Книга: От атомов к древу. Введение в современную науку о жизни
Назад: 7. нуклеотиды
Дальше: ЧАСТЬ II МЕХАНИЗМ ЖИЗНИ
8. нуклеиновые кислоты
Биохимией называют у нас те случаи, когда скверные химики занимаются грязными и плохими работами на малоподходящем для химии материале. Не это биохимия. Биохимия — это физико-химический структурный анализ активных макромолекул.Николай Тимофеев-Ресовский (цитируется по документальному роману Даниила Гранина “Зубр”)
Полимер, мономерами которого являются нуклеотиды, называется нуклеиновой кислотой.
Что можно сказать о таком полимере? Прежде всего, он в некотором отношении сложнее, чем состоящий из аминокислот белок, потому что любой нуклеотид — это (как мы теперь знаем) куда более сложная молекула, чем любая аминокислота. В нуклеиновой кислоте остатки сахара, принадлежащие разным нуклеотидам, соединены между собой через фосфатные группы. В результате получается длинная цепочка чередующихся остатков сахара и фосфата — так называемый сахаро-фосфатный остов, вбок от которого торчат азотистые основания. Представим эту картину, и можно считать, что общее представление о нуклеиновой кислоте мы уже получили.
Разные нуклеиновые кислоты называются по-разному в зависимости от того, какой у них в нуклеотидах сахар. Если это рибоза, то кислота рибонуклеиновая, а если дезоксирибоза — соответственно, дезоксирибонуклеиновая. Сокращения, обозначающие эти кислоты, — ДНК и РНК — вряд ли будут для кого-то из читателей большой новостью. В нашем перенасыщенном информацией мире про них труднее не услышать, чем услышать.
Объединить нуклеотиды в нуклеиновую кислоту в принципе очень просто. От гидроксильной группы, принадлежащей сахару одного нуклеотида, отщепляется водород (–H). От фосфата, принадлежащего другому нуклеотиду, отщепляется гидроксильная группа целиком (–OH). Эти отщепленные фрагменты образуют воду (H–O–H), а нуклеотиды соединяются по освободившимся связям. В результате между сахаром одного нуклеотида и фосфатом другого остается атом кислорода (–O–). Правда, в реальной живой клетке все происходит несколько сложнее (синтез нуклеиновой кислоты там начинается не с нуклеозидмонофосфатов, а с нуклеозидтрифосфатов). Но нам это сейчас неважно. Важно, что в итоге получается цепочка нуклеотидов, сахара которых соединены фосфатными мостиками.
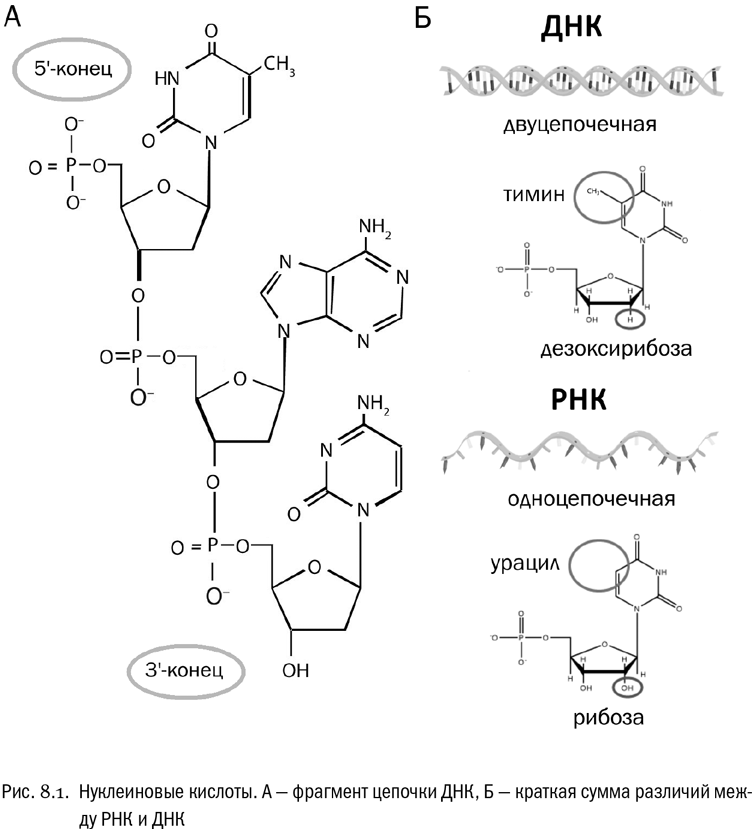
Теперь — серьезное дополнение (см. рис. 8.1А). Соединяющий нуклеотиды фосфатный мостик всегда расположен между 3'-углеродом одного сахара и 5'-углеродом другого. У возникающей цепочки нуклеотидов два конца, на одном из которых находится свободный фосфат (это 5'-конец), а на другом — свободная гидроксильная группа сахара (это 3'-конец; в устной речи прямо так и говорится — “пять-штрих-конец” и “три-штрих-конец”). Запомним это! Различать направления 5'→3 и 3'→5' очень важно: некоторые биологические функции нуклеиновых кислот без этого просто невозможно понять.
Хорошо еще, что ДНК и РНК — линейные полимеры, то есть неветвящиеся. Правда, чисто химически их ветвление вполне возможно, но современные живые организмы эту возможность почти нигде не реализуют.
Две кислоты
Между РНК и ДНК есть три главных отличия (см. рис. 8.1Б):
* ДНК имеет форму двойной спирали и используется для долговременного хранения генетической информации. РНК же почти всегда одноцепочечная и используется для передачи генетической информации, но не для ее постоянного хранения. Исключение можно найти только у такой странной (с нашей точки зрения) формы жизни, как РНК-содержащие вирусы.* Вместо рибозы в ДНК входит дезоксирибоза — сахар, имеющий на один атом кислорода меньше.* Из трех пиримидиновых азотистых оснований в РНК входят только урацил и цитозин, а в ДНК — только тимин и цитозин. Фактически в ДНК урацил заменен тимином.
Нет сомнений, что эти отличия должны иметь эволюционное объяснение, и довольно скоро мы попробуем его найти.
Исторически сложилось так, что поначалу в центре внимания биологов оказалась не РНК, а именно ДНК. В клетке ее обычно больше, и выделить ее для химического анализа легче. Кстати говоря, на самом деле природная ДНК — это не столько кислота, сколько соль. Кислотой ее называют из-за фосфатных групп. Но в условиях живого организма эти группы диссоциируют (отдают протоны), и остаются обнаженные отрицательные заряды, которые компенсируются положительно заряженными ионами натрия (Na+), благо натрий в окружающем растворе всегда есть. Так что с чисто химической точки зрения правильнее было бы называть ДНК дезоксирибонуклеатом натрия. И действительно, во многих старых классических работах ее именно так и называют. Но сейчас название “ДНК” настолько прочно вошло в язык, что менять его, видимо, уже не придется.
Главный источник ДНК в природе — ядра клеток эукариот. Как мы помним, эукариотами, собственно, и называются организмы с клеточными ядрами, то есть животные, растения, грибы и многие одноклеточные вроде инфузорий и амеб (см. главу 5). Что такое само клеточное ядро, мы пока “не знаем”, и нам не важны никакие детали его устройства, кроме самых простых фактов. Ядро — это находящаяся внутри клетки полость, окруженная оболочкой из двух мембран, в целом подобных наружной клеточной мембране, и содержащая очень много ДНК. Обычно клеточное ядро прекрасно видно под микроскопом. Само название “нуклеиновые кислоты” происходит именно от латинского слова, обозначающего ядро (nucleus). Вместе с белками, на которые она намотана, ядерная ДНК образует нити, которые тоже бывают хорошо заметны под микроскопом (эти нити принято называть хромосомами). Но и в клетках бактерий и архей, у которых нет никакого ядра, ДНК тоже всегда есть. Собственно, она есть абсолютно в любой современной живой клетке. Исключением могут быть только заведомо обреченные на скорую гибель клетки вроде, например, наших красных кровяных телец — эритроцитов (которые, надо заметить, и клетками-то не все биологи согласны считать).
К тому же молекулы ДНК просто огромны. У эукариот они могут состоять буквально из миллиардов нуклеотидных звеньев. Это самые длинные полимеры, какие только существуют в живой природе.
В общем, не заметить такое вещество было бы трудно. Но вот что и как оно делает — оставалось загадкой в течение нескольких десятков лет. Сейчас мы посмотрим, как эта тайна постепенно раскрывалась.
Тут не обойтись без небольшого авторского вступления. Эта книга ни в коем случае не труд по истории науки. Принятый в ней порядок изложения — логический, а не исторический. Это означает, что рассказ, по возможности, ведется, исходя из логики природных явлений, а не из того, что и в каком порядке открывали разные профессора, жившие сотню лет назад или еще раньше. В конце концов, наша главная цель — узнать что-то о современной биологии. Извилистые пути, пройденные научной мыслью давным-давно, тут могут подождать. И все же ради истории исследований ДНК хочется сделать исключение. Эта тема так важна и в то же время так поучительна, что мы позволим себе на ней остановиться — хотя бы кратко, в виде сжатого обзора, фиксирующего главные вехи.
Первый этап: открытие
Само существование ДНК открыл в 1869 году швейцарец Фридрих Мишер. Это открытие ни в коей мере нельзя назвать случайным. Фридрих Мишер, 25-летний на тот момент ученый, буквально чуть ли не с рождения вошел в научную элиту своего времени. Он был сыном профессора-медика, а его родной дядя — Вильгельм Гис — оказался выдающимся эмбриологом и анатомом, имя которого нередко упоминается в учебниках и сейчас, в XXI веке. (Каждый студент-медик знает, например, пучок Гиса, проходящий в продольной перегородке человеческого сердца.) Племянник и дядя дружили. И скорее всего, именно от Вильгельма Гиса еще совсем юный Фридрих Мишер воспринял мечту раскрыть самые фундаментальные тайны живой природы. В 17 лет он поступил на медицинский факультет, но работать практикующим врачом, судя по всему, не собирался ни дня. Ему просто нужна была хорошая естественно-научная база, чтобы приступить к поиску, как он говорил, “теоретических оснований жизни”. Мишер очень рано пришел к общему с Гисом убеждению, что “последние оставшиеся вопросы, касающиеся развития тканей, могут быть решены только на базе химии”. И он решил стать биохимиком. Правда, этого слова тогда еще не существовало, но было понятие “физиологическая химия”, означавшее то же самое. Поработав в великолепных немецких химических лабораториях, Мишер приобрел серьезную квалификацию химика-органика — и занялся изучением химического состава живых клеток.
Свой любимый объект — гной — Мишер обнаружил в хирургической клинике, по соседству с которой в тот момент работал. Из гноя оказалось очень удобно получать целые клетки, в первую очередь, конечно, лейкоциты — клетки иммунной системы, ответственные за воспаление. Именно из лейкоцитов Мишер и выделил вещество, обладавшее следующими четырьмя свойствами:
* оно всегда находится в высокой концентрации в клеточных ядрах, но почти отсутствует во внеядерной части клетки, так называемой цитоплазме;* его молекулы — большие, вполне сравнимые по размеру с молекулами белков;* оно определенно является по химическим свойствам кислотой;* оно состоит из углерода, водорода, кислорода, азота и довольно большого количества фосфора, но совершенно не содержит серы.
К тому времени биохимики уже знали, что в белках сера присутствует обязательно (как мы сейчас понимаем, она входит в состав некоторых аминокислот). А вот фосфора в них нет. Это со всей определенностью означало, что открытое Мишером вещество не белок, а нечто совсем другое.
Сам Мишер назвал это вещество “нуклеин”, от латинского слова nucleus — ядро. Через 20 лет Рихард Альтман переименовал “нуклеин” в “нуклеиновую кислоту”. Это название в науке и прижилось.
Мишер прекрасно понимал, что “нуклеин” не белок, и допускал, что это вещество выполняет какую-то особую функцию, свойственную только материалу клеточных ядер. Чтобы изучить химию нуклеина более детально, он использовал сперматозоиды — мужские половые клетки животных, в которых, кроме ядра, почти ничего толком и нет.
Между тем представление, что процессы передачи наследственной информации как-то связаны с клеточным ядром, к тому времени уже вошло в научный оборот. Это называлось ядерной теорией наследственности. Почему бы тогда “нуклеину” и не оказаться материальным носителем наследственных качеств? И действительно, в 1874 году Мишер записал: “Если бы мы предполагали, что какое-то одно вещество является специфической причиной оплодотворения, в первую очередь нам, несомненно, пришлось бы рассмотреть нуклеин”.
Второй этап: рутина
Фридриху Мишеру необыкновенно повезло. В своем стремлении раскрыть главную химическую тайну жизни он сразу выбрал абсолютно правильное направление поиска. Полученные им результаты подготовили науку к грандиозному прорыву. Но вот самого прорыва как раз и не произошло. В течение следующих 40 лет, примерно с 1890 по 1930 год, исследования нуклеиновых кислот оставались в общем-то непопулярной областью биохимии. Люди, которым хватало квалификации, чтобы ставить биохимические опыты, в этот период гораздо больше интересовались белками. Тогда уже было ясно, что белки — универсальные химические “слагаемые” жизни. В отношении нуклеиновых кислот такой уверенности не было даже у энтузиастов, при том что исследование этих веществ по чисто химическим причинам было заметно более трудоемким, чем исследование белков. Неудивительно, что желающих ими заниматься находилось относительно немного.
Правда, и в этот период у некоторых людей случались озарения. Вопрос о биологической роли ДНК не обошел стороной, например, известный физиолог Жак Лёб. В вышедшей в 1906 году книге под названием “Динамика живой материи” он совершенно четко сформулировал два предположения:
* наследственная информация при оплодотворении, скорее всего, передается каким-то одним строго определенным химическим веществом;* нуклеиновые кислоты являются гораздо более вероятными кандидатами на роль этого вещества, чем белки.
Лёб стремился сводить всю живую природу к простым физико-химическим факторам, и это, конечно, вызывало у многих биологов возражения, особенно когда применялось к таким сложным явлениям, как, например, поведение животных. Но именно в отношении передачи наследственной информации Лёб оказался абсолютно прав. Он, что называется, попал в яблочко. К сожалению, в 1924 году, когда ученый умер, оценить этого еще никто не мог. Предложенная им гипотеза о веществе наследственности просто затерялась среди множества других гипотез, казавшихся тогда не менее вероятными.
Американский биолог Леонард Троланд высказал другую смелую гипотезу: нуклеиновые кислоты — это своего рода небелковые ферменты, запускающие процесс копирования генетической информации. Гораздо позже стало понятно, что эта мысль не просто красива, но и (опять же) в немалой степени верна. У некоторых нуклеиновых кислот такая функция действительно есть.
Увы и увы, 100 лет назад все эти идеи разбились о полное равнодушие профессиональных генетиков, интересы которых в тот период были совершенно другими. Впрочем, самих генетиков тут тоже можно понять. Их юная наука, только в 1905 году получившая свое название, развивалась невероятно бурно — и охватить все возможные направления просто-напросто не получалось. А при этом ни концептуальный аппарат генетики, ни создавшие ей славу “фирменные” методы исследований изначально ни с какой химией связаны вовсе не были. И в результате генетики начала XX века практически единодушно решили, что поиск химического носителя наследственной информации — дело далекого будущего, а пока что на это отвлекаться не следует.
Таким образом, биохимики (вернее, те из них, кого это вообще интересовало) были вынуждены изучать ДНК в гордом одиночестве. Причем “сухой остаток” от этой долгой и трудной работы был довольно скромным. Стало понятно, что нуклеиновая кислота — это полимер, состоящий из нуклеозидов, соединенных фосфатными мостиками, то есть, иными словами, из нуклеотидов. И выяснилось, что нуклеотиды в ДНК бывают четырех типов: адениновый, гуаниновый, цитозиновый или тиминовый. Никаких далеко идущих выводов эти факты не породили. Ну, вещество себе и вещество. Ясно, что оно участвует в клеточном ядре в каких-то биохимических процессах, ну так мало ли там всего разного участвует! Где-то так, по-видимому, и думало большинство биологов к началу 1930-х годов.
Третий этап: споры
Между тем гигантский маховик под названием “развитие научных представлений” продолжал проворачиваться. В 1926 году американский генетик Герман Мёллер открыл радиационный мутагенез, то есть повышение частоты генетических мутаций под действием электромагнитных лучей, в данном случае — рентгеновских. Если попадание кванта рентгеновского излучения может изменить структуру гена, значит, ген — это молекула? А если молекула, то должна же у нее быть какая-то химическая формула! И таким образом, вопрос о химической природе наследственного вещества вновь встал на повестку дня.
В начале 1930-х практически все биологи, задававшиеся этим вопросом, считали, что гены — это белки. Почему? Во-первых, к тому времени уже все знали, что белки химически гораздо разнообразнее: в них входит 20 разных типов мономеров, а в составе ДНК — всего четыре типа. Во-вторых, биохимики, изучавшие ДНК в первой трети XX века, наряду со множеством полезных открытий допустили одну простительную, но тем не менее очень серьезную неточность. Они решили, что четыре типа нуклеотидов (адениновый, гуаниновый, тиминовый и цитозиновый) входят в состав ДНК в строго равных концентрациях:
[А] = [Т] = [Г] = [Ц]
Самое логичное объяснение этих данных выглядело так: ДНК, какой бы длины она ни была, должна состоять из одинаковых четырехнуклеотидных блоков, в каждом из которых есть по одному “А”, по одному “Т”, по одному “Г” и по одному “Ц”. Представить, что такой однообразный полимер каким-то образом хранит информацию, было невозможно. Приходилось считать, что ДНК — рядовой участник обмена веществ, разве что специфичный почему-то именно для клеточных ядер.
Впрочем, мнения и тогда бывали разные. В 1933 году работавший в США хорватский биолог Милислав Демерец опубликовал буквально витавшую к тому времени в воздухе гипотезу, что любой ген — это молекула. Пусть и большая молекула, но одна-единственная. Тогда генетическая мутация — просто изменение взаимного расположения атомов в этой молекуле. А в качестве примера того, из чего такая молекула могла бы состоять, Демерец привел не что иное, как схему гипотетического четырехнуклеотидного блока ДНК! Биология опять почти нащупала материальный носитель наследственности — и опять отступила (правда, теперь уже ненадолго). На этот раз биологов подвела химия. Структура нуклеиновых кислот была все-таки еще слишком плохо известна. К примеру, на предложенной Демерцом формуле ДНК значилась гликозидная связь между остатками сахара (см. главу 6), чего на самом деле ни в каких нуклеиновых кислотах не бывает.
Тут самое время вспомнить замечательного русского биолога Николая Константиновича Кольцова, занимавшего в интересующие нас годы должность директора Института экспериментальной биологии в Москве, на улице Воронцово Поле, что у Яузского бульвара. Кольцов — фигура ярчайшая, о его роли в нашей биологии написано немало статей и книг. Сейчас достаточно сказать, что он одним из первых задумался не только о химической основе наследственных качеств, но и о молекулярном механизме их передачи и тут сумел во многом, что называется, опередить свое время (см. главу 3). В 1935 году Кольцов опубликовал следующую гипотезу: ген — это участок очень длинной белковой цепочки, возможно состоящей из тысяч или десятков тысяч аминокислот, чередованием которых, собственно, и кодируется генетическая информация. Забегая вперед, сообщим, что если бы Кольцов заменил белок на ДНК, а аминокислоты на нуклеотиды, то он бы оказался попросту абсолютно прав. К сожалению, такого чуда не случилось. В тех же самых статьях Кольцов убежденно возражает Демерцу: ДНК — “сравнительно простое органическое соединение, которому было бы странно приписывать роль носителя наследственных свойств”. То ли дело белки! Приведем сокращенную цитату:
“Некоторые цитологи придают нуклеиновой кислоте особо важное значение. Так, Демерец считает, что все гены являются лишь вариантами или даже просто изомерами нуклеиновой кислоты. Я никак не могу с этим согласиться, так как молекулярная структура нуклеиновой кислоты слишком проста и однородна. Ведь это, прежде всего, не белковая молекула. У всех животных и растений нуклеиновая кислота одинакова или почти одинакова: думать о миллионах изомеров этой молекулы не приходится. Я считаю поэтому, что нуклеиновая кислота никакого отношения к генам не имеет”.
Поясним, что цитология — это область биологии, изучающая строение клетки. А мысль о том, что все гены являются изомерами нуклеиновой кислоты, не только красива, но и, как мы сейчас понимаем, по сути абсолютно верна. В данном случае Кольцов попал пальцем в небо из-за того, что слишком доверился биохимикам. А последние как раз в те годы стали почему-то склоняться к мнению, что ДНК — вообще не полимер, а совсем небольшая молекула, состоящая всего-то из четырех нуклеотидов (те самые “А”, “Т”, “Г” и “Ц”). Просто в ядре таких молекул очень много, вот и получается большая масса. Сам Кольцов не был биохимиком, поэтому не мог оценить, насколько эти данные надежны или ненадежны. Если бы он отважился допустить, что его открытая “на кончике пера” молекула наследственности не состоящий из тысяч аминокислот белок, а состоящая из тысяч нуклеотидов ДНК, это могло бы изменить всю мировую биологию. Но, насколько можно судить, Кольцову такое в голову не пришло. И никому другому в 1930-е годы — тем более.
Полемика Кольцова и Демерца хорошо показывает, сколь непрямыми путями обычно идет научная мысль. Две идеи, каждая из которых в отдельности была совершенно верна, столкнулись и разошлись. Слияния не произошло. Хотя ждать его оставалось уже совсем недолго.
Четвертый этап: эксперименты
В 1944 году американский биолог Освальд Эвери экспериментально показал, что ДНК, и только ДНК может вызвать трансформацию одной разновидности бактерий в другую — в данном случае безопасного штамма пневмококка в болезнетворный, вызывающий инфекционное воспаление легких. Вот этот момент и следует считать открытием генетической роли ДНК (см. рис. 8.2).
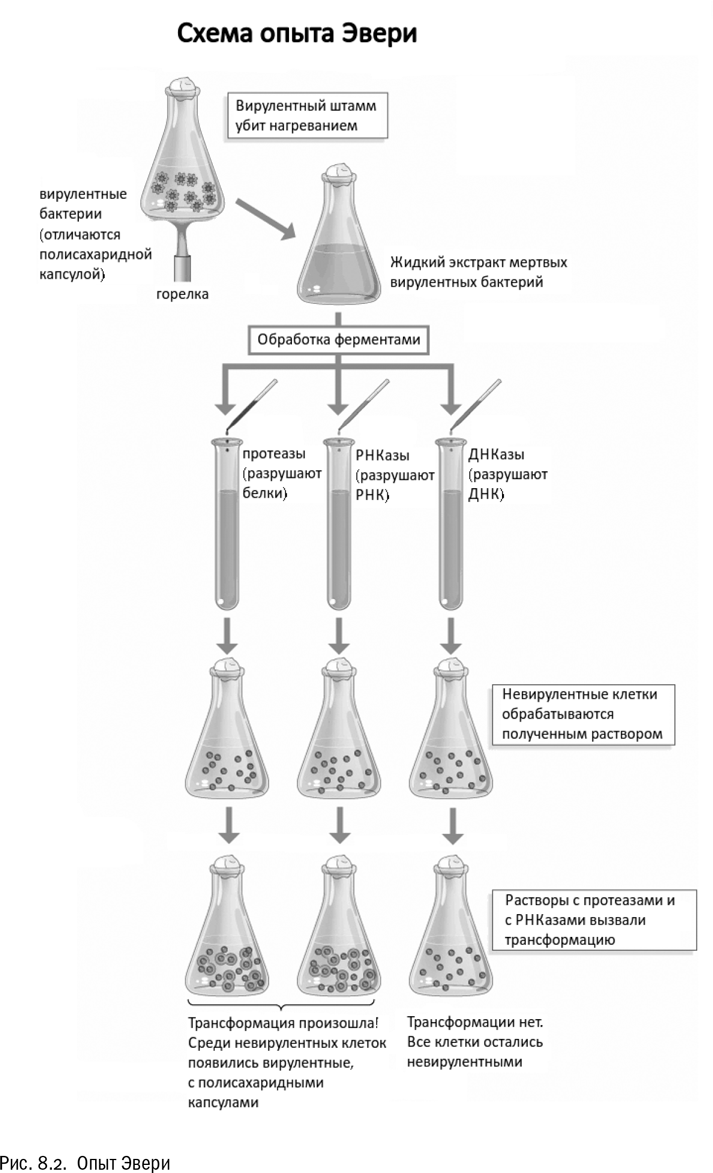
Освальд Эвери на протяжении всей своей научной карьеры, больше 40 лет, занимался только пневмококками — бактериями, которые вызывают пневмонию. Тут надо сказать, что ему невероятно повезло с объектом. Сам факт, что некая таинственная “растворимая субстанция” может передаваться от одной бактерии к другой и навсегда менять наследственные свойства бактерии-реципиента, был открыт именно на пневмококках еще в 1920-х годах. Но вот ответ на вопрос, что это за субстанция такая, требовал очень тонких, технически сложных биохимических опытов, поставить которые долгое время никто не мог. Эвери стал первым. Он многие годы работал над этой темой, продолжил работу несмотря на то, что уже вышел в официальную отставку по возрасту, и добился-таки успеха (в момент выхода главной статьи ему было 67 лет).
На самом деле поворотная точка была пройдена в марте 1943 года, когда Эвери прочитал перед попечителями Рокфеллеровского фонда небольшой доклад, завершавшийся однозначным выводом: субстанция, способная вызывать стойкую трансформацию бактерий, — это ДНК. Задержка выхода статьи на год объясняется в основном тем, что с ее подачей медлили, обдумывая каждое слово. Эвери и его сотрудники прекрасно понимали, насколько важный результат они получили.
В том же 1944 году австралиец Фрэнк Макфарлейн Бёрнет опубликовал ключевой тезис: ДНК ведет себя так, как будто гены именно из нее и состоят. После работы Эвери этот вывод вроде бы напрашивался, так что выяснение приоритета тут выглядит похожим на знаменитый спор о том, кто первым сказал “э”. Но на самом деле все было не так просто. Очень многие крупные биологи и после 1944 года все еще по инерции держались за убеждение, что гены состоят из белка, а ДНК — в лучшем случае всего лишь мутагенное вещество (правда, вызывающее какие-то уж очень странные предсказуемые мутации). Именно так поначалу решил, например, знаменитый генетик Феодосий Добржанский. Сам Эвери, насколько можно судить, колебался. И вообще, без споров не обошлось. Бёрнет — между прочим, выдающийся вирусолог и иммунолог, впоследствии совсем за другие заслуги получивший Нобелевскую премию, — оказался смелее большинства мэтров.
Так или иначе, теперь все убедились, что во всяком случае какое-то отношение к генетической информации ДНК имеет точно. Цель была определена, и охота началась.
Пятый этап: двойная спираль
Результаты Эвери (которые вскоре были подтверждены и в других лабораториях) оживили угасший было интерес биохимиков к нуклеиновым кислотам. Во второй половине 1940-х годов эта тема пережила своего рода возрождение. Эрвин Чаргафф, высококвалифицированный биохимик европейской школы, довольно быстро опроверг сбивавшую всех с толку старую “тетрануклеотидную теорию”, согласно которой ДНК должна состоять из одинаковых блоков по четыре нуклеотида. Как мы помним, эта теория исходила из того, что четыре азотистых основания присутствуют в ДНК в равных количествах. Чаргафф провел точные измерения, показавшие, что это неверно. На самом деле количество аденина строго равно количеству тимина, а количество гуанина строго равно количеству цитозина:
[А] = [Т][Г] = [Ц]
Эти соотношения называются правилами Чаргаффа. Отношение [А+Т]/[Г+Ц] константой как раз не является и может отличаться у разных организмов. А вот правила Чаргаффа соблюдаются всегда. Что же это может означать? Никаких четырехнуклеотидных блоков в составе ДНК нет, иначе соотношения совершенно точно были бы иными. Но почему бы там не быть не четверкам, а парам? Правила Чаргаффа легче всего объяснить, предположив, что нуклеотиды входят в ДНК в составе пар: или АТ, или ГЦ.
В этом месте полезно вспомнить, что, собственно говоря, означают буквы А, Т, Г и Ц. Аденин (А) и гуанин (Г) — это пуриновые азотистые основания, а цитозин (Ц) и тимин (Т) — пиримидиновые. И мы тут же получаем вытекающее из правил Чаргаффа важнейшее утверждение: в одной и той же молекуле ДНК общее количество пуриновых оснований (А и Г) всегда строго равно общему количеству пиримидиновых (Т и Ц). В этом плане предполагаемые пары АТ и ГЦ устроены одинаково: в каждой паре одно основание пуриновое, другое — пиримидиновое.
Оставалось сделать последний шаг: установить трехмерную структуру молекулы ДНК, решив задачу из области науки, которая называется стереохимия.
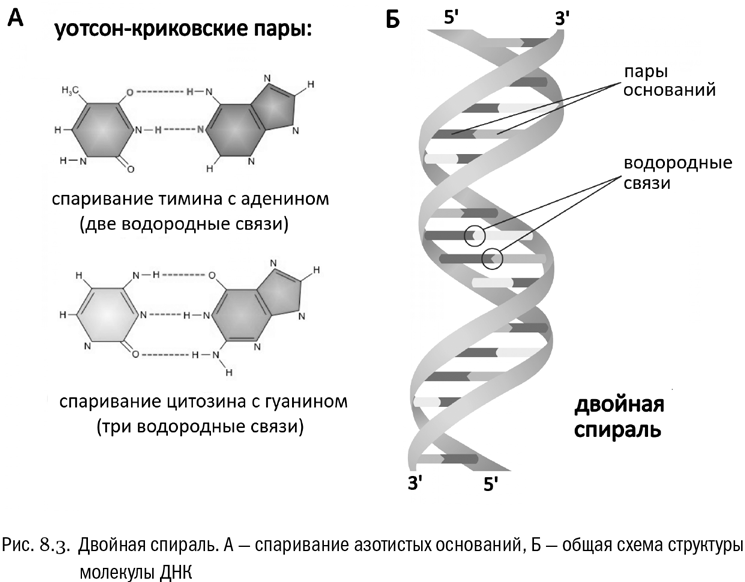
Пытаясь разобраться в устройстве ДНК, американец Джеймс Уотсон и англичанин Фрэнсис Крик увидели, что возникающие между азотистыми основаниями водородные связи делают пары аденин-тимин и гуанин-цитозин фактически идентичными друг другу по общей форме. И там и там возникает одна и та же легко узнаваемая структура из трех колец (см. рис. 8.3А).
У Уотсона и Крика получилось, что между аденином и тимином должны образоваться две водородные связи, а между гуанином и цитозином — три. Поэтому пара ГЦ должна быть прочнее, чем пара АТ (сейчас мы знаем, что это действительно так). Зато в обеих парах — один пурин и один пиримидин, как и ожидалось.
Надо заметить, что решить эту задачу Уотсону и Крику удалось не с первого раза. Помните, мы говорили, что у азотистых оснований бывают две изомерные формы — спиртовая и кетонная (см. главу 7)? Так вот, в учебниках биохимии 1940-х годов чаще всего изображалась только спиртовая форма. Уотсон и Крик исходили именно из нее, и ничего собрать у них не получалось, пока профессиональный химик Джерри Донохью не объяснил им, что это бесполезное занятие: в физиологических условиях азотистые основания всегда находятся не в спиртовой форме, а в кето-форме! Ни Уотсон, ни Крик химиками вообще-то не были, так что эта помощь оказалась драгоценной.
Но теперь у них получилась модель молекулы, идеально соответствовавшая правилам Чаргаффа. С самим Чаргаффом, кстати говоря, у Уотсона и Крика общение не сложилось — тот счел их дилетантами, с которыми и разговаривать-то серьезно не стоит. “Презрение Чаргаффа к нам достигло предела, когда Фрэнсис вынужден был признаться, что не помнит химических различий между четырьмя азотистыми основаниями”, — писал потом Уотсон. Но это было уже неважно.
Осталось сопоставить полученную модель с данными рентгеноструктурного анализа, дававшего размытые и нечеткие, но все-таки снимки молекул ДНК в рентгеновских лучах. К счастью, по соседству с Уотсоном и Криком работали сильные рентгеноструктурщики — Морис Уилкинс и Розалинд Франклин. Забегая вперед, надо сказать, что через несколько лет Уилкинс получил вместе с Уотсоном и Криком Нобелевскую премию, а Франклин, к сожалению, нет, потому что безвременно умерла от рака.
В итоге Уотсон и Крик собрали (в самом буквальном смысле, из проволоки и жести) модель молекулы ДНК, полностью отвечавшую как биохимическим, так и рентгеноструктурным данным (см. рис. 8.3Б). Оказалось, что молекула состоит из двух цепочек. И против любого основания в одной цепочке должно стоять дополнительное к нему (комплементарное) основание в другой: против аденина — тимин, против тимина — аденин, против гуанина — цитозин и против цитозина — гуанин. Причем эти цепочки должны быть антипараллельны, то есть противоположно направлены. Вот здесь нам как раз пригодится усвоенное в начале этой главы представление о 5'-конце и 3'-конце цепочки нуклеиновой кислоты. У двух цепочек, образующих молекулу ДНК, эти концы всегда направлены в противоположные стороны. С параллельными цепочками модель не собиралась, антипараллельность неожиданно оказалась совершенно обязательным условием. Никаких ковалентных связей между двумя цепочками ДНК нет — только водородные. Так что формально это две отдельные молекулы, их просто традиционно принято считать за одну.
В трехмерном пространстве изученная Уотсоном и Криком молекула ДНК имеет форму двойной спирали, напоминающую двойной штопор (такие редко, но встречаются). Если ее раскрутить, то к каждой из двух цепочек можно будет автоматически достроить комплементарную — конечно, при наличии в окружающем растворе нужных мономеров-нуклеотидов. Стереохимия азотистых оснований просто не допустит другого варианта: к аденину может пристроиться только тимин, к гуанину — только цитозин и так далее. А это означает, что Уотсон и Крик открыли не более и не менее как механизм копирования наследственной информации.
В свете этого осознания авторский комментарий, сделанный Уотсоном и Криком в их знаменитой статье 1953 года, воистину выглядит шедевром скромности. Там сказано:
“It has not escaped our notice that the specific pairing we have postulated immediately suggests possible copying mechanism for the genetic material”.
(В переводе: “От нашего внимания не ускользнуло то, что постулированное нами специфическое спаривание немедленно предполагает возможный механизм копирования генетического материала”.)
По существу, задача была решена в тот момент, когда удалось установить одинаковую форму пар АТ и ГЦ. Именно в этот день Фрэнсис Крик принялся рассказывать всем подряд в кембриджском пабе “Орел”, что они с коллегой только что раскрыли секрет жизни (отчего даже Уотсону, никогда не страдавшему избытком скромности, стало не по себе). Но и в самом деле, собрать правильную трехмерную модель ДНК после этого оказалось делом техники.
Вся эта история отлично показывает, насколько коллективным процессом является наука. Уотсон и Крик при всем их огромном таланте были еще и необычайно удачливы: в их распоряжении оказалось готовое полное условие задачи, которую оставалось только решить. Например, их модель двойной спирали была бы невозможна без результатов биохимического анализа Чаргаффа и равным образом — без рентгенограмм Уилкинса и Франклин. Причем получить все это самостоятельно Уотсон и Крик не могли — у них просто не было такой профессиональной подготовки. А Чаргафф, в свою очередь, работал на основе старой “тетрануклеотидной теории”, хоть и опроверг ее. Двигаясь таким образом вспять по цепочке ученых, каждый из которых делал возможной работу следующего, мы получим некий причудливый граф, где Уотсон и Крик будут располагаться в одной из вершин. Впрочем, о чем-то подобном писал еще в XII веке Бернар Шартрский: “Мы подобны карликам, усевшимся на плечах великанов; мы видим больше и дальше, чем они, не потому, что обладаем лучшим зрением, и не потому, что выше их, но потому, что они нас подняли и увеличили наш рост собственным величием”.
Самое главное в двойной спирали ДНК не то, что она спираль, а то, что она двойная. Теоретически двойная цепь ДНК вполне могла бы быть и линейной, как застежка-молния, без всякой спиральности. На ее свойства как носителя информации это нисколько не повлияло бы. Спиральная конформация энергетически выгоднее по чисто химическим причинам, из-за углов между ковалентными связями. Но тут химия неожиданно совпала с эстетикой: двойная спираль просто красива. Неудивительно, что она быстро стала широко известным символом глубинного механизма жизни.
И снова две кислоты
Теперь самое время прояснить вопрос: в чем же смысл химических отличий между ДНК и РНК?
Прежде всего напомним, что главное из этих отличий выражено в самом названии: в состав РНК входит рибоза, а в состав ДНК — дезоксирибоза. Добавив к этому факту кое-что еще, мы получим следующую картину:
* РНК гораздо легче синтезируется, потому что реакция синтеза дезоксирибозы требует более сложных ферментных систем, чем реакция синтеза рибозы;* ликвидация 2'-гидроксильной группы (той самой, которой дезоксирибоза лишена) резко уменьшает химическую активность нуклеиновой кислоты, и в том числе ее подверженность спонтанному распаду с участием воды;* ДНК редко бывает одноцепочечной (только у некоторых вирусов), а в двуцепочечной форме она очень устойчива из-за огромного количества водородных связей и — дополнительно — из-за гидрофобного взаимодействия между множеством налегающих друг на друга плоских колец азотистых оснований.
Создается явное впечатление, что ДНК была “специально придумана” природой как долговременное надежное хранилище информации.
Есть и еще одно важное отличие. В 1980-х годах было обнаружено, что некоторые РНК способны вести себя как ферменты, катализируя определенные химические реакции (например, разрезание или синтез других РНК). Такие РНКовые аналоги ферментов называются рибозимами. Эта способность вовсе не является редкостью, она не требует большой длины РНК или какой-то уникальной нуклеотидной последовательности. Рибозимы часто бывают длиной в 100–200 нуклеотидов или меньше. Самый маленький известный рибозим имел длину всего 13 нуклеотидов. Это означает, что химическая эволюция могла создать рибозимы очень легко. Даже при совершенно случайном переборе последовательностей РНК какие-нибудь рибозимы обязательно появятся, причем довольно быстро.
У ДНК же каталитическая способность резко ослаблена все той же потерей 2'-гидроксильной группы. Таким образом предотвращается, например, случайное возникновение саморазрезающихся рибозимов — в РНК бывает даже такое. Живым организмам совершенно не нужна каталитическая способность ДНК. Наоборот, чем она инертнее, тем лучше. Для редактирования генетической информации есть другие молекулы. А ДНК должна ее просто хранить.
Открытие рибозимов сразу вызвало к жизни так называемую гипотезу РНК-мира. Сторонники этой гипотезы считают, что первоначально живые системы состояли в основном из разнообразных молекул РНК. Эти молекулы РНК служили и носителями информации, и катализаторами всевозможных химических реакций. И только потом специализированными катализаторами стали белки, а специализированными хранителями информации — молекулы ДНК. За самой же РНК остались функции, связанные с передачей генетической информации и частично с регуляцией работы генов.
В самом деле, мы скоро убедимся, что ДНК, в отличие от РНК, не является в системе передачи генетической информации абсолютно необходимым звеном. Все генетические процессы в принципе можно было бы осуществить с помощью одних только РНК, которые и в современном мире довольно разнообразны. Генетическую систему, включающую только РНК, сконструировать можно, а вот включающую только ДНК — нельзя.
К этому можно добавить, что соединения рибозы и сами по себе, независимо от нуклеиновых кислот, играют важную роль в обмене (например, в энергетическом), чего никак нельзя сказать о соединениях дезоксирибозы: последняя нужна для синтеза ДНК, и больше ни для чего. Все это показывает, что ДНК — продукт гораздо более глубокой специализации, чем РНК, и в целом подкрепляет гипотезу РНК-мира.
Теперь обратимся еще к одному отличию, уже нам известному: в состав ДНК вместо урацила почему-то входит тимин. Напомним, что тимин отличается от урацила всего лишь одной дополнительной метильной группой (–CH3). А дело тут, скорее всего, вот в чем. Есть еще одно азотистое основание, которое входит и в РНК, и в ДНК, и при этом является химически довольно неустойчивым. Это цитозин. Он легко подвергается спонтанному дезаминированию — потере аминогруппы, на место которой приходит гидроксильная группа. А цитозин с гидроксилом вместо аминогруппы — это не что иное, как урацил. Такие уж у этих веществ формулы. Получается, что если хранить генетическую информацию на РНК, то она неминуемо будет постепенно “засоряться” урацилом, образующимся из-за спонтанного дезаминирования цитозина.
А вот из ДНК урацил исключен вообще. Всеобщая замена урацила на тимин дает возможность легко исправлять ошибку дезаминирования, “настроив” соответствующие ферменты на вырезание любых нуклеотидов с урацилом как заведомо ошибочных. Но такая замена имеет смысл только в том случае, если ДНК уже используется как носитель информации! И получается сильный чисто химический довод за то, что ДНК была выбрана живыми системами на роль долговременного хранилища генетической информации “специально”. Никакой другой функции у нее никогда и не было.
Остается понять, какие живые организмы и когда создали это новшество. Это — интереснейший и совершенно нерешенный вопрос. Есть, например, гипотеза, что ДНК как таковая была “изобретена” ДНК-содержащими вирусами, которые потом инфицировали древнейшие клетки, еще содержавшие только РНК, и передали им свою “информационную технологию”. Если это верно — значит, современная клетка, содержащая ДНК, возникла в результате взаимодействия (или даже слияния) древней РНК-содержащей клетки с крупным ДНК-содержащим вирусом. Правда, тут возможна и более сложная последовательность событий — о том, какая именно, мы поговорим в главах 12 и 14.
Назад: 7. нуклеотиды
Дальше: ЧАСТЬ II МЕХАНИЗМ ЖИЗНИ

