Книга: От атомов к древу. Введение в современную науку о жизни
Назад: Источники углерода
Дальше: ЧАСТЬ III ДРЕВО ЖИЗНИ
12. вирусы
На Земле развитие животного мира всегда шло от малого к большому, от простого к сложному. А вне Земли? В какой-то иной точке Вселенной развитие могло идти в обратном направлении, ко все более мелким формам. Подобно тому как прогресс человеческой техники ведет к миниатюризации многих вещей, так и эволюция на какой-то более совершенной стадии, вполне возможно, приводит ко все более и более мелким формам жизни.Майкл Крайтон. Штамм “Андромеда”
Двести лет назад великий французский биолог Жан Батист Ламарк писал, что всем живым организмам свойственно в ходе истории постепенно и неуклонно усложняться. Конечно, он был не прав. На самом деле биологическая эволюция может с примерно одинаковой вероятностью идти как в сторону усложнения организмов, так и в сторону их упрощения (и это останется верным, как бы мы ни определяли простоту и сложность). Фактор, который чаще всего направляет эволюцию в сторону большей простоты, — это паразитизм. Глубоко специализированные паразиты могут терять даже жизненно важные механизмы и свойства, иногда доходя в этом до крайностей. В главе 11 уже упоминались так называемые энергетические паразиты, которые лишены собственной системы выработки энергии и “воруют” АТФ у клеток, внутри которых живут. Энергетический паразитизм — не столь уж и редкое явление. Внутриклеточные энергетические паразиты есть даже среди эукариот: это микроспоридии, до неузнаваемости изменившиеся родственники грибов. У более простых организмов, например у бактерий, такой образ жизни встречается еще чаще. Ну а следующей ступенью упрощения после энергетического паразитизма, по логике вещей, мог бы стать паразитизм генетический.
Можно ли представить себе паразитическое существо, у которого нет собственной полноценной системы оборота генетической информации и которое, чтобы выжить, должно подключиться к генетическому аппарату хозяина, перенастроив его в своих интересах? Конечно, можно. Более того, таких существ на Земле великое множество. Они называются вирусами.
Внимание: ни в коем случае не надо думать, что вирусы (подобно множеству других паразитов) непременно произошли от организмов, которые были устроены сложнее их. Скорее всего, именно в случае с вирусами это неверно. Их происхождение — особое. Но тем не менее их тоже можно считать паразитическими живыми существами. Пусть и очень простыми (к вопросу о том, следует ли считать их живыми существами вообще, мы еще вернемся).
Вирус — это внутриклеточный паразит, состоящий из нуклеиновой кислоты (ДНК или РНК) и белка, который служит для защиты этой нуклеиновой кислоты и кодируется ею же. Только вот синтезировать этот белок самостоятельно вирус не может. Вирусы тем и отличаются от любых других живых организмов, что у них нет собственной системы трансляции, то есть синтеза белка. Свои системы репликации (копирования генома) и транскрипции (синтеза РНК) у вируса вполне могут быть, а вот к трансляции это не относится. В деле синтеза белка вирус целиком полагается на ту клетку, которую он заражает. Правда, у самых крупных и сложных вирусов иногда встречаются собственные белки, участвующие в процессе трансляции — но только в отдельных его этапах. Полной системы трансляции (и особенно рибосом, без которых трансляция невозможна) у них не бывает никогда. Это, пожалуй, самое главное отличие вирусов от клеток. Вирусы — единственная принципиально неклеточная форма жизни на современной Земле, да и вообще в известной нам части Вселенной, если уж на то пошло.
Другое важное отличие вирусов от клеток состоит в том, что вирусы никогда не размножаются делением. Каждая вирусная частица собирается заново из нуклеиновых кислот и белков, которые синтезируются совершенно отдельно друг от друга (очень часто — в разных местах зараженной вирусом клетки). Эта особенность размножения еще лучше подчеркивает, насколько сильно вирусы и клетки различаются между собой.
Все вирусы без исключения — облигатные внутриклеточные паразиты. Слово “облигатный” буквально значит “обязательный”. Облигатный внутриклеточный паразит — тот, кто не способен жить ни в каких других условиях, кроме как внутри чужой клетки. Вне клетки вирус всегда представлен только неактивной стадией — компактной частицей, которую для краткости называют вирионом. “Вирусная частица” и “вирион” — синонимы. Сам по себе вирион пассивен, “вести себя” он начинает только при встрече с живой клеткой; парадокс в том, что именно в этот момент он и теряет форму вириона — или даже любую форму вообще. Вирус, активно действующий в клетке, часто (хотя и не всегда!) бывает неотличим от ее собственного внутреннего содержимого.
Как бы ни были просты вирусы, у любого из них есть собственный геном. Там обычно закодированы белки, необходимые для репликации самого генома и для постройки новых вирусных частиц. Это — минимальный “функционал”. Но у некоторых вирусов, главным образом у крупных, геном обогащен множеством дополнительных генов. Большинство из них — это гены, продукты которых (то есть белки) так или иначе участвуют в многоэтапном взаимодействии вируса с клеткой хозяина.
На стадии вириона генетический материал вируса заключен в белковую оболочку, которая называется капсидом (см. рис. 12.1). Это чисто вирусное “изобретение”, у живых клеток никаких капсидов нет. У многих вирусов (но далеко не у всех) капсид имеет форму икосаэдра, то есть правильного двадцатигранника, одного из знаменитых Платоновых тел. Бывают и другие капсиды, например спиралевидные, нитевидные, палочковидные. Некоторые вирусные капсиды устроены очень сложно. Например, у миовирусов, которые паразитируют в клетках бактерий и архей, капсид (по словам одного известного вирусолога) похож на лунный посадочный модуль: он состоит из икосаэдрической головки, трубчатого хвоста с винтовой симметрией и нитевидных ножек, обеспечивающих “посадку” вирусной частицы на клетку. А есть вирусы, у которых капсид, в свою очередь, заключен в мембрану, похожую на клеточную: например, таковы вирусы СПИДа и оспы. Эту внешнюю мембрану, в отличие от капсида, у вирусов принято называть просто оболочкой.
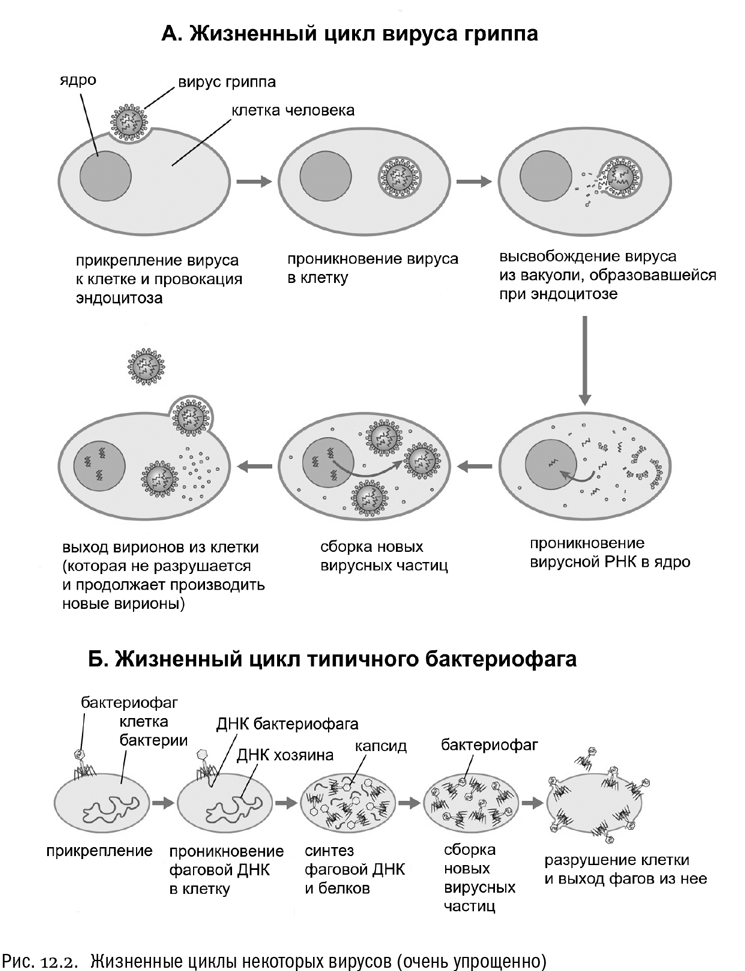
Разнообразие вирусных частиц связано, прежде всего, с разнообразием способов воздействия вирусов на живые клетки. Вирусы — великолепные манипуляторы. Например, вирусы животных часто попадают внутрь клеток, в самом буквальном смысле провоцируя эндоцитоз, то есть активный захват клеткой посторонних частиц (см. рис. 12.2А). (Частным случаем эндоцитоза является фагоцитоз, о котором мы говорили в главе 10.) Оболочка такого вируса содержит сигнальные белки, которые входят в контакт с рецепторами, сидящими в клеточной мембране, и заставляют их запустить в клетке механизм захвата. В результате поглощенный клеткой вирус оказывается у нее внутри, что, собственно, и требовалось. Это немного напоминает описанную Станиславом Лемом в “Звездных дневниках Ийона Тихого” охоту на курдля: охотник, намазавшийся специальной пастой и обсыпавшийся мелко накрошенным зеленым луком, проглатывается этим зверем, после чего устанавливает у него в брюхе бомбу с часовым механизмом.
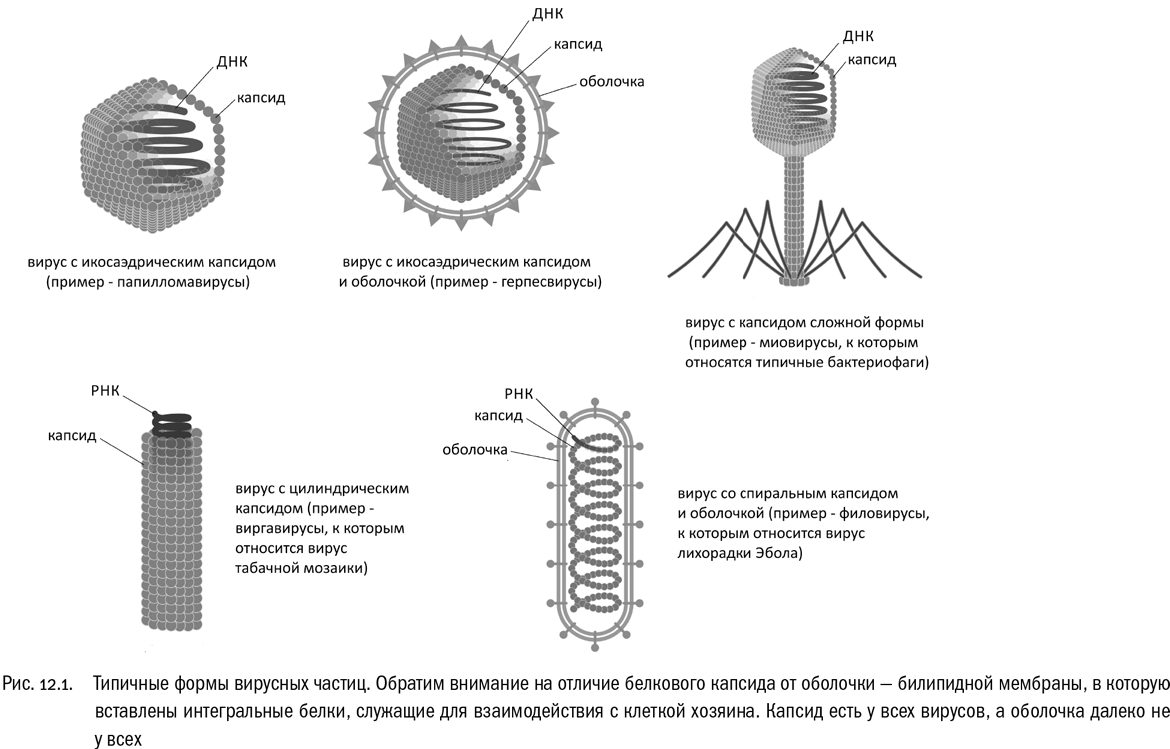
Вирусы, поражающие бактерий (так называемые бактериофаги), действуют не столь изощренно, но тоже по-своему изящно (см. рис. 12.2Б). Типичный бактериофаг закрепляется на клетке бактерии, проделывает отверстие в ее клеточной стенке и впрыскивает внутрь свою нуклеиновую кислоту, используя для этого механизм вроде шприца. Пустой белковый капсид вируса при этом остается снаружи. В простейшем, очень распространенном случае в бактериальную клетку вводится только чистая ДНК. Такой вирус и правда можно назвать “завернутой в белок скверной новостью”, как однажды выразился известный английский биолог Питер Медавар. Но мы еще увидим, что не со всеми вирусами дело обстоит так просто.
Вирусы есть абсолютно у всех клеточных организмов, включая животных, растения, грибы, водоросли, одноклеточных эукариот, бактерий и архей. Во всех без исключения случаях, когда биологи задавались вопросом: (есть ли у такого-то живого существа вирусы? — им удавалось найти если не сами вирусы, то уж по крайней мере генетические следы того, что в этом организме они когда-то присутствовали. А как правило, и сами вирусы тоже с успехом находятся. Живая природа Земли насыщена вирусами до предела. Более того, очень часто на одном и том же биологическом виде паразитирует сразу несколько разных вирусов. Мы отлично знаем это на своем собственном опыте — человека поражают десятки вирусных болезней, от желтой лихорадки до бешенства и от гриппа до СПИДа. А ведь есть и такие вирусы, которые вообще не вызывают никаких заболеваний, и их тоже весьма немало. Ведь в некотором смысле вершина эволюционного успеха вируса достигается тогда, когда он перестает убивать хозяина и вообще доставлять ему неприятности, а заражает всю популяцию как безвредный сожитель. У человека таких вирусов хватает, и их настоящую биологическую роль еще предстоит оценить.
Неудивительно, что вирусов во всех смыслах очень много. Например, литр океанской воды содержит в среднем миллиард вирусных частиц. Клеточных организмов, даже самых мелких, там раз в десять меньше. Общее число вирионов на планете Земля составляет 1031 (по грубым подсчетам). Это на несколько порядков больше полного числа звезд в наблюдаемой части Вселенной, которое, по мнению астрономов, равно примерно 1024.
И наконец, у вирусов невероятно разнообразны способы хранения и передачи генетической информации. Клеткам такое разнообразие, что называется, и не снилось. Да и ученые не сразу поверили, что оно возможно. Что ж, взглянем на это разнообразие поближе.
Генетические стратегии
Начнем с того, что любой вирус обязательно так или иначе взаимодействует с какой-нибудь клеткой. Он встраивается в ее генетический аппарат, переключая некоторые (а иногда и все) функции на себя. Поток генетической информации, идущий в самой клетке, в целом описывается уже известным нам заклинанием “ДНК-РНК-белок”, со смыслом которого мы познакомились в главе 9. Вспомним вкратце, в чем там было дело. Генетическая информация постоянно хранится на ДНК, которая перед каждым клеточным делением удваивается — этот процесс, как мы знаем, называется репликацией. Когда информацию настает время пустить в дело, происходит транскрипция, то есть ее перенос с ДНК на РНК. В результате транскрипции могут синтезироваться разные виды РНК, из которых нам сейчас важнее всего информационная РНК (иРНК), — она несет информацию, необходимую для синтеза белка. Сам синтез белка иначе называется трансляцией, это сложный процесс, идущий на рибосоме с помощью довольно большого набора специальных молекул. А все эти процессы вместе описываются центральной догмой молекулярной биологии: формула “ДНК-РНК-белок” именно ее и выражает.
Как видим, аппарат передачи генетической информации в клетке достаточно сложен. И подключаться к нему разные вирусы могут совершенно по-разному.
Есть вирусы, генетический материал которых представляет собой готовую информационную РНК. Их называют вирусами с позитивным РНК-геномом, сокращенно (+)РНК. Такой вирус действует, пожалуй, наипростейшим способом из всех возможных: ему достаточно ввести в клетку свою РНК, пусть даже совершенно “голую”. Попав в клетку, эта РНК распознается хозяйскими рибосомами и приводит к синтезу вирусных белков. Только после этого начинается репликация вирусной РНК. Для нее нужен фермент РНК-зависимая РНК-полимераза, способный синтезировать РНК на основе другой РНК и имеющийся только у вируса (хотя синтезируют его “под диктовку” вируса рибосомы хозяина). И наконец, накопившиеся молекулы вирусных белков и РНК монтируются в новые вирусные частицы. К вирусам с позитивным РНК-геномом относятся, например, вирусы полиомиелита и желтой лихорадки. А также знаменитый вирус табачной мозаики, с которого в свое время началось открытие вирусов как таковых.
Есть вирусы, генетический материал которых представляет собой не саму информационную РНК, а ее “зеркальную” комплементарную копию. Они называются вирусами с негативным РНК-геномом, сокращенно (–)РНК. Такая РНК не распознается рибосомами — для них она бессмысленна. Поэтому обойтись введением в хозяйскую клетку чистой РНК такой вирус не может. Вместе с РНК он вводит туда РНК-зависимую РНК-полимеразу, уже запасенную в готовом виде внутри вирусной частицы. Этот фермент копирует вирусный геном, образуя в результате “нормальные”, то есть позитивные, молекулы РНК. А уж они связываются с рибосомами, диктуя им “инструкции” для синтеза вирусных белков (в том числе и той же РНК-зависимой РНК-полимеразы, которая здесь не только обеспечивает дальнейшую репликацию вируса в данной клетке, но и упаковывается в вирусные частицы, чтобы принять участие в следующем цикле размножения). К вирусам с негативным РНК-геномом относятся, например, вирусы гриппа, бешенства и лихорадки Эбола.
Есть вирусы, у которых РНК двуцепочечная (дцРНК). Она свернута в двойную спираль примерно так же, как обычно сворачивается ДНК, и состоит из “позитивной” и “негативной” цепей, комплементарных друг другу. Жизненный цикл этих вирусов в целом похож на жизненный цикл вирусов с негативным РНК-геномом: в хозяйскую клетку сразу вводится РНК-зависимая РНК-полимераза, которая синтезирует как (+), так и (–)РНК. В конце клеточного этапа жизни такого вируса РНК собирается в двуцепочечные комплексы, которые — как и у всех вирусов — одеваются белковыми капсидами. К вирусам с двуцепочечной РНК относится, например, ротавирус, часто вызывающий у человека кишечную инфекцию.
Общая черта перечисленных типов РНК-содержащих вирусов следующая: они паразитируют только на аппарате трансляции, не затрагивая клеточную ДНК. Таким образом, внутриклеточную цепочку передачи генетической информации они захватывают только частично. В этом смысле их паразитизм не очень глубок.
Кроме РНК-содержащих вирусов в природе есть и много ДНК-содержащих, у которых (как понятно из термина) генетическим материалом служит не РНК, а ДНК. Довольно часто вирусная ДНК бывает двуцепочечной (дцДНК). Такая ДНК ничем принципиально не отличается от обычной клеточной — а это означает, что, попав в клетку, она может точно так же подвергаться транскрипции. Что и происходит. В результате синтезируется вирусная информационная РНК, которая, связываясь с рибосомами, делает свою обычную работу: диктует последовательности белков, которые надо синтезировать. Только в данном случае эти белки — вирусные, со всеми вытекающими отсюда последствиями для клетки.
Разумеется, вирусная ДНК в зараженной клетке заодно и реплицируется. Ее репликацией занимается ДНК-зависимая ДНК-полимераза, а транскрипцией — ДНК-зависимая РНК-полимераза. Эти ферменты есть абсолютно в любой клетке, поэтому ДНК-содержащий вирус в принципе может обойтись без них, и некоторые действительно обходятся. Но бывают и более сложные вирусы, которые “предпочитают” кодировать эти ферменты в собственном геноме и синтезировать их самостоятельно (самостоятельно — в том смысле, что по собственной инструкции, но все равно, конечно, с помощью клеточных рибосом). Затем из ДНК и вирусных белков, как обычно, собираются новые вирусные частицы.
К вирусам с двуцепочечной ДНК относится, например, вирус оспы. Сюда же входят герпес-вирусы, папиллома-вирусы (вызывающие рак шейки матки и обычные бородавки) и аденовирусы (вызывающие простудные заболевания, то есть всем нам хорошо знакомую острую респираторную вирусную инфекцию — ОРВИ).
Вирусы с одноцепочечной ДНК (оцДНК) тоже существуют. При попадании ДНК такого вируса в клетку она первым делом достраивается до двуцепочечной формы — это делает ДНК-зависимая ДНК-полимераза, которая, как мы уже знаем, и без того есть в любой клетке. А дальше все происходит примерно так же, как и у тех вирусов, ДНК которых двуцепочечная. К вирусам с одноцепочечной ДНК относятся, например, парвовирусы, способные вызывать у человека инфекционную анемию и некоторые другие болезни.
Совершенно особую группу вирусов представляют собой ретровирусы. Геном этих вирусов состоит из одноцепочечной “позитивной” РНК, которая в данном случае служит основой для обратной транскрипции, то есть для синтеза своей ДНК-копии. Служащий для этого фермент — РНК-зависимая ДНК-полимераза, она же обратная транскриптаза, она же просто ревертаза (как знает каждый, читавший повесть Стругацких “За миллиард лет до конца света”). Это вирусный фермент, который должен попасть в клетку в готовом виде в составе вирусной частицы. Полученная с его помощью вирусная ДНК встраивается в геном хозяйской клетки (для этого есть еще один специальный фермент — интеграза) и начинает вести себя там примерно так же, как “родные” хозяйские гены: успешно реплицироваться, транскрибироваться, а после синтеза РНК и транслироваться. Только в данном случае все это делается в интересах вируса и приводит к сборке новых вирусных частиц. Самый знаменитый из ретровирусов, безусловно, вирус СПИДа, он же вирус иммунодефицита человека (ВИЧ). Кроме того, известно, что некоторые ретровирусы могут вызывать опухоли.
И наконец, бывают вирусы с двуцепочечной ДНК, которая сперва транскрибируется, а потом... транскрибируется обратно. В этом случае на основе вирусной ДНК сначала синтезируется обычная информационная РНК, а потом в дело включается вирусная ревертаза, которая создает на основе этой РНК обратный транскрипт (то есть вновь молекулу ДНК) и встраивает его в хозяйский геном. Дальше все происходит примерно так же, как и у ретровирусов. Но ретровирусы — по определению РНК-содержащие. Поэтому их ДНК-содержащие аналоги называются ретроидными вирусами или параретровирусами. Самый известный параретровирус — это вирус гепатита B. Отметим, что у него нет ничего общего с вирусами гепатита A и гепатита C: последние относятся к РНК-содержащим вирусам с позитивным геномом, а значит, имеют совсем иную природу. Разница между этими вирусами гепатита гораздо более существенная, чем, к примеру, разница между животными и растениями.
Итак, вирусы делятся на семь классов по генетическим стратегиям. Это очень логичная классификация, которую предложил еще в 1971 году — по меркам биологии, целую эру назад! — крупнейший американский вирусолог Дэвид Балтимор. Правда, в системе Балтимора классов было шесть (параретровирусы тогда еще не успели изучить), но в остальном она прекрасно выдержала проверку временем.
В то же время надо иметь в виду, что эта классификация все-таки неполна. Во-первых, бывают полноценные вирусы, которые в нее не вписываются: например аренавирусы, у которых на одной и той же цепи РНК могут сосуществовать участки с полярностью (+) и (–). Во-вторых, классификация Балтимора не охватывает все разновидности генетических паразитов — хотя бы потому, что она не относится к паразитам, которым не нужна трансляция. А такие существуют. Например, вироиды — небольшие одноцепочечные РНК, лишенные белковых капсидов и (что самое интересное) не кодирующие никаких белков, но тем не менее способные размножаться в условиях чужих клеток и вызывать некоторые болезни. Именно вироиды вызывают, скажем, веретеновидность картофеля. Есть и другие подобные явления, о которых лучше почитать в книгах, написанных профессиональными генетиками (например, в “Логике случая” Евгения Кунина). Мы здесь ограничимся разговором о классических вирусах — в основном из тех соображений, что “нельзя объять необъятное”.
Каждая из перечисленных генетических стратегий по-своему способствует выживанию вирусов (что и логично — иначе носители этих стратегий просто давно исчезли бы). Каждая из них имеет свои уникальные преимущества. Чтобы понять это, давайте представим себе вирус с позитивным РНК-геномом. Он устроен предельно просто; чтобы запустить инфекционный процесс, ему нужно ввести в клетку свою РНК и больше ничего. Очень изящный механизм, к которому, казалось бы, незачем что-то добавлять. Проблема, однако, в том, что РНК такого вируса будет вынуждена конкурировать за доступ к рибосомам с “нормальными” клеточными информационными РНК, которые весьма многочисленны и перед которыми у вирусной иРНК нет никаких особых преимуществ. И количеством тут не возьмешь: в природе часто бывает, что клетка инфицируется всего-навсего одной вирусной частицей. А вот вирус с негативным РНК-геномом вводит в клетку “зеркальную” РНК, на которой, как на типографской матрице, можно создать сколько угодно комплементарных ей полноценных информационных РНК (и фермент, способный это сделать, при ней тоже есть). После чего эти вирусные иРНК во множестве загружают клеточные рибосомы, значительно повышая шансы вируса на выживание. Таким образом, тут возникает дополнительный каскад усиления, которого у вирусов с позитивным РНК-геномом нет. На первый взгляд жизненный цикл вируса с негативным РНК-геномом может показаться ненужным усложнением, а на самом деле это полезная адаптация (причем ее описанная функция, скорее всего, еще и не единственная).
В любом случае, разнообразие генетических стратегий вирусов само по себе изумительно. В этом плане вирусы резко контрастируют с клетками, генетическая стратегия которых — на всех одна-единственная: генетическая информация хранится на двуцепочечной ДНК, которая и служит основой для транскрипции. А вот вирусы, похоже, реализовали все генетические стратегии, какие только вообще можно придумать, — за единственным исключением. Это исключение касается процесса трансляции. Можно утверждать, что в живой природе трансляция никогда не идет прямо с ДНК. Для нее всегда, при любой самой необычной генетической стратегии, используется посредник — информационная РНК. Возможно, тут мы видим единственный во всей этой системе фундаментальный запрет. Причем этот запрет — не химический, а чисто биологический. С химической точки зрения трансляцию напрямую с ДНК вполне можно вообразить, и более того — ее можно даже попытаться осуществить в искусственных условиях. Но в биологической эволюции эта возможность, насколько мы знаем, не реализовалась ни разу.
Живые или нет?
Обсуждение вопроса, живые вирусы или нет, мы начнем с часто цитируемого высказывания англичанина Питера Медавара, когда-то назвавшего вирус “завернутой в белок скверной новостью” (или, как иногда переводят, “дурной вестью в белковом конверте”). Что он, собственно, хотел сказать? Это легко выяснить. Приведенные слова есть в популярной книге, которая называется “Наука о живом” (с подзаголовком “Современные концепции в биологии”); эта интересная и полезная книга была написана Питером Медаваром в соавторстве с его женой Джейн, впервые вышла в 1977 году, а в 1983-м появился ее хороший русский перевод. О природе вирусов там сказано следующее:
“Совершенно неважно, будет ли вирус определен как живой организм или нет. Одни свойства вирусов, например их экстрактивность, заразность и изменчивость, склоняют нас к тому, чтобы видеть в них мельчайшие живые организмы, еще более мелкие, чем бактерии; другие же их свойства — особенно способность нарушать механизм синтеза в живой клетке таким образом, что он начинает производить бесчисленное множество их копий, — заставляет видеть в них пакеты генетической информации. И вместо того, чтобы быть живым, вирус оказывается всего лишь завернутой в белок скверной новостью”.
Ключевые слова тут, как видим, в начале: “Совершенно неважно, будет ли вирус определен как живой организм или нет”. Медавар не решает вопрос о том, куда следует отнести вирусы, — он отказывается его решать. Это типичный ход мысли ученого-позитивиста. Есть такие-то доводы за то, чтобы считать вирусы живыми, и такие-то доводы против, но поскольку этот вопрос все равно не имеет никакого практического значения, то тратить на него время (по мнению Медавара) не стоит. Как говорится, пусть в этом разбираются наши схоласты. И не более.
Сейчас, в XXI веке, некоторые биологи продолжают считать, что вирусы нельзя относить к живым существам. Правда, такие заявления сразу же вызывают бурные дискуссии. Никакого единодушия среди ученых по этому вопросу на данный момент нет. Количественно сторонники того, чтобы все-таки считать вирусы живыми, среди современных интеллектуалов, пожалуй, преобладают (проведенный при написании этой главы опрос в ЖЖ показал, что 70% читателей считают вирусы живыми и только 30% неживыми; судя по всему, это довольно типичное соотношение). Впрочем, часто от этого вопроса вообще отмахиваются, объявляя его бесполезным для дела — как Питер Медавар. Но думается, что это пренебрежение все же напрасно. Понятия живого и неживого слишком глубоко укоренены в системе нашего мышления, в том, что англосаксы называют емким словом mind. И это не случайность, а отражение структуры Вселенной, в которой жизнь занимает достаточно важное место. Пожалуй, даже хорошо, что на Земле есть объекты, про которые не сразу понятно, живые они или нет: это прекрасная “пограничная ситуация”, столкнувшись с которой люди волей-неволей возвращаются к исходным понятиям, чтобы прояснить их. Поступим так и мы. А заодно немного обсудим, что такое вообще жизнь: именно сейчас для этого наступил самый подходящий момент.
Итак, есть три главные причины, по которым вирусы в разное время отказывались относить к живому. Вирусы не могут самостоятельно размножаться, у них нет обмена веществ, и они (как многие раньше думали), скорее всего, представляют собой что-то вторичное по отношению к клеткам. Разберем эти доводы.
1. Неспособность самостоятельно размножаться
Действительно, ни один вирус не может воспроизвести себя в среде, где нет живых клеток. Эта несамодостаточность всегда была важнейшим основанием для того, чтобы исключить вирусы из живой природы. Ведь способность воспроизводить себя безусловно, одно из важнейших свойств живых объектов. Но что это, собственно, значит?
Мы знаем, что существуют молекулы, способные побуждать некоторые среды к созданию копий этих молекул. Такие молекулы называются репликаторами. Все системы, которые мы называем живыми, обязательно содержат те или иные репликаторы (обычно это молекулы ДНК, но в случае вирусов иногда и РНК). Любому репликатору просто по определению нужна для самовоспроизводства определенная среда, причем требования, предъявляемые к этой среде, часто бывают очень строгими. Набор всех возможных сред, которые данный репликатор может заставить создавать его копии, философ Дэвид Дойч назвал нишей данного репликатора (по аналогии с известным термином “экологическая ниша”). Любой репликатор может функционировать только внутри своей ниши. За ее пределами он будет представлять собой мертвую молекулу, не проявляющую никаких свойств, кроме обычных химических. Вирусный геном — это типичный репликатор, ниша которого (в данном случае) находится внутри чужой клетки. Ну и что тут особенного? Ведь и любой другой репликатор всегда нуждается в определенной среде, часто причудливой и специфичной. В этом смысле ни один репликатор не самодостаточен.
Уточним, что репликатором мы сейчас называем не вирус целиком (это было бы совершенно некорректно), а вирусный геном, сравнивая его с геномом клетки. Сам же по себе вирус — это отнюдь не голый репликатор, а целый организм, пусть и более простой, чем клетка. По определению, которое дает “Биологический энциклопедический словарь”, организм — это “любая биологическая или биокосная целостная система, состоящая из взаимозависимых и соподчиненных элементов, взаимоотношения которых и особенности строения детерминированы их функционированием как целого”. Вирус соответствует этому определению: как и клетка, он состоит из набора разнородных компонентов, сведенных вместе общей функцией. Уж во всяком случае у любого вируса помимо молекул, несущих генетическую информацию, есть белковый “футляр” для них, то есть капсид. С этой точки зрения между самовоспроизводством вирусного и клеточного генома нет принципиальной разницы: и тому и другому нужна внешняя среда, которую он частично сам формирует. (Можно ли считать организмами и относить к вирусам репликаторы, лишенные капсидов, мы сейчас обсуждать не будем, чтобы не уйти “в область безбрежного”. Во всяком случае, такое отнесение пока не общепринято.)
Что же касается представления о самодостаточности, то оно в биологии, честно говоря, иллюзорно. Очевидно, что ни один внутриклеточный паразит не самодостаточен — ведь он при всем желании не может самовоспроизвестись без участия другого организма. Но это можно сказать и про “обычных” паразитов (не внутриклеточных), а заодно и про хищников, и про растительноядных, и про поедателей мертвой органики. С этой точки зрения самодостаточным нельзя признать ни одно животное на свете. То же самое относится и к грибам, и к цветковым растениям (которые сплошь и рядом нуждаются в симбиозе с микоризными грибами и насекомыми-опылителями), и вообще к большинству живых организмов на Земле. В этом отношении вирусы не представляют собой ничего особенного.
2. Отсутствие метаболизма, то есть обмена веществ
Утверждение, что у вирусов нет метаболизма, будет верно только до тех пор, пока мы отождествляем вирус с его компактной расселительной формой — вирионом. Если же взглянуть на жизненный цикл вируса целиком, включая и ту форму, в которую он переходит внутри зараженной клетки, этот аргумент сразу исчезнет. У вириона действительно нет ни метаболизма, ни экспрессии генов. Но внутри зараженной клетки у вируса все это есть: там экспрессируются вирусные гены, реплицируется вирусная ДНК (или РНК) и идут запущенные вирусом химические реакции. Между тем ясно, что мы в любом случае увидим проблему объемнее, рассматривая не изолированную вирусную частицу, а полный жизненный цикл — от вириона до вириона следующего поколения.
Обсуждение этого вопроса имеет долгую историю. Проницательный австралиец Фрэнк Макфарлейн Бёрнет, едва ли не первым во всеуслышание заявивший о генетической роли ДНК (мы упоминали об этом в главе 8), выпустил еще в 1945 году целую книгу, красноречиво озаглавленную “Вирус как организм”. А в 1983 году американский исследователь Клаудиу Бандеа, уже опираясь на молекулярно-биологические данные, предложил осознанно взглянуть на жизненный цикл вируса так, как если бы это был жизненный цикл самого обычного живого паразита. Под этим углом зрения “взрослой” стадией вируса, несомненно, будет выглядеть его внутриклеточная форма: именно она питается, размножается и преобразует энергию. На этой стадии вирус проявляет все физиологические свойства живого организма. А вирион — это покоящаяся стадия, аналогичная неактивным зачаткам, с помощью которых обычно расселяются растения и грибы. С этой точки зрения вирусные частицы подобны, например, спорам гриба-дождевика или летучим пушистым семенам ивы, которые Ричард Докинз назвал “одноразовыми парашютами для ДНК”.
Особую наглядность эти соображения обрели после открытия в начале XXI века гигантских ДНК-содержащих вирусов. Такой вирус может заразить, например, пресноводную амебу — одноклеточного эукариота, который двигается, выпуская и втягивая ложноножки. В результате заражения в клетке амебы возникает так называемая “вирусная фабрика” — сложное образование, в котором вирусные белки подчиняют себе клеточный цитоскелет, митохондрии и элементы системы внутренних мембран. Получается совершенно оригинальная структура, не имеющая никаких аналогов в незараженной клетке и работающая только на воспроизводство вируса. В ней реплицируется вирусный геном и собираются новые вирусные частицы. “Вирусные фабрики” — очень широко распространенное явление, по крайней мере у вирусов эукариот. Выглядеть они могут по-разному, иногда скромно, а иногда и грандиозно (по клеточным меркам, конечно). “Вирусная фабрика” гигантского ДНК-содержащего вируса, возникающая внутри амебы, — это уж точно весьма впечатляющая конструкция, расположенная прямо посреди клетки и не уступающая размером ее собственному ядру. Бывали случаи, когда микробиологи даже принимали “вирусные фабрики” за ядра. Очевидно, “вирусная фабрика” — это и есть активная форма вируса, аналогичная зрелому живому организму.
У бактерий и архей нет ни цитоскелета, ни митохондрий, ни системы замкнутых внутренних мембран. Поэтому инфицирующие их вирусы не образуют различимых под микроскопом “вирусных фабрик”. Вместо этого они превращают в “вирусную фабрику” всю пораженную клетку целиком. Хозяйский геном при этом обычно полностью разрушается (или, по крайней мере, инактивируется), так что в клетке — правильнее сказать, в бывшей клетке — экспрессируется только геном вируса. Особое искусство использования хозяйской клетки выработали, например, некоторые вирусы цианобактерий — организмов, более знакомых нам как синезеленые водоросли. Поражающие их вирусы для краткости называют цианофагами. Существует вирус-цианофаг, который, разрушив зараженную клетку изнутри, начинает синтезировать в ней свои собственные фотосинтетические белки (от аналогичных белков цианобактерии они отличаются особо высокой устойчивостью к ультрафиолету). В итоге в разрушенной клетке создается новый аппарат фотосинтеза, обеспечивающий энергией процессы репликации вирусной ДНК и сборки вирусных частиц. И, таким образом, бывшая клетка цианобактерии превращается не более и не менее как в фотосинтезирующий вирус. Более яркую иллюстрацию существования вирусного метаболизма просто трудно представить.
Исторически сложились два разных подхода к вирусам. Одни авторы считают, что вирус как таковой (the virus “self”) — это не что иное, как компактная вирусная частица, то есть вирион. А другие авторы убеждены, что вирион и внутриклеточная “вирусная фабрика” суть равноправные стадии единого жизненного цикла, который надо рассматривать не иначе как целиком. Переходя от первого подхода ко второму, мы получаем заведомо более полную систему. Очевидно, что это уже большое преимущество. В конце концов, любой грамотный зоолог или ботаник согласится, что всегда лучше исследовать полный жизненный цикл интересующего нас организма, чем какую-то одну стадию (неважно, взрослую или нет). Этот подход вполне можно распространить и на вирусы. Более того, он распространяется на них сам собой. В конце концов, не случайно никто никогда не сомневался в том, что вирусами должны заниматься биологи, а не химики.
Представим себе внутриклеточного паразита, в жизненном цикле которого есть две стадии — вегетативная (питающаяся и растущая) и расселительная. Вегетативная стадия обладает метаболизмом, ростом и экспрессией генов, но существовать она может только внутри чужой клетки, потому что нигде больше для нее нет подходящей среды. Расселительная стадия метаболически неактивна, гены в ней не экспрессируются, и вообще никакие жизненные процессы не идут, но зато она благодаря плотной оболочке может перемещаться по планете на большие расстояния, заражая новых хозяев. У клеточного организма такая расселительная стадия называлась бы спорой. Во всей этой картине нет ровно ничего фантастического. Облигатные внутриклеточные паразиты, неспособные жить ни в какой другой среде и размножающиеся неактивными спорами, есть не только среди бактерий, но даже среди эукариот. Ну и что, собственно, мешает считать, что вирион — это спора вируса, а после проникновения в клетку он просто переходит в вегетативную стадию? Да ничего.
3. Вторичность вирусов по отношению к клеткам
В ХХ веке было широко распространено мнение, что вирусы являются не более чем побочными продуктами клеточной жизни — то ли “взбесившимися” фрагментами клеток, освоившими самостоятельное существование, то ли целыми клетками, которые перешли к паразитизму внутри других клеток и в результате до предела упростились. Однако современные исследования вирусных геномов показывают, что это почти наверняка неверно. Дело в том, что обнаружено довольно много специфически вирусных генов, не встречающихся ни в каких клетках, — например, гены, кодирующие белки вирусного капсида (для любой клетки они бесполезны). То же самое можно сказать о некоторых генах и белках, обеспечивающих вирусную репликацию. Биоинформатика довольно быстро выделила набор чисто вирусных белков — в основном обеспечивающих копирование вирусных генов и сборку вирусной частицы, — которые широко распространены в мире вирусов, но не найдены ни у одной клеточной формы жизни. Скорее всего, это означает, что ни у каких клеточных организмов этих белков и соответствующих им генов просто-напросто никогда и не было. А это, в свою очередь, приводит к выводу, что вирусы вовсе не произошли от клеток. Их генетическое разнообразие имеет самостоятельный источник — вероятно, не менее (если не более) древний, чем первая живая клетка.
Правда, гены и белки клеточного происхождения у вирусов тоже встречаются. Но тут есть очень важная тонкость. Вирусный геном довольно четко делится на генетическое “ядро” (общее для многих вирусов и обеспечивающее жизненно важные функции — репликацию, построение капсида) и генетическую “периферию” (разную у разных вирусов и обеспечивающую в первую очередь взаимодействие с клеткой хозяина). Так вот, клеточное происхождение имеют только (или, во всяком случае, почти исключительно) гены “периферии”. У крупных вирусов их может быть очень много, и тогда они составляют большую часть генома. Но это все равно гены “периферии”, только раздутой. А вот для генов “ядра” генома клеточное происхождение как раз не обнаруживается. Отсюда и сделан вывод, что изначально было очень древнее “ядро”, на которое потом уже наслоились разнообразные заимствования из клеток.
Добавим, что аргумент насчет эволюционной вторичности вирусов мог бы лишить их своего места в живой природе, только если бы оказалось, что вирусы — это “сбежавшие” фрагменты клеток, но не целые клетки, упрощенные до неузнаваемости. Если бы верным было последнее, это, наоборот, означало бы, что вирусы с эволюционной точки зрения должны быть отнесены к той же категории, что и клетки. Но в любом случае все эти гипотезы плохо подтверждаются современными данными и сейчас непопулярны. Скорее всего, современные вирусы никогда не были ни клетками, ни их частями. Это просто совсем иная форма жизни.
Подавляющее преобладание клеточных форм в той части живой природы Земли, которая доступна людским органам чувств (это важная оговорка), успело было приучить людей полагать по умолчанию, что клетка и есть жизнь. Например, крупный вирусолог Андре Львов отказывался считать вирусы живыми существами. Он даже свою Нобелевскую лекцию начал с утверждения, что все живые организмы состоят только из клеток. Но Львов говорил об этом полвека назад. С позиции наших современных знаний представляется, что строгое приравнивание “жизнь = клетка” сейчас может быть только произвольно установленной догмой. С тем же успехом можно было бы ограничить понятие живого, например, подвижными организмами (потому что только их “живость” заметна с первого взгляда), или автотрофными (потому что только их питание напрямую не зависит от других живых существ), или способными самостоятельно синтезировать АТФ (тогда пришлось бы объявить неживыми довольно многих внутриклеточных паразитов). Очевидно, что любое из этих ограничений было бы искусственным, то есть случайным с точки зрения природы.
Впрочем, произвольность и искусственность определения — сама по себе не такая уж беда. Гораздо важнее, чтобы определение было полезным инструментом, помогающим расширять кругозор, а не ограничивающим его заранее. Например, можно ли считать живым Океан, описанный в лемовском “Солярисе”? И что мы сказали бы на месте исследователей, которые его открыли?
Одно из самых популярных определений жизни было выработано североамериканским Национальным аэрокосмическим агентством (НАСА) специально на случай встречи с инопланетной жизнью, которую первым делом надо будет опознать в качестве таковой. Согласно определению НАСА, “жизнь — это самоподдерживающаяся химическая система, способная к дарвиновской эволюции”. Процесс, называемый дарвиновской эволюцией, требует четырех условий: самовоспроизводство, наследственность, изменчивость и конкуренция за субстрат. Самовоспроизводство — это, как мы знаем, по определению главное свойство любого репликатора. Изменчивость возникает из-за случайного характера движения молекул, в силу которого при копировании любых репликаторов неизбежна некоторая доля ошибок. Эти ошибки передаются репликаторам следующих поколений, то есть наследуются. В результате возникает разнообразие репликаторов, которые начинают конкурировать между собой за субстрат (например, за мономеры, из которых можно собрать новые полимеры). А при совпадении этих условий автоматически запускается естественный отбор и начинает работать механизм биологической эволюции, вскрытый когда-то Чарльзом Дарвином (лучшее современное описание этого механизма можно найти в книге Ричарда Докинза “Слепой часовщик”). Попросту говоря, одни репликаторы копируются быстрее — их становится больше, другие медленнее — их становится меньше. Вирусы эволюционируют именно так, поэтому, по определению НАСА, их следует считать живыми. Например, компьютерная программа в принципе тоже может эволюционировать по-дарвиновски, но она не является химической системой и поэтому не подходит под это определение. А вот вирусы подходят под него однозначно.
Другое определение жизни, менее распространенное, но тоже очень интересное, гласит, что жизнь — это система с двойным развитием. Тут имеется в виду следующее: живой организм всегда обладает индивидуальным развитием (жизненным циклом), любая стадия которого потенциально способна измениться. Сумма этих изменений складывается в историческое развитие (эволюцию). Таким образом, биологическая эволюция — это развитие второго порядка, то есть развитие развития. “Все и только живые системы обладают взаимообусловливающими друг друга процессами индивидуального и исторического развития”. По этому определению вирусы тоже следует считать живыми: у них есть жизненный цикл, являющийся субъектом эволюции.
Итак, мы видим, что вирусы соответствуют даже довольно строгим формальным определениям жизни — если уж вообще браться такие определения искать.
А надо ли их искать? Знаменитый французский вирусолог Патрик Фортерр имеет по этому вопросу особое мнение. Он совершенно справедливо замечает, что само стремление создать общее определение, характеризующее сущность жизни, есть идеалистический предрассудок, восходящий в конечном счете к философии Платона. Поиск такого определения как бы предполагает, что где-то в бестелесном мире существует идеальная форма жизни, платоновскими “тенями” которой являются все реальные живые объекты. Однако мир, с которым имеет дело современная наука, оказался гораздо ближе не к платоновскому царству абсолютных идей, а к гераклитовскому космосу, в котором все течет и изменяется. Сколько бы определений жизни мы ни придумали, это не отменит того факта, что исторически мы знаем только земную жизнь (включая вирусы). Вот на ней-то и надо сосредоточиться — по крайней мере, пока на других планетах не найдут нечто живое. Тогда и можно будет подумать, как расширить наше понимание жизни и стоит ли его расширять. А пока условимся, что мы называем живыми природные объекты планеты Земля, существование которых основано на активном копировании информационных макромолекул (это не определение, а просто-напросто кратчайшее описание). И будем стремиться не к платоновским абстракциям, а к исследованию того, что реально происходило и происходит на этой планете. С этой точки зрения жизнь есть не более чем эпизод земной истории.
Две великие империи
Латинское слово virus вообще-то значит “яд”. Например, змеиный яд и яд, которым смазывают наконечник отравленной стрелы, обозначаются в классической латыни именно этим словом. Ну а в конце XIX века биологи назвали вирусами болезнетворные агенты, обладающие тремя очень характерными особенностями:
* они невидимы под световым микроскопом;* они свободно проходят сквозь фильтры, предназначенные для задержки бактерий (отсюда устойчивое, хотя и совсем уж сейчас устаревшее выражение “фильтрующийся вирус”);* они не поддаются выращиванию ни на каких искусственных питательных средах.
Не приходится удивляться, что при таких вводных природа вирусов первое время была совершенно загадочна. Вирус мог оказаться мельчайшим живым организмом, а мог и просто ядовитой молекулой. Лет 100 назад биологи вполне допускали и то и другое.
В 1935 году американский биохимик Уэнделл Стэнли ухитрился не только выделить из табачного сока вирус табачной мозаики, но и кристаллизовать его — так, как если бы это была обычная молекула. В то время это произвело сенсацию, тем более что под микроскопом вирусы на тот момент еще никто не видел. Неудивительно, что, получив этот результат, Стэнли посчитал вирус просто белковой молекулой, пусть и имеющей кое-какие особые свойства. “Вирус табачной мозаики может рассматриваться как автокаталитический белок, способный использовать живые клетки для собственного умножения”.
Тем не менее постепенно биологам становилось ясно, что вирус — не простая молекула. Одну из попыток дать вирусам четкое определение сделал уже нами упоминавшийся Андре Львов. По Львову, вирус — это мелкий (не больше 200 нанометров) инфекционный агент, не обладающий автономностью и не способный размножаться делением, в отличие от живых клеток. (Для справки: нанометр — это миллионная часть миллиметра или, соответственно, 1/1000 часть микрона.) К этому Львов добавил, что любой вирус обязательно содержит помимо белка нуклеиновую кислоту, но только одного типа — или ДНК, или РНК. Последнее резко отличает вирусы от живых клеток, в состав которых (как тогда уже было хорошо известно) всегда входят оба типа нуклеиновых кислот. Это и понятно, иначе никакая клетка просто не смогла бы функционировать. А вот вирусу содержать в себе сразу оба типа нуклеиновых кислот не нужно.
В любом случае такие взгляды подразумевали, что вирусы — это по сути своей нечто более простое, чем клетки. Гораздо, качественно более простое. Иная ступень устройства природы, если угодно.
Вот это мнение и оказалось неверным. Скажем аккуратнее: не вполне верным. Потому что в 2003 году был открыт мимивирус.
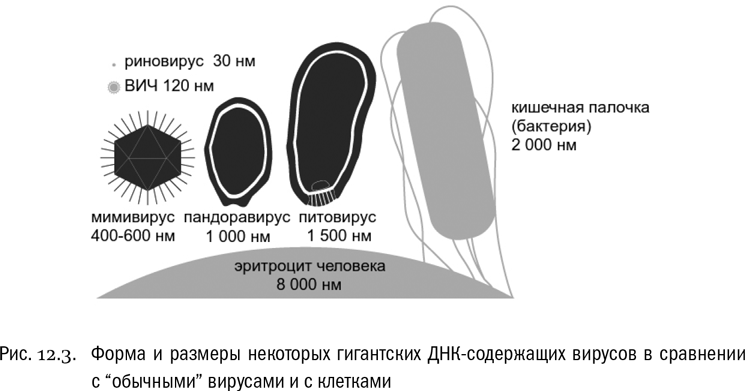
Выше мы уже упоминали это существо, хотя и не назвали тогда его по имени. Мимивирус — тот самый гигантский вирус, который паразитирует в пресноводных амебах и создает в них “вирусные фабрики”. Его вирион приближается к невероятному для вирусов размеру в 500 нанометров, то есть 0,5 микрона. Этот размер больше, чем у некоторых бактерий (среди последних есть внутриклеточные паразиты со средним размером 200–300 нанометров). Такой вирус вполне можно увидеть в световой микроскоп, а не только в электронный. И бактериальные фильтры его задерживают. Собственно говоря, поначалу мимивирусы за бактерий и приняли. Неудивительно, учитывая, что средний размер обычных вирусов — всего-то около 100 нанометров.
Все гигантские вирусы оказались ДНК-содержащими, причем ДНК у них двуцепочечная. Прочтение генома мимивируса показало, что этот геном состоит из 1,18 миллиона пар нуклеотидов и включает 979 генов, кодирующих белки. Это очень серьезные цифры. Для сравнения: вирус натуральной оспы — тоже крупный вирус с двуцепочечной ДНК (группа, к которой он относится, называется поксвирусами), однако его геном состоит только из 186 000 пар нуклеотидов и включает всего-навсего 197 генов. По вирусным меркам и это немало. Но у мимивируса геном в несколько раз больше. Он приближается по величине к геномам клеточных организмов — и не просто приближается, а вполне достигает их размеров.
Действительно, самый маленький полноценный архейный геном (он принадлежит одной морской архее) состоит из 490 000 пар нуклеотидов и включает 540 генов. А самый маленький полноценный бактериальный геном (принадлежащий паразиту микоплазме) состоит из 580 000 пар нуклеотидов и включает 475 генов. Как видим, эти геномы почти вдвое меньше, чем у мимивируса. Причем их обладатели не уникальны, одноклеточных организмов с подобными размерами геномов еще десятки. Многие из них — внутриклеточные паразиты, но не все.
Конечно, у большинства бактерий и архей (не говоря уж об эукариотах) геномы все же крупнее. Например, геном кишечной палочки состоит из 4,6 миллиона пар нуклеотидов и включает 4288 генов. (Тут учитываются только гены, кодирующие белки, но это сейчас неважно.) Вот это как раз “средняя” бактерия, ее геном не слишком велик и не слишком мал. И он, уж конечно, больше, чем у любого вируса, во всяком случае — насколько мы пока знаем.
Но ведь и мимивирус не уникален. Исследования быстро показали, что гигантских ДНК-содержащих вирусов в природе не так уж мало (см. рис. 12.3).
За прошедшие годы было открыто еще несколько близких друг к другу родов гигантских ДНК-содержащих вирусов. Они получили разнообразные названия: марсельвирусы, мамавирусы, мегавирусы, пандоравирусы, молливирусы, питовирусы и даже моумоувирусы. Недавно к этому списку добавились клоснойвирусы, индивирусы, катовирусы и хоковирусы. И каждый из них внес что-то свое в постоянно растущую сумму знаний о подобных созданиях.
У мамавируса, который тоже паразитирует в пресноводных амебах, в геноме 1023 гена. Одна из статей о нем так и озаглавлена: “Вирус, у которого больше 1000 генов”. Да, это действительно впечатляет.
У мегавируса число генов, подсчитанных тем же способом, достигает 1120 штук. Для сравнения: у возбудителя сифилиса — бледной трепонемы, которую никогда не считали ни карликовой, ни деградировавшей бактерией, — геном состоит из 1,14 миллиона пар нуклеотидов и включает 1039 генов. Чуть больше, чем у мамавируса, но чуть меньше, чем у мегавируса.
Еще позже были открыты пандоравирусы — огромные вирусы, тоже паразитирующие в амебах и отлично видимые под обычным световым микроскопом. Вирион пандоравируса, одетый мембраной и слегка напоминающий по форме античную амфору, вполне может быть длиной больше микрона. Геном одного из видов пандоравирусов состоит из 2,47 миллиона пар нуклеотидов и включает 2556 генов, кодирующих белки. Такой геном уже превосходит размером геномы не только многих бактерий, но и некоторых эукариот! Например, у микроспоридий — упоминавшихся выше эукариот, которые освоили внутриклеточный паразитизм, — геномы совершенно точно бывают меньше, чем геном пандоравируса.
В 2017 году был описан еще один гигантский вирус — клоснойвирус, названный по имени города Клостернойбург в Нижней Австрии. Его геном состоит из 1,47 миллиона пар нуклеотидов и включает 1545 генов. По нынешним меркам, это не рекорд — у пандоравируса, как мы знаем, геном еще крупнее. Но у клоснойвируса есть другая удивительная особенность. Его геном содержит необычайно большой (для вируса) набор генов, служащих для трансляции: 25 генов транспортных РНК и 19 генов ферментов-кодаз, которые “пришивают” к транспортным РНК транспортируемые ими аминокислоты (см. главу 9). Судя по всему, вирус получил эти гены из геномов своих хозяев — одноклеточных эукариот; в мире вирусов подобные захваты чужих генов вообще не редкость. Но вот уж чего-чего, а трансляции у вирусов быть не должно.
Впрочем, полного аппарата трансляции, способного к самостоятельной работе, нет и у клоснойвируса. У него полностью отсутствуют гены, кодирующие рибосомную РНК и рибосомные белки. Этих генов нет ни у одного известного вируса, что в общем-то и неудивительно: ведь именно отсутствие рибосом — это, как мы уже говорили, одно из самых главных различий между вирусами и клетками. По этому признаку никаких переходных форм между ними пока не видно.
Между прочим, в том же исследовании было заодно показано, что у разных эволюционных ветвей гигантских вирусов (например, у мимивирусов и у клоснойвирусов) наборы заимствованных генов совершенно разные. Общим у них оказалось только жизненно необходимое “ядро” генома. Тут напрашивается аналогия с компьютерной операционной системой: есть устойчивое “ядро”, а есть состоящая из множества программ-приложений “периферия”, которую можно легко менять и дополнять. Разнообразие этой “периферии” у гигантских вирусов показывает, что их эволюция шла очень быстро. Возможно, что и гигантами они стали независимо друг от друга, а их общий предок был куда скромнее.
Так или иначе открытие гигантских вирусов смело можно назвать одним из важнейших в современной биологии. Оно целиком датируется XXI веком (некоторые гигантские вирусы были известны и раньше, но тогда их принимали за каких-то странных бактерий). И тут замечателен не просто сам факт открытия новых, необычных организмов. Дело еще и в том, что открытие гигантских вирусов заметно изменило наши представления о структуре живой природы в целом. Конечно, никто не утверждает, что пандоравирус устроен сложнее кита-полосатика. Верхний предел сложности (даже чисто генетической) у клеточных организмов явно гораздо выше; это видно хотя бы по размерам их геномов. Но вот сказать, что любая клетка устроена сложнее любого вируса, теперь нельзя никак.
Итак, главный вывод: вирусы и клеточные организмы существенно перекрываются друг с другом как по размеру, так и по сложности. Это не две эволюционные ступени, а две ветви.
А теперь обратимся к роли личности в истории. Открытие гигантских вирусов, как и многие из дальнейших исследований на эту тему, связано с именем крупнейшего французского вирусолога и микробиолога Дидье Рауля (он тут уже упоминался). Рауль, конечно, работает далеко не в одиночку; самый известный его коллега — вирусолог Патрик Фортерр. Так вот, в некоторый момент Рауль и Фортерр написали совместную статью, в которой предложили новый взгляд на само понятие живого организма.
Будем исходить из того, что структура любого организма в достаточно неплохом приближении определяется его геномом. В некоторых ситуациях это утверждение может быть спорным, но в том масштабе, какой нам сейчас нужен, оно отлично работает. Можно сказать, что организм — это производное генома, фрагмент среды, которую геном организует вокруг себя. Итак, сравним геномы клеточных организмов (бактерий, архей и эукариот) с геномами вирусов. Чем они принципиально отличаются друг от друга? Во всяком случае, не величиной. Мы уже видели, что некоторые вирусные геномы превосходят числом генов некоторые клеточные, причем принадлежащие и бактериям, и археям, и эукариотам (число генов и размер генома на самом деле далеко не одно и то же, но сейчас это неважно: сравнение числа генов и сравнение размеров геномов, измеренных в парах нуклеотидов, в нашем случае дадут примерно один и тот же результат). Значит, разница между вирусными и клеточными геномами не количественная. А какая же?
Ответить на этот вопрос нетрудно, благо многие геномы сейчас уже полностью прочитаны. Любой вирусный геном отличается от любого клеточного (даже имеющего такой же размер) долями, которые отведены в нем некоторым строго определенным категориям генов. Бросается в глаза, что есть как минимум две важные группы генов, свойственных клеткам, но не свойственных вирусам. Это гены энергетического обмена и гены, ответственные за создание рибосом. У подавляющего большинства вирусов никаких генов энергетического обмена нет вообще, а если даже они есть, то кодируют только отдельные ферменты, но не полную систему синтеза АТФ (выше мы обсуждали такой пример — вирус-цианофаг, модифицирующий систему фотосинтеза в разрушенной клетке синезеленой водоросли). А по поводу рибосом Рауль и Фортерр замечают, что, судя по биоинформатическим данным, последний общий предок всех клеточных организмов имел как минимум 34 рибосомных белка, и эти белки (вместе с кодирующими их генами) сохранились у всех бактерий, архей и эукариот. У вирусов же нет ни одного из них.
Таким образом, бактерий, архей и эукариот можно с полным основанием назвать рибосомокодирующими организмами (ribosome-encoding organisms, REO). Этот признак четко отделяет всех их, вместе взятых, от вирусов.
Пока что мы выявили, так сказать, “негативные” отличия — констатировали, чего вирусы лишены. А есть ли отличия “позитивные”? Обладают ли вирусы чем-то таким, что ни у каких клеток не встречается? Да. В геноме любого вируса, заслуживающего этого названия, есть гены, кодирующие белки капсида — белкового “футляра”, заключающего в себе ДНК или РНК. Слово “футляр” тут, пожалуй, вводит в заблуждение, на самом деле вирусные капсиды частенько бывают довольно сложными структурами, построенными из разнородных частей и способными изменять свою геометрию (как, например, капсид бактериофага, который будто бы шприцем впрыскивает вирусный генетический материал в клетку бактерии). Вирус без капсида — это не вирус, а cубвирусная частица. Итак, особенность, которая есть у всех вирусов, и только у них, — это экспрессия генов капсида.
Таким образом, вирусы можно с полным основанием назвать капсид-кодирующими организмами (capsid-encoding organisms, CEO). Этот признак четко отделяет их от всех, кто состоит из клеток.
Человеческий разум любит дихотомии — отмечают Рауль и Фортерр. Деление организмов на рибосомокодирующие и капсид-кодирующие — это первичная дихотомия, на которой может основываться вся система живой природы. Она соответствует двум огромным эволюционным ветвям, разошедшимся на заре жизни, — двум ветвям, каждая из которых породила свой собственный биологический мир. Одни организмы “изобрели” рибосому и получили автономный аппарат трансляции — эффективный, но требующий постоянного снабжения энергией. А другие организмы сделали ставку на манипулятивное использование в своих интересах чужих аппаратов трансляции и на очень быстрое размножение. Сложностью при этом пришлось по большей части пожертвовать, но зато выживание генетического материала, рассеянного в чуждой и часто агрессивной среде, потребовало “изобретения” капсида.
Более того, не исключено, что заодно некоторые древние вирусы “изобрели” и ДНК. Ведь приспособительный смысл особенностей ДНК, отличающих ее от РНК, в том, что она чисто химически более устойчива (см. главу 8). А в условиях, когда вирионы переносились на большие расстояния водой или ветром, это могло быть особенно важно для сохранности их генетического материала. Между тем есть независимо обоснованная гипотеза, что первые клетки были РНК-содержащими — аналогично тому, как бывают РНК-содержащие вирусы. Это хорошо согласуется с тем фактом, что рибосомы со всем прилагающимся к ним набором РНК и белков появились раньше, чем клеточные механизмы репликации ДНК (во всяком случае, к таким выводам приводят данные сравнительной геномики). А если первые ДНК-содержащие вирусы уже существовали в эпоху первых клеток, то... Почему бы этим клеткам не получить “ноу-хау” хранения генетической информации на ДНК прямо от вирусов? Такой обмен генами вполне возможен. Тогда получается, что взаимодействие (или даже слияние) ДНК-содержащего вируса и РНК-содержащей клетки произвело на свет ДНК-содержащую клетку — ту самую форму жизни, которая сейчас доминирует на Земле.
Еще более вероятно, что некоторые клеточные организмы, а именно эукариоты, получили от вирусов механизм кэпирования информационной РНК (см. главу 10). Интрига тут вот в чем. Вирус, стремящийся подчинить себе крупную клетку, крайне заинтересован в том, чтобы его информационная РНК отличалась от обычной клеточной — например, несла на каком-нибудь своем конце специальную химическую метку. Тогда все молекулы иРНК, лишенные этой метки, можно будет разрушить, разобрав на отдельные нуклеотиды, а молекулы иРНК с меткой (принадлежащие вирусу) сохранить и направить на рибосомы, чтобы они дали вирусные белки. И в ходе эволюции вирусов такие метки действительно были “изобретены”. Например, у поксвирусов — крупных ДНК-содержащих вирусов, к которым относится вирус оспы, — к 5'-концу каждой иРНК “пришивается” мостиком из трех фосфатов особый модифицированный нуклеозид (гуанозин с добавочной метильной группой), причем “пришивается” он уникальным способом, как бы задом наперед. Поксвирусы — паразиты эукариот. А у самих эукариот, во всяком случае у современных, информационная РНК всегда помечается абсолютно так же! Собственно, эта-то метка и называется кэпом. Причем ни у бактерий, ни у архей кэпирования нет. Очень похоже, что эукариоты заимствовали этот механизм, вместе с обеспечивающими его генами, от своих вирусов.
Но зачем эукариотам это понадобилось? Ведь если бы кэпирование не приносило им никакой пользы, оно не удержалось бы в их геноме — во всяком случае, в работоспособном виде. Подсказку здесь дает наличие у эукариот, и только у них, особых ферментов, которые расщепляют свободную РНК на отдельные нуклеотиды, начиная с 5'-конца. Эти ферменты называются 5'-экзорибонуклеазами, и они стремятся разрушить любую информационную РНК, у которой на 5'-конце нет кэпа. Скорее всего, сочетание механизма кэпирования с присутствием 5'-экзорибонуклеаз является механизмом защиты клеток от РНК-содержащих вирусов, которые особенно склонны поражать именно эукариот (бактерий — гораздо реже). Вирусная РНК исходно не имела никакого кэпа, поэтому 5'-экзорибонуклеазы были обязаны ее атаковать, в то время как клеточные иРНК были защищены своим кэпом от такой атаки. Если все эти соображения верны, то получается, что один вирус (ДНК-содержащий) передал эукариотам гены ферментов кэпирования и тем самым подарил им способ защиты от других вирусов (РНК-содержащих). Ну а почему бы и нет?
Иное дело, что все современные РНК-содержащие вирусы эукариот, в свою очередь, успели приспособиться к этому защитному механизму и научиться его так или иначе обходить (например, вирус гриппа “ворует” кэп у клеточных иРНК, перенося его оттуда специальными ферментами). Так что эукариотам приходится создавать новые способы защиты от вирусов, основанные, например, на распознавании двуцепочечной РНК: самим эукариотам она не нужна, а вот в жизненном цикле большинства РНК-содержащих вирусов хотя бы короткая стадия дцРНК есть обязательно, и это хороший признак объекта, который нужно атаковать. Как сказал герцог Карл в финале пьесы Августа Стриндберга “Король Эрик XIV”, “борьба не кончается — никогда!”.
Однако вернемся к основной теме. По мнению Патрика Фортерра, современные РНК-содержащие вирусы, скорее всего, являются не чем иным, как остатками древнейшего РНК-мира, в котором РНК служила не только передатчиком, но и единственным носителем генетической информации (см. главу 9). Что касается ДНК-содержащих вирусов, то они почти наверняка произошли от РНК-содержащих. Переходной стадией между РНК- и ДНК-содержащими вирусами могли быть ретровирусы: у них как раз есть фермент, “переписывающий” генетическую информацию с РНК на ДНК.
По всей вероятности, первый вирус появился очень быстро — почти сразу же после появления первой рибосомы (независимо от того, возникла ли она в клетке или в какой-то доклеточной системе). Утверждать это можно с уверенностью, потому что возникновение паразитов неизбежно в любом эволюционном процессе. И эта неизбежность, как принято в таких случаях выражаться у ученых, имеет фундаментальный характер. Это означает, что она не является каким-то случайным “привходящим обстоятельством”, а логически вытекает из самой сути рассматриваемых объектов. В любом множестве репликаторов после нескольких циклов размножения (когда накопится случайная изменчивость) обязательно найдутся такие, которые тем или иным способом повысят скорость своей репликации за счет чужих ресурсов. Для систем, состоящих из самореплицирующихся РНК, этот вывод подтвержден математическими моделями. Но понять его легко и “на пальцах”. С точки зрения дарвиновской эволюции повышение скорости собственной репликации за чужой счет попросту слишком выгодно, чтобы упустить такую возможность. Именно поэтому во всех без исключения природных экосистемах есть огромное количество паразитов. Причем эти паразиты могут принадлежать к каким угодно эволюционным ветвям, от вирусов до высших растений и многоклеточных животных. Даже у гигантских вирусов, как мы теперь знаем, есть свои собственные паразиты — вирофаги, оказавшиеся ДНК-содержащими вирусами обычного среднего размера. Это наглядно показывает, насколько глубоко укоренен паразитизм в любой биологической эволюции, или, правильнее сказать, в биологической эволюции как таковой.
В древнейшем РНК-мире между предшественниками вирусов и предшественниками клеток не было никакой разницы. И те и другие реплицировались за счет рибозимов — особых РНК, обладающих каталитическими свойствами (см. главу 9). Рубежом, который резко отделил вирусные формы жизни от клеточных, стало появление трансляции. Приобретя систему трансляции, всякий организм получал возможность синтезировать по собственным инструкциям какой угодно набор белков — сложных молекул, которые, как мы знаем, являются великолепными инструментами для всевозможных взаимодействий с окружающим миром (см. главу 3). Но и цена этого приобретения была достаточно высокой. Система трансляции требует бесперебойного снабжения энергией и к тому же замедляет размножение — просто потому, что она слишком громоздка. Неудивительно, что в тогдашнем живом сообществе тут же нашлись любители пользоваться преимуществами, которые дает система трансляции, но не платить за нее положенную цену. Единственным способом сделать это, конечно же, оказалась манипуляция теми соседями, у которых система трансляции была честно установлена. С этой задачей первые вирусы (которых с этого момента уже можно так называть) вполне справились. А их главным преимуществом стала скорость размножения, доведенная до максимума за счет отказа от всего, от чего только можно отказаться. Этой стратегии вирусы успешно следуют и по сей день.
Эволюционным новшеством, маркирующим вирусы “в хорошем смысле этого слова”, стало, как мы уже знаем, создание капсида — футляра, защищающего генетический материал от разрушения внешней средой. Именно поэтому вирусы и называют капсид-кодирующими организмами. Впрочем, иногда против такого названия возражают — надо признать, что не без некоторых оснований. Дело в том, что в природе есть множество еще более простых паразитических репликаторов, которые представляют собой “голые” РНК или ДНК без всяких капсидов, но при этом имеют примерно такой же жизненный цикл, как и у вирусов. И мир этих репликаторов эволюционно связан с миром вирусов. В этом смысле выделение обладателей капсидов в особую категорию выглядит и вправду искусственным. Впрочем, с той или иной точки зрения искусственной выглядит любая система, стремящаяся разбить на четкие категории непрерывно меняющиеся природные объекты. От этого никуда не деться.
В оправдание классификации Рауля — Фортерра можно добавить, что главный объект биологии — все-таки организм, а “голый” репликатор считать таковым, пожалуй, нельзя. Биология вообще изучает историю организмов, а не генетических текстов; последнее — всегда лишь средство. И та система, в создании которой поучаствовали Рауль с Фортерром и о которой пойдет речь дальше в этой книге, — это именно система организмов. Репликаторы, не имеющие ни рибосом, ни капсидов, Рауль и Фортерр называют “сиротами” (orphan replicons). А граница между этими “сиротами” и настоящими вирусами, по их мнению, проходит по наличию структурных белков вириона. Вполне резонно.
В любом случае мы можем быть уверены, что миру клеточных организмов всегда, на всех этапах его развития и во всех эволюционных разветвлениях, сопутствовала виросфера — колоссальный невидимый мир вирусов и субвирусных частиц, скрытый от обычных органов чувств, но невероятно разнообразный и никогда не прекращавший свою стремительную, запутанную, бурную эволюцию. Иногда виросфера убивала, иногда внезапно делала ценные подарки, а иногда и манипулировала клеточными существами, подталкивая их собственную эволюцию в ту или иную сторону.
Полиднавирусы
У замечательного английского писателя Лоренса Даррелла есть роман “Месье, или Князь Тьмы”, входящий в пенталогию “Авиньонский квинтет”. Герои этого романа, живя в XX веке, сталкиваются с сектой египетских гностиков — продолжателей древних религиозных учений, согласно которым повелитель мира — не Бог, а Князь Тьмы. Гностики убеждены, что весь материальный мир, полный смерти, боли и распада, есть царство зла. Их предводитель, таинственный человек по имени Аккад, читает что-то вроде лекции, в которой поясняет эту мысль, пользуясь примерами из биологии:
“...Самка богомола, которая пожирает своего самца в то самое время, когда он ее оплодотворяет. Паук, который ловит муху в западню. Помпил, закалывающий паука ударом жала. Церцерис, который поражает тремя уколами три главных центра нервной системы златки, а потом его личинка поедает эту златку, еще живую, с чудовищной научной точностью обходя жизненно важные части, пока жертва не доедена до конца. А есть еще и наездники. И траурница, которая присасывается к личинке пчелы-каменщицы, постепенно выпивая ее досуха, и в конце концов съедает искусно сохраненный все еще живой остаток. И филант, убийца пчел, который, прежде чем утащить жертву в норку, давлением вынуждает ее извергнуть свой мед и сосет язык несчастного умирающего насекомого... Что за зрелище это Творение! Всеобщая бойня!”
Надо сказать, что все эти биологические примеры реальны. Скорее всего, Даррелл (как и его персонаж) взял их из книг великого французского энтомолога Жана Анри Фабра. Там можно найти рассказ и про помпила, и про церцериса, и про наездников, и про траурницу, и про филанта. И даже основное место действия “Авиньонского квинтета” по прихотливой случайности близко к местам, где Фабр вел свои исследования (он жил поблизости от Авиньона, на юге Франции).
Сам Фабр был типичным ученым-позитивистом, интересовавшимся только фактами (и одновременно — популяризатором науки, умевшим, надо отдать ему должное, блистательно эти факты описывать). Персонаж Даррелла — человек совершенно иного склада; открытия Фабра служат ему иллюстрацией гностического представления о материальной природе как о цепи существ, бесконечно пожирающих друг друга. “Природы вековечная давильня”, как выразился русский поэт по сходному поводу. Именно из-за такого взгляда на мир символом гностицизма считается уроборос — змей, заглатывающий свой хвост (см. рис. 1.5).
Однако посмотрим на приведенные примеры с чисто биологический точки зрения.
Филанта иногда называют пчелиным волком. Это оса, которая убивает пчел и кормит своих личинок их трупами. А вот помпилы, церцерисы, наездники и траурницы поедают своих жертв живьем. Вернее, это делают их личинки. Ради питания личинок некоторые насекомые парализуют свою добычу точными уколами жала, превращая ее буквально в живые консервы; помпил поступает так с пауком, а церцерис — с жуком-златкой. У других, например у наездников, жала нет, но их жертвам от этого ничуть не легче.
Хищники это или паразиты? В мире насекомых между этими понятиями нет четкого рубежа. Паразиты отличаются от хищников только тем, что их добыча крупнее их самих и нападение не является смертоносным. Паразитов на грани хищничества, которые постепенно потребляют все тело своего хозяина и в конце концов его убивают, принято называть паразитоидами. Некоторые паразитоиды относятся к мухам (например, траурница), некоторые — к другим группам насекомых. Но большинство из них — это осы. И помпилы, и церцерисы, и наездники входят в отряд перепончатокрылых, то есть являются осами, — конечно, в широком смысле этого слова.
Из всех этих существ вершины специализации к паразитизму достигли, пожалуй, наездники. Жала у них нет, вместо него — яйцеклад, иногда очень длинный, с помощью которого оса аккуратно помещает свои яйца внутрь другого насекомого. Из яйца вылупляется личинка, которая осваивает тело жертвы, поедая его постепенно, начиная с менее важных органов; жизненно важные она оставляет напоследок. Иногда жертва заражается одной личинкой наездника, а иногда и сразу несколькими — это зависит от вида.
В 1960-х годах канадский энтомолог Джордж Солт, детально исследовавший взаимоотношения наездника с организмом хозяина (в данном случае это была гусеница), выяснил, что действия самки наездника при заражении новой жертвы вовсе не сводятся к тому, чтобы просто отложить в нее яйца. Наездник — воистину совершенный паразит. Чтобы сделать организм жертвы комфортной для себя средой, он вводит туда некий яд, вызывающий (как мы сейчас знаем) удивительно многогранные физиологические эффекты. Во-первых, — это самое главное — подавляется иммунная система гусеницы, в норме активно стремящаяся уничтожить яйца паразита или, по крайней мере, не дать им развиваться. Без подавления иммунной системы жертвы наезднику придется плохо. Во-вторых, блокируется метаморфоз. Гусеница, пораженная наездником, никогда не превратится в бабочку, даже если чудом выживет. В-третьих, обмен веществ гусеницы слегка модифицируется, чтобы личинке наездника было выгоднее питаться ее внутренностями (особенно жировым телом, в котором у насекомых накапливается важнейший запасной углевод — гликоген).
А еще через несколько лет биологи обнаружили, что некоторые наездники зачем-то вводят внутрь гусеницы огромное количество плавающих в растворе вирусных частиц. Этот раствор впрыскивается в тело жертвы вместе с яйцами, через яйцеклад. Очень скоро было экспериментально показано, что именно загадочные вирусы и вызывают в организме гусеницы физиологические изменения. Получается, что в качестве яда наездник использует вирус. Не химическое оружие, а биологическое! (Иногда вместе с вирусами в тело жертвы вводятся и чисто химические яды, но в таких случаях они только дополняют действие вирусов, а не наоборот.)
Вирусы, которые наездник вводит в организм гусеницы, были вскоре выделены и описаны. Называются они полиднавирусами. Это название возникло как сокращение от довольно неуклюжего словосочетания “полидисперсный ДНК-содержащий вирус”. Использование полиднавирусов в качестве “биологического оружия” характерно для двух семейств наездников — браконид и ихневмонид; связанные с этими семействами вирусы называются, соответственно, браковирусами и ихновирусами. Между ними есть мелкие отличия, но в главных чертах они очень похожи.
Полиднавирусы оказались вирусами с двуцепочечной ДНК, в общем-то довольно обычными на вид. Ни по размерам, ни по структуре они не представляют собой ничего уникального — вирусы как вирусы. Но вот геном полиднавирусов поначалу сбил исследователей с толку. Обычно весь вирусный геном заключается в одной крупной молекуле ДНК (если вирус ДНК-содержащий, конечно). Так вот, у полиднавируса такой молекулы не нашлось. Вместо нее внутри вирусной частицы оказалось множество (10–30 штук, в зависимости от вида) маленьких кольцевых ДНК, несущих какой-то странный набор генов. Мы уже знаем, что любому вирусу в первую очередь нужны гены, обеспечивающие репликацию и построение капсида. А их-то в кольцевых ДНК полиднавирусов как раз и не оказалось. И прошло еще некоторое время, прежде чем поразительный жизненный цикл этих вирусов был полностью выяснен (см. рис. 12.4).
Полиднавирус имеет двух хозяев. Один из них — это оса-наездник, второй — поедаемая личинкой наездника гусеница. Неудивительно, что их роли в жизни вируса (как и роли, которые играет вирус в их жизни) абсолютно различны.
Оказалось, что геном полиднавируса целиком “вмонтирован” в геном осы. Он копируется и передается из поколения в поколение вместе с обычными осиными генами, с точки зрения молекулярных механизмов не отличаясь от них ровно ничем. Получается, что у каждой особи наездника в ядре каждой клетки тела содержится полный вирусный геном, включенный в какую-то из хромосом (или рассеянный по разным хромосомам — может быть и так и так, для нас это сейчас непринципиально).
У самцов наездников вирусные гены, по-видимому, постоянно “спят”. У самок же эти гены приобретают активность только по достижении половой зрелости и только в одной строго определенной части организма, а именно в клетках выстилки яйцевода. В этих клетках собираются вирусные частицы и синтезируются многочисленные маленькие кольцевые ДНК, содержащие — внимание! — далеко не все вирусные гены, а только те, которые нужны вирусу для действия на организм второго хозяина, гусеницы. Судя по всему, эти гены происходят из генома самой осы, хотя за время эволюции, связанной с вирусом, они успели заметно измениться, подстроившись под него. В результате можно считать, что это гены вируса, заимствованные им у осы, а можно считать, что это гены осы, которые она в определенный момент своего жизненного цикла “одалживает” вирусу, используя последний в качестве действующего на жертву внешнего эффектора. Самое интересное, что и то и другое будет совершенно верно. Ген вовсе не обязан непременно принадлежать какому-то одному организму! (Желающим понять, откуда берется это утверждение, можно посоветовать почитать книги Ричарда Докинза “Эгоистичный ген” и “Расширенный фенотип”, в которых все великолепно объяснено; здесь мы в теорию вдаваться не будем, потому что ничего нового по сравнению с этими книгами все равно не сообщим.)
Итак, вместе с яйцами осы, в составе жидкости, наполняющей ее яйцевод, вирионы полиднавируса попадают в тело гусеницы. Вот тут-то вирус и начинает свою работу. В первую очередь он поражает клетки иммунной системы гусеницы, делая их неактивными и не способными уничтожать яйца паразита. Есть и другие эффекты, причем довольно разнообразные: например, вирус заставляет клетки гусеницы синтезировать специальный белок, блокирующий гормональный запуск метаморфоза. Ведь если гусеница превратится в куколку, а потом и в бабочку, это грозит спутать все планы живущей в ней личинке наездника. В общем, вирус и в самом деле работает послушным эффектором, делая все, чтобы превратить тело гусеницы в максимально комфортную для наездника среду.
А вот никакого размножения полиднавируса в теле гусеницы не происходит. Его полностью обеспечивает оса. Именно поэтому гены репликации и капсида, которые необходимы для создания новых вирусных частиц, не включаются в состав вирионов: они там не нужны. Того, что эти гены хранятся в геноме осы, более чем достаточно, чтобы вирус (если можно так выразиться) был уверен в собственном выживании.
Интересно, что известен по крайней мере один вид полиднавирусов, у которого внутри вирионов вообще нет никакой ДНК — только готовые белки. В этом случае абсолютно все генетические процессы идут в теле осы, а вирионы используются только для переноса непосредственно действующих молекул. Еще одна прекрасная иллюстрация того, что эффекты генов могут и не зависеть от границ организмов.
Вирусные частицы, внедренные в гусеницу, — это тупиковая ветвь жизненного цикла, они в любом случае вместе с гусеницей и погибнут. Воспроизводство полиднавируса происходит исключительно путем передачи из поколения в поколение его генома, включенного в состав генома осы. Иными словами, он передается точно так же, как обычные осиные гены. При этом на организм самой осы скрытый в ней вирус не действует вообще никак. Даже вирусные частицы там создаются только тогда, когда настает пора вывести их наружу вместе с яйцами, и только в тех клетках, которые находятся прямо на пути этих яиц.
Теперь посмотрим на всю эту ситуацию с точки зрения интересов участников. Поселившись в наезднике, вирус получает эффективнейший механизм для выживания своих генов: целый биологический вид, все 100% особей которого даже не просто заражены этим вирусом, а несут в своих хромосомах его полный геном. С точки зрения дарвиновской эволюции это огромный успех — для вируса. А для наездника? Если бы присутствие вирусных генов не приносило наезднику никакой пользы, тот, вероятно, давно избавился бы от них и уж наверняка отключил бы. Последнее в таких случаях происходит само собой: нарушающие работу гена случайные ошибки накапливаются в нуклеотидном тексте очень быстро, если естественный отбор их специально не вычищает. А раз вирусные гены долгое время остаются в осином геноме целыми и работающими, значит, они чем-то помогают выживанию осы. И мы уже знаем, чем. Поступившие из осиного яйцевода вирусные частицы активно осваивают организм обреченной гусеницы — по словам исследователей, “генетически колонизируют” его, готовя к заселению наездником. Собственно, без вирусного сопровождения личинка наездника и не разовьется: сами по себе защитные системы гусеницы обычно справляются с паразитом. Это означает, что полиднавирус просто-напросто необходим осе для выживания. А это, в свою очередь, вполне позволяет считать, что он стал частью ее организма.
Однако не только оса пользуется вирусом в своих интересах, но и вирус использует осу как машину для собственного воспроизводства. И правильно делает: ведь за десятки миллионов лет совместной жизни он довел ее до состояния, когда она просто не может существовать без его поддержки. Теперь ни у вируса, ни у осы выбора нет.
Надо заметить, что сложные серии взаимных манипуляций с разнообразными исходами вообще самое обычное дело во взаимоотношениях паразитов и хозяев. Известный японский вирусолог Сусуму Маэда в свое время выразил эту мысль чеканной фразой: “Вирус — это хозяин”. Почти как у Стругацких: “А милях в двадцати отсюда, если идти вдоль рва, находится область, где людей поработили пришельцы с Альтаира, разумные вирусы, которые поселяются в теле человека и заставляют его делать, что им угодно”.
Одна хорошая статья о паразитах насекомых, написанная в том же ключе, получила заглавие “Игры, в которые играют паразиты”. Это — явный парафраз названия знаменитой книги психоаналитика Эрика Берна “Игры, в которые играют люди”. Хотя по сути тут скорее отсылка к современной математической теории игр. Действительно, в случае с полиднавирусом перед нами типичная игра с ненулевой суммой. Три участника, двое из которых в результате сложных взаимных манипуляций выигрывают, и только третий — гусеница — в итоге не получает ничего. Горе побежденным.
Интересно, что жизненный цикл полиднавируса поразительно похож на жизненный цикл не кого-нибудь, а многоклеточного животного. Это сравнение, кажущееся парадоксальным, на самом деле бросается в глаза. В теле многоклеточного животного всегда выделяется относительно небольшая группа клеток, которые потенциально бессмертны: они становятся половыми и могут передать генетическую информацию следующим поколениям. Эти клетки называют клетками зародышевого пути. Все остальные клетки половыми стать не могут и умирают вместе с телом данной особи (если нет бесполого размножения, но оно у животных не слишком распространено). Эти клетки называют соматическими, от греческого слова σωμα — “тело”. Соматические клетки — это тупиковая ветвь жизненного цикла. Они заведомо смертны, несмотря на то что именно из них создаются все сложные органы — или, может быть, как раз по этой причине. Так или иначе функции передачи наследственной информации из поколения в поколение и создания системы эффекторов, взаимодействующих с внешней средой, у многоклеточных животных четко разделены.
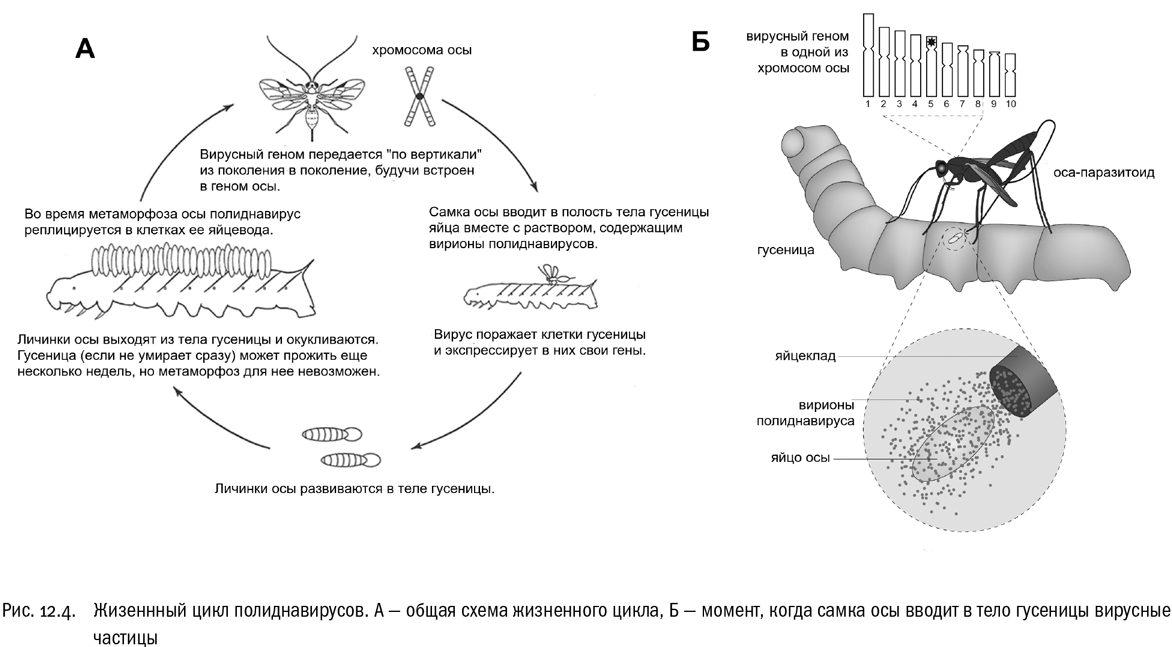
Но в точности такое же разделение этих функций выработали и полиднавирусы! У них есть “зародышевый путь” — скрытая форма вируса, заключенная в геноме осы, и есть система внешних эффекторов — вирусные частицы, набитые копиями специализированных генов или даже готовыми белками, которые атакуют гусеницу и затем погибают вместе с ней. Эти вирусные частицы — не что иное, как “соматическая” часть вируса, его смертное “тело”. Бессмертен у него только “зародышевый путь”. Перед нами разновидность жизненного цикла, вполне аналогичная жизненному циклу многоклеточных животных, но выработанная существами, которые и из клеток-то не состоят. Поистине инопланетная форма жизни; до такого и сам Станислав Лем не сразу бы додумался. В данном случае открытие ученых опередило фантастику.
Остается разобраться в том, как эта удивительная форма жизни вообще появилась. Впрочем, большой тайны тут нет. Можно не сомневаться, что в ходе эволюции у насекомых произошло постепенное “одомашнивание” изначально самого обычного ДНК-содержащего вируса, геном которого интегрировался в геном осы. Вирусов с двуцепочечной ДНК, паразитирующих на насекомых, известно достаточно много. В том, что кто-то из них стал полиднавирусом, нет ничего невероятного, известные генетические механизмы это вполне допускают.
Интересно другое: таких событий было как минимум два. Мы уже упоминали, что есть две большие группы полиднавирусов — браковирусы и ихновирусы, связанные с разными семействами ос. Так вот, эти группы вирусов не имеют общего происхождения. Во всяком случае, их общий предок полиднавирусом еще не был.
Все браковирусы, скорее всего, произошли от одного-единственного “обыкновенного” ДНК-содержащего вируса, геном которого некогда встроился в геном древней осы-бракониды. Генетики подсчитали, что интеграция этого вируса в осиный геном произошла примерно 103 миллиона лет назад. Причем общим у всех браковирусов — как и можно было ожидать — оказалось только “ядро” генома, состоящее из генов репликации и капсида. Геномную “периферию” они заимствовали у ос лишь недавно и во многом независимо друг от друга.
Об эволюционных корнях ихновирусов пока известно меньше, но есть все основания думать, что история их происхождения была примерно такой же. И это не случайное сходство. Тут мы видим прекрасный пример параллельного хода эволюционного процесса в двух близких, но уже успевших разойтись эволюционных ветвях.
Два интересующих нас семейства ос — бракониды и ихневмониды — разошлись примерно 150 миллионов лет назад, а вирусных сожителей и те и другие, надо думать, приобрели еще спустя десятки миллионов лет. Причем ни бракониды, ни ихневмониды не “охвачены” этими вирусами полностью. В обоих семействах есть много представителей, у которых никаких полиднавирусов нет — и, вероятно, никогда не было. Этот факт как нельзя нагляднее показывает, что обретение полиднавирусов внутри семейства браконид и внутри семейства ихневмонид произошло совершенно независимо.
Так или иначе сейчас полиднавирусами заражено — если тут применимо это слово — примерно 50 000 видов ос-браконид и примерно 14 000 видов ос-ихневмонид (в скобках отметим, что у всех этих видов до единого их, конечно, не искали — тут перед нами прикидки, сделанные на основании конфигурации эволюционного древа). Это означает, что все особи всех этих видов несут во всех своих клетках вирусные геномы. Впечатляющий эволюционный успех — особенно учитывая, что начаться он мог всего-навсего с двух индивидуальных вирусных частиц, которым повезло некогда встроить свои геномы в половые клетки двух древних ос. Вот так эволюция может усиливать случайные события.
Вирусы и эволюция
“Отнесение вирусов к неживой природе, долго преобладавшее в современной биологии, имело одно непредусмотренное последствие: большинство исследователей стало игнорировать вирусы при изучении эволюции”. И в самом деле: ну что такое вирусы для обычного зоолога или ботаника? Полумифические объекты, невидимые даже под микроскопом, и вообще непонятно, живые они или нет. А если неживые, то так ли уж важно биологу их знать? Казалось, что биологическая картина мира вполне могла бы обойтись без всяких вирусов. И действительно, в большинстве написанных в XX веке трудов на эволюционные темы упоминание вирусов (если оно там вообще есть) по духу своему похоже на подстрочное примечание, которое ставят для порядка, но к которому никогда больше не возвращаются.
После открытия реальной генетической роли вирусов маятник качнулся в другую сторону. Появилась тенденция считать (или, по крайней мере, молчаливо предполагать), что вирусы — величайшие творческие агенты и что мир вирусов таит в себе скрытые причины чуть ли не всех крупных эволюционных событий, какие только происходили в истории жизни на Земле. Неудивительно, что первыми так начали думать некоторые профессиональные вирусологи. В это русло попала, например, вирусная гипотеза происхождения клеточного ядра — впрочем, так и не ставшая общепринятой (см. главу 10).
Значение вирусов в живой природе, безусловно, огромно. Даже в роли простых возбудителей болезней они заметно влияют и на структуру экосистем, и на эволюцию своих жертв. Тут как раз нет ничего особенного: как говорится, “убивать и рукоед может”. Но ведь на самом деле отношения вирусов с клеточными организмами невероятно многогранны. К паразитизму и убийству они никак не сводятся. Вирусы и клеточные организмы могут, например, вступать в симбиоз, то есть взаимовыгодное сожительство. Мы обсудили всего лишь один случай такого симбиоза (полиднавирусов с осами), а на самом-то деле подобных примеров море. Чтобы описать хотя бы только те из них, что уже детально изучены, понадобилась бы отдельная книга. Симбиотические вирусы в большом количестве есть у бактерий, грибов, растений, животных — да в общем, у кого угодно. Хуже всех в этом плане изучены археи; но и вирусы архей, судя по всему, иногда участвуют в симбиотических отношениях. При этом “бонусы”, которые дают симбиотические вирусы своим клеточным партнерам, поразительно разнообразны. Синтезируя те или иные белки, вирус может защищать своего хозяина от других вирусов, или от бактерий, или от хищников, или от конкурентов за пищу, или помогать ему на ком-нибудь паразитировать, или регулировать его жизненный цикл, или понижать вероятность каких-нибудь совершенно незаразных болезней. Есть, например, вирус, который — внезапно — предотвращает заболевание мышей инсулинозависимым сахарным диабетом, потому что подавляет приводящую к нему аутоиммунную реакцию. Бывают и симбиотические союзы, состоящие более чем из двух участников: например, гриб, растение и вирус; или животное, бактерия и вирус; или животное и два вируса... Одним словом, творческие возможности природы тут колоссальны.
Совершенно исключительную роль в биологической эволюции играют ретровирусы. (И параретровирусы тоже, особенно в эволюции растений — но здесь мы не будем в это вникать.) Как мы знаем, главная особенность ретровируса состоит в том, что он может полностью интегрировать свой геном в геном хозяина, поместив его туда на правах обычных генов, принадлежащих клетке: что называется, как будто так и было. Иногда такое удается и другим вирусам, но у ретровируса это попросту является обязательной стадией жизненного цикла. Если ретровирус поражает соматические клетки, это не имеет никаких особых эволюционных последствий (например, такое происходит при СПИДе). Совсем иное дело, если клетка, в которую интегрировался геном ретровируса, оказывается клеткой зародышевого пути. Тогда вирус — а точнее, его геном — попадает в половые клетки и начинает передаваться по наследству, получая тем самым шанс на бессмертие. Это событие называется эндогенизацией ретровируса.
Довольно часто бывает, что образовавшийся эндогенный ретровирус становится нормальной частью генома целого вида животных. Его гены при этом могут “молчать”, а могут и экспрессироваться — с самыми разнообразными последствиями, от вредных до полезных. Эндогенные ретровирусы, накапливающиеся в геноме, образуют своего рода генетическую летопись древних вирусных инфекций. Ретровирусы, гены которых не экспрессируются вообще, в одной статье назвали “молчаливыми пассажирами” — действительно, у них встречается и такой способ выживания. Но не все они таковы. Эндогенные ретровирусы могут провоцировать болезни, а могут приносить хозяину и пользу — как повезет.
В силу своих особых свойств ретровирусы служат отличными агентами горизонтального переноса генов (про этот процесс мы упоминали в главе 10). Благодаря ретровирусам может происходить мгновенный обмен генами между организмами, относящимися к совершенно разным эволюционным ветвям. Впрочем, считать вирусы не более чем пассивными переносчиками чужих генов будет большой ошибкой. Дело в том, что у вирусов обычно очень быстро идет собственная эволюция, поэтому за время пребывания в геноме вируса захваченные им гены вполне могут успеть измениться до неузнаваемости, обретя совершенно новые функции. Бурная эволюция вирусов создает новые гены так активно, что иногда их называют “генными фабриками”. И во всех этих процессах ретровирусы играют не последнюю роль.
Особенно интересен в этом плане класс животных, к которому относимся мы сами — млекопитающие.
Дело в том, что разные классы вирусов предпочитают разных хозяев. Причины этого зачастую неизвестны, но факты остаются фактами. Например, большинство вирусов бактерий относится к вирусам с двуцепочечной ДНК. Грибам наиболее свойственны вирусы с двуцепочечной РНК, цветковым растениям — вирусы с позитивной одноцепочечной РНК. А вот млекопитающие почему-то пользуются особой “любовью” ретровирусов (хотя и другие группы вирусов не обходят их своим вниманием).
Уже довольно давно известно, что встроенные эндогенные ретровирусы составляют примерно 8% генома человека. Это более-менее рядовая величина; у большинства позвоночных доля эндогенных ретровирусов в геноме — от 4 до 14%. Надо заметить, что тут речь идет только об эндогенных ретровирусах как таковых, без учета других включенных в геном мобильных элементов, часть которых тоже наверняка имеет ретровирусное происхождение; если их добавить, цифры будут гораздо больше. Но так или иначе 8% генома человека — это тоже совсем не мало. Многие из входящих в эти проценты ретровирусных генов давно утратили активность. Но не все. Некоторые из них экспрессируются — с разнообразными последствиями, в том числе и неприятными.
Например, установлено, что эндогенные ретровирусы каким-то образом участвуют в развитии такой страшной болезни, как боковой амиотрофический склероз, которым страдал Стивен Хокинг. Как знать, может быть, мы когда-нибудь научимся и лечить подобные болезни, манипулируя вирусами? В конце концов, лечили же в свое время бактериофагами дизентерию и раневые инфекции. А современная биотехнология наверняка сможет создавать искусственные полезные вирусы с заданными свойствами. Но для этого надо будет сначала разобраться в тончайших скрытых механизмах, о самом существовании которых ученые узнали лишь недавно.
Есть у млекопитающих и полезные приобретения, связанные с ретровирусами. Например, это касается плаценты — органа, обеспечивающего питание зародыша за счет организма матери. К плацентарным относятся все современные млекопитающие, кроме сумчатых, утконоса, ехидны и проехидны, — иначе говоря, подавляющее большинство. Со стороны раннего зародыша в образовании плаценты принимает участие наружный слой его клеток, который называется трофобластом. А во внешнем слое самого трофобласта, непосредственно контактирующем с тканями матери, клетки обычно сливаются в единое многоядерное целое; видимо, так удобнее регулировать обмен веществ между двумя организмами. Многоядерный слой, образующийся из бывших клеток трофобласта, называется синцитиотрофобластом. Это одна из самых необычных структур в организме млекопитающих.
Не у всех обладателей плаценты есть синцитиотрофобласт. Например, у слонов и у большинства копытных его нет, и это — исходное для плацентарных млекопитающих состояние. А тем, у кого он есть, в определенный момент эмбрионального развития приходится обеспечивать слияние клеток трофобласта — изначально одноядерных, как и все другие клетки, — в единый протяженный слой с общей цитоплазмой.
Вот тут-то на помощь и пришли ретровирусы. Дело в том, что ретровирусы млекопитающих обычно проникают внутрь клеток, провоцируя слияние мембран. Каждая ретровирусная частица одета собственной мембраной (примерно такой же, как клеточная) и несет встроенные в эту мембрану сигнальные белки. Когда вирусная мембрана контактирует с клеточной, эти белки связываются с определенными клеточными рецепторами и передают сигнал, заставляющий мембраны просто-напросто слиться. И в результате генетический материал вируса оказывается внутри клетки, что ему и требовалось.
Точно такое же слияние мембран — только клеточных — происходит при возникновении синцитиотрофобласта млекопитающих. И происходит оно за счет тех же самых “провоцирующих” сигнальных белков (у млекопитающих они называются синцитинами). Эти белки — вирусные, они кодируются находящимися в геноме млекопитающих ретровирусными генами. Причем заимствование этих генов, с приданием им функции участия в образовании плаценты, произошло в эволюции млекопитающих независимо как минимум четыре раза: у обезьян Старого Света, у грызунов, у зайцеобразных и у хищных. А скорее всего, таких событий было еще больше. Человек в этом отношении не отличается от других обезьян Старого Света: у него есть синцитиотрофобласт, формирование которого обеспечивается ретровирусными белками-синцитинами. Вот так ретровирусы подарили млекопитающим полезный молекулярный инструмент.
Большинство эндогенных ретровирусов человека — общие с обезьянами Старого Света. Но не все. Есть по меньшей мере одна группа эндогенных ретровирусов, которая у человекообразных обезьян представлена слабо, а у человека — отлично. Эта группа называется HERV-K (human endogenous retrovirus K). Например, из десяти обнаруженных в человеческом геноме “встроенных” ретровирусов этого семейства всего два встречаются у африканских человекообразных обезьян — остальные восемь уникальны для человека. Значит, интеграция этих ретровирусов произошла только в человеческой эволюционной линии. С чем это связано — сказать пока не может никто. Известно, что гены HERV-K активируются при некоторых болезнях, в их числе рассеянный склероз, шизофрения и биполярное расстройство. Так что дело может быть и в нервной системе: все-таки она у человека отличается от обезьяньей. Так или иначе эта тайна еще не разгадана.
Итак, мы можем убедиться, что любая эволюционная ветвь клеточных организмов на всем протяжении своего существования отбрасывает густую вирусную “тень”. Роль вирусов в эволюционной драме поистине всеобъемлющая. Тем не менее эту роль не стоит (пока) преувеличивать, изображая вирусы какими-то невидимыми повелителями всего живого на Земле. На данный момент для такого мнения нет достаточных оснований. Вирусы не создали ни эукариот, ни плацентарных млекопитающих, ни ос-наездников, ни человека. Во всех этих четырех случаях (как, надо думать, и во многих других) они всего лишь ускорили эволюцию клеточных организмов в направлении, которое было к тому моменту уже выбрано.
Что вирус действительно может сделать — это помочь точно “притереть” какой угодно организм к своей экологической нише, снабдив его дополнительными молекулярными инструментами для полезных адаптаций. И конечно, организм всегда за это чем-то платит, так что процесс получается с обратной связью.
Великий эволюционный генетик Рональд Фишер в свое время сравнил биологическую эволюцию с тонкой настройкой оптического прибора — например, микроскопа. Представим, что мы смотрим в обычный микроскоп и хотим повысить качество изображения. Очевидно, для этого придется покрутить винты, чтобы поманипулировать расположением линз, углом наклона зеркала и другими доступными переменными. Причем этот процесс должен идти постепенно (от резкого перемещения линзы на большое расстояние вряд ли будет толк) и обязательно включать в себя обратную связь: подняв или опустив линзу, мы оцениваем: стало изображение лучше или хуже — и в зависимости от этого решаем, в какую сторону двигать ее дальше. Действуя подобным образом, мы почти наверняка довольно быстро получим требуемый результат.
Вот именно такую “тонкую настройку” клеточных организмов могут невольно осуществлять вирусы. И получается это у них прекрасно. Можно сказать, что вирусы вложили добавочный и очень острый резец в руку того самого слепого скульптора, с которым Станислав Лем (в который раз поминаемый в этой книге) сравнил однажды биологическую эволюцию.
Назад: Источники углерода
Дальше: ЧАСТЬ III ДРЕВО ЖИЗНИ

