Книга: Динозавры. 150 000 000 лет господства на Земле
Назад: Завроподы и их родственники: завроподоморфы
Дальше: 3. Анатомия
Птицетазовые динозавры: бронированные, утконосые, рогатые и их родственники
И последняя, третья большая группа динозавров – птицетазовые. Это клада в основном травоядных динозавров, в которую входят бронированные анкилозавры, пластинчатые стегозавры, гребненосные утконосые гадрозавры и рогатые цератопсы. Наиболее известные птицетазовые – крупные четвероногие животные размером обычно с носорога или слона. Хотя было немало и небольших двуногих видов. Например, самые ранние птицетазовые были маленькими, двуногими всеядными животными с хватательными передними конечностями и напоминали ранних представителей теропод и завроподоморфов.
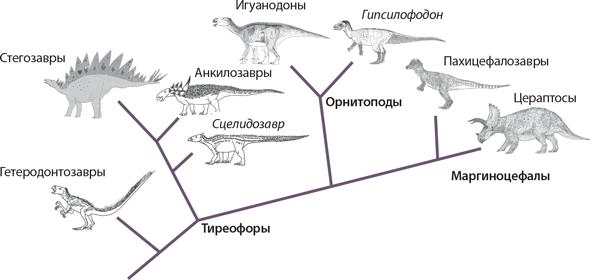
Упрощенная кладограмма птицетазовых показывает, что к ним относятся три основные клады: бронированные тиреофоры, довольно обычные орнитоподы и рогатые маргиноцефалы
Несколько ключевых признаков позволяют отличить даже самых ранних птицетазовых динозавров от представителей других групп. Самый очевидный – лишенная зубов кость на конце нижней челюсти, которая называется «предзубная кость». В жизни эту кость покрывал роговой чехол. Таким же чехлом был покрыт кончик верхней челюсти, и вместе они образовывали режущую кромку, которая помогала срывать листья, ветки и другие части растений. Мы знаем, что челюсти многих птицетазовых заканчивались клювом, потому что эти клювы сохранились у некоторых окаменелостей. Конечно, птицетазовые полагались не только на свои клювы. У них были и зубы, которыми они измельчали и перетирали пищу, а у некоторых (в первую очередь у гадрозавров и цератопсов) появились зубные батареи, состоящие из сотен плотно скомпонованных зубов. Мы подробнее рассмотрим зубные батареи и принцип их действия в главе 4.
Другая ключевая особенность относится к тазовому поясу. Обычно у рептилий лобковая кость направлена вниз и вперед, но у птицетазовых она направлена вниз и назад. Зачем появилась такая необычная конфигурация? Обычно это объясняют так: направленная назад лобковая кость позволила кишечнику увеличиться в размерах и занять больше места в брюшной полости – полезная особенность для животных, которые питались растительной пищей. Но с этим объяснением есть проблемы. Например, у других травоядных динозавров (таких как завроподоморфы) лобковая кость не повернута назад, хотя им это, конечно, пригодилось бы. Другое возражение: у манирапторовых теропод лобковые кости тоже обращены назад, хотя они почти наверняка не так полагались на растительную пищу, как ранние птицетазовые. Этот аспект эволюции птицетазовых пока что остается загадочным и требует дополнительного изучения.
Самые ранние птицетазовые
Самые ранние птицетазовые – лакинтасаура из Венесуэлы, а также эокурсор, гетеродонтозавр и лесотозавр из Южной Африки. Большинство из них относятся к ранней юре. Все они родом с южных континентов – еще одно свидетельство того, что ранняя эволюция динозавров в основном происходила в Гондване. По-видимому, они не были исключительно травоядными, как более поздние птицетазовые, и ели насекомых и других мелких животных, а также отдельные части растений. Кроме этого, об их образе жизни и поведении мало что известно. Особый интерес вызывает лакинтасаура, потому что множество особей этого рода погибло в одном и том же месте. Это может означать, что они были социальными, стадными животными, как более поздние и продвинутые птицетазовые в поздней юре и мелу. Возможно, птицетазовые динозавры были социальными с самого начала своей эволюции.

Один из древнейших известных птицетазовых – венесуэльская лакинтасаура. Она была небольшой, менее 1 м в длину, что типично для ранних представителей всех групп динозавров. Зубы и форма тела предполагали некоторую степень всеядности
Примечательная группа под названием гетеродонтозавриды может быть одной из самых ранних птицетазовых. На это указывают их длинные хватательные передние конечности, как у теропод, и отсутствие многих продвинутых признаков в строении зубов и челюстей, которые есть у большинства других птицетазовых. Но читайте дальше.
Название «гетеродонтозавриды» означает «разнозубые ящеры» и связано с тем, что у них имеются разные зубы: похожие на резцы, на клыки и на коренные зубы (моляры). Щечные зубы – обычное дело для травоядных животных, а вот клыки и хватательные передние лапы навели некоторых экспертов на мысль, что гетеродонтозавриды могли охотиться на мелких животных. Изначально их обнаружили в отложениях, относящихся к ранней юре, но теперь мы знаем, что они жили и гораздо позже, например в поздней юре в Китае и в раннем мелу в Англии. Китайский гетеродонтозаврид тяньюйлун особенно важен, потому что у него сохранились длинные нитевидные образования на теле и хвосте. Гетеродонтозавриды также интересны тем, что по большей части были мелкими. Позднеюрский фрутаденс, найденный в штате Колорадо, США, не превышал 75 см в длину даже во взрослом состоянии, что делает его одним из самых маленьких птицетазовых, найденных на сегодняшний день.

Череп гетеродонтозавра, найденный в Южной Африке в начале 1960-х гг., вызвал сенсацию. Хорошо виден костный стержень, частично закрывающий глазницу. При жизни он был соединен мягкими тканями с вершиной черепа и нависал над глазным яблоком
Как уже говорилось, большинство экспертов по динозаврам согласны, что гетеродонтозавриды были примитивными, «архаичными» птицетазовыми, которые появились на ранних этапах истории группы. Но у них есть особенности, которые встречаются у рогатых и купологоловых динозавров, в том числе крупные, похожие на клыки зубы в верхней челюсти. Таким образом, вероятно, что гетеродонтозавриды на самом деле являются ранними представителями рогатых и костеголовых динозавров, а не просто «архаичными» птицетазовыми.
Большинство птицетазовых относится к одной из трех или четырех основных клад с разной формой тела и образом жизни. Первая из них – тиреофоры. Это бронированные животные, самые известные из которых – стегозавры (пластинчатые динозавры) и анкилозавры (бронированные динозавры). У обеих групп были ряды костных пластин, называемых остеодермами, вдоль верхней части шеи, спины и хвоста. Самый известный из ранних тиреофоров – скутеллозавр из ранней юры США. Он был чуть более 1 м в длину, а его анатомия и пропорции конечностей говорят, что он передвигался на двух ногах. Его остеодермы располагались несколькими параллельными рядами и были простыми по форме. По сути, это были просто скругленные костяные пластинки с приподнятым центральным гребнем.

Раннеюрский сцелидозавр из Англии известен по нескольким хорошо сохранившимся экземплярам. Тот, что на фото, был найден первым в 1858 г. Это почти полный сочлененный скелет, частично заключенный в ряд блоков аргиллита
Близким родственником скутеллозавра был более крупный и тяжелый сцелидозавр из ранней юры Англии. Пропорции его конечностей говорят, что он уже был четвероногим, а его броня была больше и сложнее, чем у скутеллозавра. Что интересно, у одних сцелидозавров было гораздо больше брони, чем у других. У некоторых были рога, шипы на конечностях и участки брони на теле, которые совершенно отсутствовали у других особей. Возможно, количество брони варьировалось между полами или у старых особей брони было больше, чем у молодых.
Анкилозавры: ходячие крепости
Со временем тиреофоры, похожие на сцелидозавра, породили более крупных животных с мощными конечностями и все более обширной и сложной броней. В конечном итоге от них произошли анкилозавры, которые впервые появились в палеонтологической летописи в средней юре и сохранились вплоть до конца мелового периода. Обсуждения истории анкилозавров обычно выливаются в разговор об их удивительно разнообразной броне. Действительно, большую часть шеи, спины и хвоста многих анкилозавров покрывали прямоугольные или овальные остеодермы. У некоторых видов боковая часть тела и хвоста была покрыта треугольными пластинами, а у других на спине торчали большие шипы или конусы. Вдобавок шею одних видов защищали костяные «воротники», у других были огромные костяные «булавы» на хвостах, а третьи щеголяли острыми шипами на передних и задних ногах.

Кончик хвоста некоторых анкилозаврид был жестким и оканчивался массивной «булавой», состоящей из округлых костяных глыб. Эта «булава» шириной более 60 см принадлежит гигантскому североамериканскому анодонтозавру
Анкилозавры – одни из самых необычных динозавров в смысле анатомии. Отверстия черепа, которые, как правило, очевидны у других динозавров, в этом случае были закрыты вновь образованной костью, а верхнюю поверхность черепа покрывали толстые прочные костяные наросты. Часто встречалось необычное расположение ноздрей (иногда в передней части морды, а не по бокам), а внутренняя анатомия носа была удивительно сложна. Таз у анкилозавров тоже был очень странный – очень широкий, образованный в основном большими и плоскими сверху, как полки, подвздошными костями. Тазобедренный сустав динозавров обычно характерен сквозным отверстием. Но у анкилозавров это была чашеобразная впадина, обычно полностью закрытая костью с внутренней стороны. Также у анкилозавров часто срастались вместе позвонки, по-видимому, для большей жесткости тела и поддержки веса брони. Наконец, кости конечностей у анкилозавров были короткие и прочные.

Этот североамериканский анкилозаврид сколозавр сохранился на боку (на фото он снят со стороны брони). Туловище было широкое, а спина почти плоская
Различают несколько групп анкилозавров, но по-прежнему ведутся споры о том, кто кому родственник. Многие известные анкилозавры входят в кладу анкилозавриды. У них были короткие, широкие черепа и треугольные рога на задней части головы. Сюда относятся знаменитые динозавры с «булавами» на хвосте, анкилозавр и эуплоцефал из позднего мела Северной Америки. Вторая группа называется нодозавриды. У ее представителей более длинные и узкие черепа, которые выглядят попроще, хотя у них часто бывают округлые костяные наросты над и позади глаз. У некоторых нодозаврид были длинные мощные шипы в области шеи и плечей. У одних видов эти шипы были направлены наружу и назад, у других – например, у эдмонтонии (Edmontonia rugosidens) из позднего мела Северной Америки – наружу и вперед, а иногда разветвлялись на конце.

Тело анкилозавров было покрыто несколькими видами брони. Остеодермы на фото принадлежат европейскому меловому анкилозавру полакантусу. Мелкие, чешуеподобные образования покрывали часть спины животного, а более крупные пластины торчали вверх и в стороны из тела и хвоста
Некоторые специалисты выделяют еще одну группу раннемеловых анкилозавров из Западной Европы и США, которая называется полакантиды. Самый известный их представитель – полакантус из Англии и Испании. Щитовидная пластина закрывала у него область таза, а череп сочетал признаки анкилозаврид и нодозаврид. По-прежнему неясно, к кому ближе полакантиды – к нодозавридам или анкилозавридам и вообще стоит ли выделять их в отдельную группу. Они могут оказаться ранними представителями одной из основных групп анкилозавров.
Стегозавры: пластины и шипы
Самый известный представитель стегозавров так и называется – стегозавр. Он жил в позднем юрском периоде в США, Португалии и, возможно, Китае. Знаменит он своими ромбовидными пластинами вдоль спины. Сочлененные скелеты показывают, что пластины были расположены асимметрично, в шахматном порядке – странная конфигурация, которая резко контрастирует с обычными симметричными структурами у других стегозавров и вообще тиреофоров. Почему и как появилась такое асимметричное расположение, до сих пор сложно сказать наверняка, особенно если учесть, что оно, видимо, было вызвано каким-то загадочным генетическим событием.
Помимо пластин у стегозавра был длинный низкий череп, а узкая пасть заканчивалась клювом. У него были крепкие, мускулистые передние конечности, длинные задние и хвост с двумя парами длинных конических шипов. Изучение гибкости хвоста стегозавра и мышечной силы показало, как он мог использовать эти шипы. Мы еще вернемся к этому вопросу в главе 3.

Череп стегозавра – самого знаменитого и изученного из всей группы – длинный и тонкий, почти трубчатый по форме. Заметим, что предглазничное окно сильно уменьшено, почти закрыто
На самом деле стегозавр – не обычный представитель своей группы. Во-первых, он намного крупнее большинства других стегозавров: достигал 9 м в длину, тогда как другие были в основном от 4 до 7 м. Во-вторых, его пластины располагались асимметрично, в отличие от симметричных и гораздо меньших пластин и шипов других представителей группы. Не все стегозавры были одинаково шипасты. У стегозавраи его близких родственников шипы были только на конце хвоста, а у других видов шли вдоль всего хвоста и части спины. Классический пример – кентрозавр из поздней юры Танзании. В-третьих, у стегозавра не было плечевых шипов. Если бы не стегозавр, их можно было бы считать типичной особенностью стегозавров: у многих видов на плечах торчали длинные шипы, иногда широкие и округлые у основания. У китайского гигантспинозавра огромные плечевые шипы выгибались назад и наружу, причем каждый шип по длине был сравним со всей грудной клеткой животного.
Орнитоподы: игуанодон, утконосые динозавры и их родственники
Вторая крупная группа птицетазовых впечатляет куда меньше. Она называется орнитоподы. Они обитали и по всему миру от середины юрского до конца мелового периода и занимали нишу мелко- и среднеразмерных травоядных в многочисленных средах обитания. Самые крупные орнитоподы – гадрозавры (например, шаньдунозавр из позднего мела Китая) – были сравнимы по размеру с завроподами и достигали 15 м в длину, а весили, возможно, до 13 т. Челюсти и челюстные суставы орнитопод не похожи на челюсти других птицетазовых. Со временем количество зубов увеличилось (у некоторых видов было более тысячи зубов), а само строение зубов усложнилось.
В «ядре» группы находятся: небольшой двуногий гипсилофодон из раннего мела Англии, огромный раннемеловой игуанодон из Европы, а также множество «утконосых» динозавров (они же гадрозавры) с гребнями или без них. Гадрозавры произошли в раннем мелу от игуанодонподобных животных, скорее всего в Азии. Оттуда они распространились по всем континентам, кроме Африки и Австралазии. Игуанодон, гадрозавры и множество их родственников отличаются от маленьких гипсилофодоновналичием U-образной (а не V-образной) предзубной кости и более глубокой нижней челюстью. Эти игуанодоноподобные орнитоподы включены в кладу под названием игу анодонты.
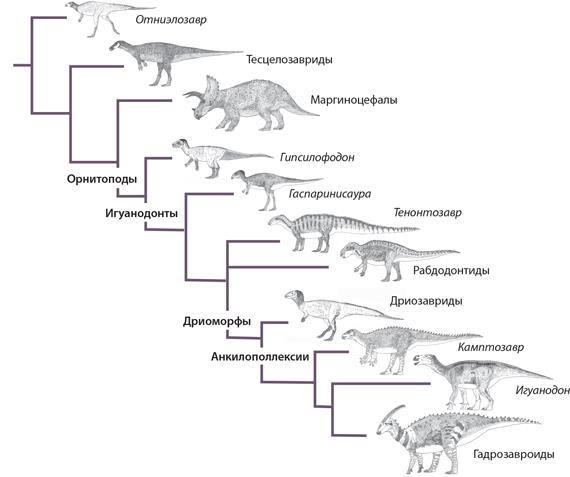
Большую часть своей истории орнитоподы были небольшими резвыми двуногими длиной менее 3 м. Однако в юрском периоде возникла новая клада под названием игуанодонты. В ней появились гигантские четвероногие формы, такие как игуанодон и гадрозавры. Недавние исследования показывают, что несколько групп птицетазовых, ранее считавшихся орнитоподами, в том числе и тесцелозавриды, на самом деле находятся вне группы, в которую входят маргиноцефалы и орнитоподы
На протяжении большей части XX в. считалось, что практически все мелкие двуногие птицетазовые были близкими родственниками гипсилофодона, и поэтому всех их объединили в семейство гипсилофодонтиды. Предполагалось, что в это семейство входили архаичные птицетазовые ранней юры (например, лесотозавр), а также крупные игуанодонподобные животные вроде тенонтозавра из раннего мела США. Но явных анатомических признаков, которые действительно могли бы продемонстрировать тесную связь между этими животными, так и не нашлось. Похоже, что некоторые из рассматриваемых орнитопод вообще не родственники гипсилофодону, а скорее, близки к игуанодонтам или даже входят в их кладу. Крепко сложенные рабдодонтиды из позднего мела Европы тоже состоят в этой кладе, как и дриозавриды – юрская и раннемеловая группа, отличающаяся длинными, стройными ногами, короткими передними лапами и короткими мордами. Дриозавриды найдены в Англии, Танзании, Нигере, США и других странах. Несколько групп, некогда отнесенных к гипсилофодонтидам, теперь, по-видимому, находятся вне клады, в которую входят гипсилофодон и игуанодонты. Может оказаться, что они вообще не орнитоподы. Это, например, североамериканские зефирозавры, тесцелозавры и ородромины, а также китайские джехолозавры. Тем не менее общее строение тела у всех этих животных похоже – все они мелкие двуногие травоядные. Рабдодонтиды, возможно, выделялись, поскольку странные пропорции их конечностей и бедер намекают на передвижение на четырех ногах.

Череп игуанодонтов (на фото выше показан череп западноевропейского мантеллизавра) длинный и имеет обширный беззубый участок спереди. Их ключевая особенность – гигантский размер. Этот череп, например, 45 см в длину
Зубы игуанодона, мантеллизавра и похожих игуанодонтов (посередине) имеют большие ромбовидные коронки с выраженной режущей кромкой и грубо зазубренными краями. Со временем коронки стачивались, как это произошло с зубом вверху справа
Дриозавриды показывают, что игуанодон, гадрозавры и их родственники произошли от двуногих травоядных средних размеров, которые довольно хорошо бегали и имели относительно короткие передние лапы и кисти. Новые, более крупные игуанодонты в раннем мелу стали очень многочисленны и разнообразны и начали играть важную – возможно, самую важную – роль растительноядных животных в сообществах Европы, Азии и Северной Америки в течение мелового периода.
Гадрозавры – крупнейшая и самая важная группа из них. Они были оснащены зубными батареями, широким клювом, могли передвигаться как на двух, так и на четырех ногах и породили огромное разнообразие размеров тела и форм черепа. Часто подразумевается, что гадрозавры по существу все похожи и отличаются только формой головы. В конце концов, известны гадрозавры с длинными мордами и широкими, утиными клювами (эдмонтозавр), с глубокими, изогнутыми вниз мордами и большими носовыми полостями (майазавр), с твердыми шиповидными гребнями (зауролоф), с полыми плоскими гребнями (коритозавр) и с полыми трубковидными гребнями (паразауролоф). Зачем этим динозаврам было нужно столько видов гребней – интересный вопрос, который мы еще рассмотрим в главе 4.

Джехолозавр из раннего мела Китая – довольно обычный мелкий птицетазовый. У него сохранился примитивный признак: шесть зубов в передней части верхней челюсти. Поздние крупные орнитоподы уже утратили зубы в этой части черепа
Обобщение, что все гадрозавры «по сути одинаковы», неточно, потому что форма тела у многих линий сильно отличается. У брахилофозавров и грипозавров были длинные и тонкие передние конечности, у паразауролофин – короткие и мощные, а у ламбеозаврин был высокий костяной гребень на спине, который придавал им горбатый вид. Предположительно, эти различия были связаны с образом жизни и питанием.
Странноголовые динозавры: костяные головы и рога
У орнитопод есть несколько общих признаков еще с двумя группами: пахицефалозаврами (костеголовыми или купологоловыми динозаврами) и цератопсами (они же рогатые динозавры). Цератопсы известны благодаря славе своих гигантских четвероногих позднемеловых представителей, таких как трицератопс. У большинства были рога на носу и над глазами, а также воротник, выступающий вверх и назад от черепа. Пахицефалозавры совсем другие. Все они были двуногими, у многих из них верхушка черепа была толстая, часто куполообразная.

На фото – череп стегоцераса из позднего мела Канады. Хорошо видна костяная «полка» в задней части черепа, типичная для маргиноцефалов. Мелкие листовидные зубы подходят для измельчения растительности
Хотя самые известные пахицефалозавры и цератопсы сильно отличаются, с ранними видами все совсем не так. У обеих групп есть признаки, которых нет больше ни у кого. Самый очевидный – костяной «воротник», выступающий из задней части черепа. Именно из-за этого признака обе группы объединены в кладу маргиноцефалы, что означает «краеголовые». Как мы уже говорили, есть вероятность, что гетеродонтозавриды – близкие родственники этой группы.
Пахицефалозавры – одни из самых загадочных динозавров, так как большинство видов известно лишь по фрагментарным остаткам. Найдено всего один или два хороших скелета. По ним видно, что у пахицефалозавров были широкое туловище и таз, короткие передние конечности и длинный тонкий хвост. Хвост пахицефалозавров необычен, потому что помимо обычных позвонков в нем есть многочисленные изогнутые косточки. Это окостеневшие сухожилия по краям мышц, идущих вдоль хвоста. Такая особенность хорошо известна у некоторых рыб (например, костистых, к которым относятся сомы, лосось, треска и т.д.), но у наземных позвоночных ничего подобного нет. Зачем такое строение хвоста пахицефалозаврам – полнейшая загадка.

Удивительный куполообразный череп пахицефалозавра из позднего мела США сочетает толстый округлый купол с шишками, наростами и рожками. Все животное было около 4,5 м в длину
Более знакомая их черта – плоский или куполообразный череп, с краев и по бокам украшенный костяными наростами и рожками. У самых известных видов – лучшим примером является пахицефалозавр (Pachycephalosaurus wyomingensis) из позднего мела США – куполообразная кость имеет толщину до 40 см. У других – например, гомалоцефала (Homalocephale calathocercos) из позднего мела Монголии – верхняя часть черепа плоская.

Про тело пахицефалозавров известно немного. Насколько мы можем судить, все они имели короткие, тонкие передние конечности, были быстрыми и передвигались на двух ногах. Здесь изображены преноцефалы, которые водились в Монголии и США
Представления о том, как разные виды пахицефалозавров были связаны друг с другом, сейчас меняются. С 1970-х гг. считалось, что пахицефалозавры с плоскими и куполообразными головами – это разные группы, которые произошли от общего предка, а потом разделились. Впоследствии появилось мнение, что плоскоголовые виды более тесно связаны с купологоловыми, чем с другими плоскоголовыми. В таком случае группа «плоскоголовые пахицефалозавры» не является кладой. А совсем недавно некоторые эксперты высказали мысль, что плоскоголовые – это просто молодые купологоловые. Мы еще обсудим эту идею в главе 4.
Цератопсы известны в основном из мелового периода, но несколько окаменелостей показывают, что они существовали уже в поздней юре. Самые древние из них – это иньлун и хуаляньцератопс из Китая. Оба они были мелкими двуногими динозаврами длиной меньше 2 м. Как и у других ранних цератопсов, у иньлуна не было рогов и большого костяного воротника. Задняя часть черепа была глубокая и широкая с огромными отверстиями для мышц, закрывающих пасть, а на щеках и позади глаз были большие костяные наросты. Как у всех цератопсов (но не всех прочих птицетазовых), на кончике верхней челюсти иньлуна была особая кость, которая называется клювовой или ростральной. Возможно, она помогала укрепить или увеличить клюв, который явно был важной частью черепа цератопсов.
Практически все ранние цератопсы были мелкие – размером от кошки до овцы. Несколько двуногих птицетазовых, которых определили как возможных ранних цератопсов (например, альбалофозавр из раннего мела Японии), с виду не особенно отличаются от двуногих орнитопод. Явные ранние цератопсы, как правило, имели узкие клювы, костяные бугры или рога на щеках, а также короткий, похожий на полочку воротник. Некоторые были двуногими. Классический пример – пситтакозавр из раннего мела Китая, Монголии и Сибири. Другие были четвероногими, например протоцератопс из позднего мела Китая и Монголии. Практически все они из Восточной Азии, где водились повсеместно в течение всего мелового периода. До конца мелового периода они также обитали в западной части Северной Америки и в некоторых областях Европы.
Примерно 90 млн лет назад от предка, похожего на протоцератопса и жившего в Азии или Северной Америке, произошла линия более крупных животных с длинными рогами, увеличенным воротником и большим количеством зубов. Большинство их видов входят в кладу цератопсиды. Именно к ним относятся трицератопс, шипастый стиракозавр и пахиринозавр, знаменитый тем, что вместо рога у него на носу костяной бугор. Все цератопсиды были крупными – размером от носорога до слона, – и все были приспособлены к передвижению на четырех ногах. Цератопсиды обитали повсеместно во многих сообществах позднего мела западной части Северной Америки. В то время в регионе обитало более 30 различных видов.

Как у почти всех цератопсид, у хасмозавра были массивный костяной воротник, острый клюв и большой рог на носу. Края воротника были украшены симметричными шипами. Разные цератопсиды сильно отличались друг от друга размерами и формой воротника, а также расположением и количеством рогов и шипов
Поразительные рога, воротники, шипы и костяные наросты цератопсид почти наверняка играли роль предупредительных сигналов и оружия. Как ими пользовались и какое эволюционное давление привело к их появлению, остается предметом споров. Этот вопрос очень важен для того, чтобы мы лучше поняли поведение динозавров, и мы к нему еще вернемся в главе 4.
Назад: Завроподы и их родственники: завроподоморфы
Дальше: 3. Анатомия

