, но подробности этого от нас пока скрыты.
Окончания эпизодов “Земли-снежка” тоже были по-своему драматичны. Во время всепланетных оледенений все процессы, связанные с поглощением больших объемов углекислого газа, в буквальном смысле замораживались. А между тем вулканы (работу которых никто не останавливал) выбрасывали и выбрасывали CO2 в атмосферу, постепенно доводя его концентрацию до огромных величин. В какой-то момент ледяной щит уже не мог сопротивляться парниковому эффекту, и тогда начинался лавинообразный процесс разогрева планеты (см. рис. 16.4). Буквально за несколько тысяч лет — то есть геологически за мгновение — весь лед таял, освободившаяся вода заливала значительную часть суши мелкими окраинными морями, а температура земной поверхности, судя по расчетам, подскакивала до 50 °С. И только после этого начинался постепенный возврат Земли к “нормальному” для нее внеледниковому состоянию. За время криогения весь этот цикл был пройден минимум дважды.
Необычный мир венда
Эпоха “Земли-снежка” закончилась 635 миллионов лет назад. Начался последний период протерозоя — эдиакарий (635–542 млн лет назад). Теперь нам будет удобнее вести счет времени не на миллиарды лет, а на миллионы — это наглядно показывает, насколько события ускоряются. Хотя, возможно, дело просто в том, что они к нам ближе и от них сохранилось больше следов.
Раньше эдиакарий называли вендом, в честь нескольких групп древних индоевропейских племен — венедов (от этого же слова, по-видимому, произошло название города Венеция). К сожалению, сейчас это красивое наименование сохранилось лишь как нестрогий синоним.
Главным событием эдиакария (нельзя не добавить: с нашей антропоцентричной точки зрения) следует назвать появление многоклеточных животных. Датировать это событие нелегко. В палеонтологической летописи эдиакария хватает свидетельств перехода к многоклеточности животного типа — правда, чем они более ранние, тем более спорные. Во второй половине эдиакария в изобилии появляются вендобионты — крупные (до метра длиной) загадочные существа с плоским дисковидным или листовидным телом, состоящим из множества однотипных повторяющихся “сегментов” (см. рис. 16.5). “Сегменты” здесь поставлены в кавычки, потому что сегментация вендобионтов почти наверняка не имеет ничего общего с сегментацией настоящих многоклеточных животных. Сам термин “вендобионты” предложил немецкий палеонтолог Адольф Зейлахер, который считал этих существ совершенно особой формой жизни — гигантскими многоядерными клетками.
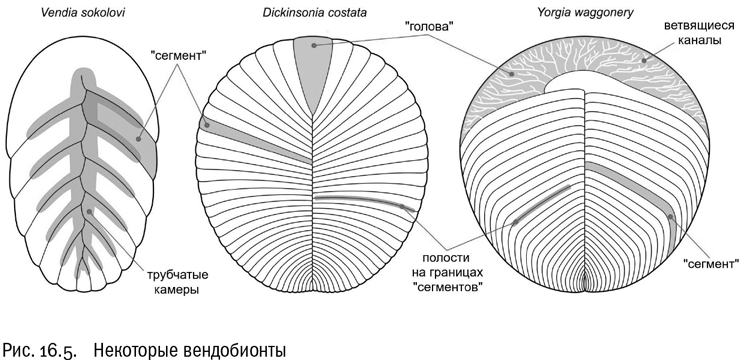
Действительно, есть серьезные основания считать, что вендобионты были во многих отношениях ближе не к многоклеточным животным, а к амебам или грибам (между прочим, и у тех и у других большие многоядерные клетки не редкость). Они совершили попытку выхода в крупный размер, которая сначала привела к успеху, но в итоге кончилась неудачей: в конце эдиакария вендобионты вымерли.
При этом надо учитывать, что вендобионты были очень разнообразны. Не факт, что их вообще можно хоть в каком-то приближении считать единой группой. Это скорее эволюционный уровень. И несмотря на то, что большинство вендобионтов никаких потомков не оставило, от кого-то из них вполне могли напрямую произойти некоторые современные животные — например, такие типы, как пластинчатые и гребневики . В эдиакарских корнях этих эволюционных ветвей нет ничего невероятного.
Самое древнее, абсолютно бесспорное ископаемое многоклеточное животное называется Kimberella quadrata (см. рис. 16.6). Это двусторонне-симметричное существо длиной до 15 сантиметров, ползавшее по морскому дну. Характер изменений формы тела найденных кимберелл (а найдено их много, в разных частях света) вместе с отпечатками следов не оставляет никаких сомнений, что они активно ползали, растягиваясь, сжимаясь и изгибаясь с помощью мышц. Характерные признаки кимбереллы — вытянутое, но компактное тело с ногой (мускулистой нижней поверхностью) и мантией (складкой, окаймляющей туловище). По этим признакам она очень похожа не на кого-нибудь, а на моллюсков. Есть мнение, что у кимбереллы была даже радула — свойственный моллюскам “язык” с хитиновыми зубцами, приспособленный для соскребания водорослей. Так или иначе это уже по всем статьям настоящее многоклеточное животное — с мышцами, нервной системой, ротовым аппаратом и вообще всем, что такому животному полагается иметь.
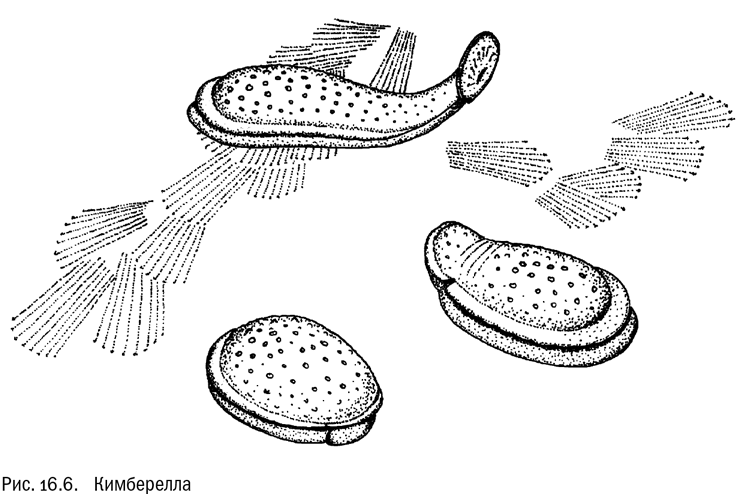
Кимберелла жила 555 миллионов лет назад. И примерно в это же время впервые появляются многочисленные ископаемые следы животных, явно активно ползавших по дну. Тут надо заметить, что “настоящие многоклеточные животные” — не особенно строгий термин. Сейчас нам достаточно условиться, что мы называем так животных с мускулатурой, нервной системой и ртом. У вендобионтов, насколько можно судить, ничего этого не было. Они питались в лучшем случае микроскопическими водорослями, а скорее всего — просто веществами, растворенными в морской воде. И только в конце эдиакария появились многоклеточные существа, способные активно отыскивать добычу и захватывать ее крупными кусками, чтобы переварить внутри. Вендобионты были перед такими чудовищами беззащитны — неудивительно, что их “золотой век” на этом закончился. В истории донных сообществ началась совершенно другая эпоха.
Назад: ЧАСТЬ IV ИСТОРИЯ ЖИЗНИ
Дальше: 17. фанерозой

