Книга: Динозавры. 150 000 000 лет господства на Земле
Назад: Тероподы: хищные динозавры и птицы
Дальше: Птицетазовые динозавры: бронированные, утконосые, рогатые и их родственники
Завроподы и их родственники: завроподоморфы
Диплодок, брахиозавр и их родственники принадлежат к крупной группе динозавров под названием завроподы. Все они были большими или очень большими травоядными с длинной шеей, которые ходили на четырех ногах. Их челюсти и зубы явно были приспособлены для срывания и поедания листьев, вай папоротников и других частей растений. Завроподы славятся не только своими невероятными размерами, но и фантастически длинными шеями. У некоторых видов шея была в четыре-пять раз длиннее тела, состояла из 15 позвонков и была более 10 м длиной. В целом сходятся во мнении, что такая шея служила инструментом для сбора пищи и давала этим динозаврам конкурентное преимущество перед другими видами, когда дело доходило до того, как далеко они могли дотянуться. Несмотря на это единство, по-прежнему ведутся споры насчет того, в каком именно положении они держали шею, насколько гибкой она была и как ее использовали.
Завроподы необычны не только размерами и длиной шеи. Они также научились по-новому поддерживать свой огромный вес при помощи уникальных модификаций костей передних и задних конечностей. У них появилась чрезвычайно сложная система заполненных воздухом мешков, карманов и камер по всему скелету. А еще они развили пищеварительную систему, которая позволила им извлекать энергию из огромного количества низкокалорийной растительной пищи. Ученые, изучающие завропод, рассматривали современных птиц, крокодилов и крупных млекопитающих, чтобы лучше понять биологию и анатомию вымерших гигантов, но проблема в том, что завроподы не очень похожи ни на одно современное животное. Мы еще вернемся к вопросам их биологии.

Продвинутых завропод, таких как диплодоки, ни с кем не спутаешь, стоит лишь взглянуть на их колоннообразные конечности и длиннющие шею и хвост. Но это еще не все: череп и передние конечности у них тоже очень необычные
Долгие годы мы не могли понять, как сформировалось строение тела завроподовых. Давно известны динозавры, явно не относящиеся к завроподам, но куда более тесно связанные с ними, чем с другими динозаврами. У них тоже длинная шея, и они часто бывают очень большими, но при этом они в основном двуноги, их зубы и челюсти соответствуют всеядному образу жизни, а конечности не приспособлены выдерживать большой вес. Самые известные из этих родичей завропод – платеозавр из позднего триаса Германии, Швейцарии и других стран Европы, а также массоспондил из ранней юры Южной Африки, Лесото и Зимбабве. Вместе с завроподами они образуют кладу завроподоморфов, которая возникла в триасе от того же самого ящеротазового предка, от которого произошли и другие группы динозавров. В течение большей части XX в. завроподоморфов вроде платеозавра и массоспондила называли прозавроподами. Теперь это название вышло из употребления, поскольку оказалось, что прозавроподы – это не клада, а просто собрание разных линий, из которых одни тесно связаны с завроподами, а другие – нет.
Самые древние завроподоморфы сильно отличались от завропод, да и вообще от всех видов крупных завроподоморфов. Это были маленькие (всего 1,5 м длиной) легкие двуногие животные с зубами-лезвиями и большими крючковатыми когтями на длинных тонких пальцах. Недавно открытый пример – буриолестес из позднего триаса Бразилии. По сравнению с поздними завроподоморфами он неспециализирован, и трудно сказать, какое место он и ему подобные занимают на семейном древе динозавров. Буриолестес и похожие на него животные – в том числе панфагия и пампадромей – видимо, были хищниками, которые ели еще и листья, хотя неизвестно, какую пищу они употребляли чаще – животную или растительную. Точно сказать нельзя, пока не будет найден экземпляр с сохранившимся содержимым желудка или какое-нибудь другое прямое доказательство.
В триасовом периоде от подобных животных произошли более крупные длинношеие завроподоморфы, которые все больше и больше сосредотачивались на поедании растений. Сатурналия из позднего триаса Бразилии похожа на растянутую версию буриолестеса с маленькой головой. Ее шея и туловище длиннее, а череп меньше по отношению ко всей длине животного, чем у других ранних динозавров (сатурналия была всего метра два в длину). Новая форма тела с длинной шеей и маленькой головой оказалась крайне удачной, и двуногие, в основном травоядные, завроподоморфы заселили большинство континентов в остальной части триаса и большей части ранней юры.
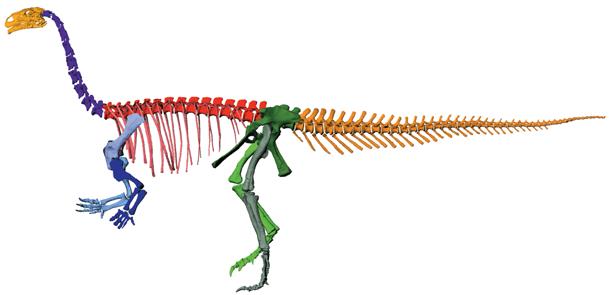
Платеозавр из позднего триаса Европы известен по многим скелетам, в том числе почти полным. Благодаря компьютерному моделированию специалисты выяснили, как он двигался и каков был его диапазон возможных поз
До недавнего времени ранних завроподоморфов вроде платеозавра или массоспондила часто изображали способными ходить и бегать на четвереньках, поставив ладонь на землю. Изучение костей и суставов, а также цифровое моделирование скелетов показали, что такое положение тела было невозможно. Передние конечности этих динозавров были зафиксированы в положении «ладонями внутрь», то есть они не могли развернуть передние лапы и запястья ладонями вниз. Значит, они не могли опираться ладонями о землю и должны были ходить только на задних ногах. Такое положение ладоней также характерно для теропод и птиц.
Есть и другие доказательства того, что платеозавр и его родственники передвигались на двух, а не на четырех ногах. Например, их передние конечности больше подходят не для ходьбы, а для защиты или хватания пищи. Да и пропорции в целом больше соответствуют двуногому передвижению – передние конечности настолько короче задних, что поза бега на четырех ногах, предложенная некоторыми экспертами, выглядит крайне маловероятной.
В какой-то момент завроподоморфы превратились из двуногих животных с короткими передними лапами в четвероногих, с длинными. Мы знаем, что это произошло примерно 207 млн лет назад, в конце триаса, так как у нас есть окаменелости больших четвероногих завроподоморфов из этого периода. Среди них меланорозавр и антетонитр из Южной Африки и лессемзавр из Аргентины. Эти гигантские завроподоморфы были не меньше по размеру, чем многие другие виды, которые появились гораздо позже.
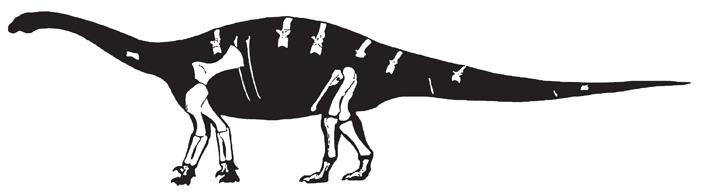
Антетонитр из Южной Африки – один из древнейших завроподоморфов, у которого ладони и передние лапы приспособлены к тяжелой нагрузке. Это крупное, тяжеловесное животное было травоядным или всеядным и могло достигать 10 м в длину. На схеме показаны только те части скелета, которые найдены на данный момент
Передние конечности антетонитра являются чем-то средним между легкими хватательными передними лапами завроподоморфов (вроде платеозавра) и колоннообразными, поддерживающими вес передними конечностями завропод. Похоже, лучевая кость – кость руки, которая идет от запястья до локтя «со стороны большого пальца», изменила свое положение во время эволюции, и положение ладоней тоже изменилось с «внутрь» на «назад». Теперь стало возможно опереться пальцами о землю, а сами пальцы стали лучше приспособлены для переноса тяжести.
Происхождение и анатомия завропод
Если окинуть взглядом все, что мы уже знаем, можно увидеть, что окаменелости завроподоморфов ясно показывают историю эволюционных изменений с начала существования группы. Небольшие двуногие легковесные завроподоморфы появились в позднем триасе и, по-видимому, были плотоядными. От них произошли куда более крупные и тяжелые виды, у которых шеи были длиннее, черепа меньше, а зубы больше подходили для питания листвой. Впрочем, острые зубы в передней части челюсти говорят о том, что эти животные все еще могли быть всеядными. Несколько групп крупных завроподоморфов с более длинными шеями расселились по всему миру и с тех пор постоянно присутствовали в различных экосистемах более 30 млн лет. Благодаря изменениям в строении кистей, анатомии передних и задних конечностей, дальнейшему увеличению размера тела и другим изменениям от одной из линий произошли завроподы.
По форме тела и конечностей завропод очевидно, что они жили на суше. У них высокая и относительно узкая грудная клетка, конечности обычно длинные и тонкие, а ладони и ступни компактные и не так уж велики по сравнению с размером животного. Износ зубов показывает, что они питались наземными растениями, например хвойными и папоротниками, а большинство окаменелостей завропод происходят из сезонно засушливых мест, например парковых лесов. В таких местах сохранились тысячи следов и яиц завропод.
Удивительно, но, несмотря на все эти доказательства, в конце XIX и начале XX в. был период, когда их считали водными животными, которые всю жизнь лениво плескались в озерах и болотах, поедая водоросли. Считалось, что длинная шея была нужна, чтобы дотягиваться до поверхности воды, стоя на дне озера, а ноздри якобы располагались сверху головы, так что они высовывались на поверхность, когда завроподу нужно было вдохнуть. Не совсем ясно, откуда появились эти «водяные гипотезы». Возможно, дело в том, что Ричард Оуэн изначально определил английского среднеюрского завропода цетиозавра как морскую рептилию, похожую на кита, и это могло стать примером для других ученых. Другое возможное объяснение: отложения, в которых находились завроподы, поначалу ошибочно определили как гигантские тропические болота, и это могло повлиять на идеи о поведении завропод.
Как обычно, было бы неверным говорить, что у всех ученых было одно мнение. В Великобритании Гидеон Мантелл в 1852 г. и Джон Филлипс в 1870 г. утверждали, что завроподы жили на суше. Отниел Марш и Эдвард Коуп, великие американские специалисты по динозаврам конца 1800-х гг., тоже писали в своих публикациях 1877 г., что завроподы были наземными животными вроде жирафов. Элмер Риггс, описавший нового длинношеего позднеюрского завропода брахиозавра, в нескольких публикациях в начале 1900-х гг. выступал за их сухопутный образ жизни. Напоследок стоит сказать, что даже современное представление о завроподах как о полностью сухопутных животных не исключает возможность, что некоторые виды вполне могли отдыхать или плавать в воде или заходить в озера и реки, чтобы поесть водных растений. Однако по большей части они были сухопутными, как современные слоны или жирафы.

Некоторые завроподы были необычными и не укладывались в формат «длинная шея, длинный хвост». Брахитрахелопан из поздней юры Аргентины – необычный завропод с короткой шеей. Тем не менее у него колоннообразные конечности, а передние лапы приспособлены для поддержания веса
Ноги завропод куда необычнее, чем можно было предположить. Кости передних конечностей у некоторых видов были длинными и изящными, у других – короткими и толстыми, но у них всегда было особое колоннообразное расположение, идеально подходящее для поддержания веса. Особенно необычны передние конечности. Одно из самых интересных эволюционных преобразований в завроподовой истории связано с тем, как передняя конечность, изначально приспособленная для хватания и сражений, превратилась в поддерживающую колоннообразную конструкцию. Завроподы не ходили, расставив пальцы или опираясь ладонью о землю. Вместо этого длинные кости, из которых состоит ладонь (пястные), располагались в виде полукруглого вертикального блока, вогнутого сзади. Кости пальцев были короткими, редуцированными, а у большинства завроподовых вообще отсутствовали. То же касается когтей: у продвинутых завропод вообще не было когтей на передних лапах, кроме одного на большом пальце, а у многих видов не было и его.
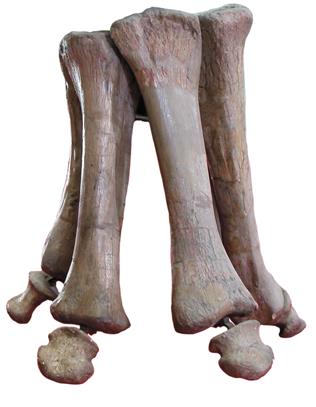
Передние конечности завропод уникальны. Часто на них не было ни когтей, ни копыт, а пястные кости – кости, которые образуют ладонь, – формировали нечто вроде трубы или колоннады. На фото: передняя конечность африканского брахиозавра жираффатитана
Напротив, задние конечности были широкими и длинными, и пять пальцев ног как бы продолжали мощную овальную или округлую подошву. Голеностопный сустав находился близко к земле. Следы и сочлененные скелеты показывают, что гигантская жировая подушка смягчала нижнюю часть стопы и поддерживала плюсневые кости над землей. Три или четыре больших изогнутых когтя торчали на пальцах ног на внутренней стороне стопы. Такие ноги явно предназначались для поддержания веса, и тот факт, что задние конечности завропод почти всегда намного больше передних, согласуется с другими данными, показывающими, что большая часть веса была перенесена в заднюю часть тела, рядом с тазом.

Платеозавр и его родственники были менее приспособлены к растительноядному образу жизни, чем более поздние завроподы. У платеозавра была более узкая пасть, а зубы меньше подходили для измельчения и срывания растений
Архаичные завроподы
Ранние завроподы из позднего триаса и ранней юры демонстрируют, как у животных вроде меланорозавра конечности полностью адаптировались для поддержания большого веса. Череп тоже изменился, что позволило им хватать больше пищи за один укус. Морда завропод была намного шире, чем у ранних завроподоморфов, нижние челюсти имели U-образную, а не V-образную форму, а рядом с основаниями зубов появились необычные костные пластины, усиливающие зубы для укуса. Сильно увеличенные носовые отверстия также являются заметной особенностью ранних завропод. Мы не знаем, зачем им были нужны такие огромные носовые отверстия. Возможно, они помогали регулировать температуру тела, издавать громкие резонирующие звуки или улучшали обоняние. Еще до начала юры некоторые ранние завроподы превратились в гигантов с ногами-колоннами. Например, исанозавр из позднего триаса Таиланда мог достигать 15 м в длину. Это в два с лишним раза меньше более поздних завропод, но по сравнению с ранними завроподоморфами он был великаном.
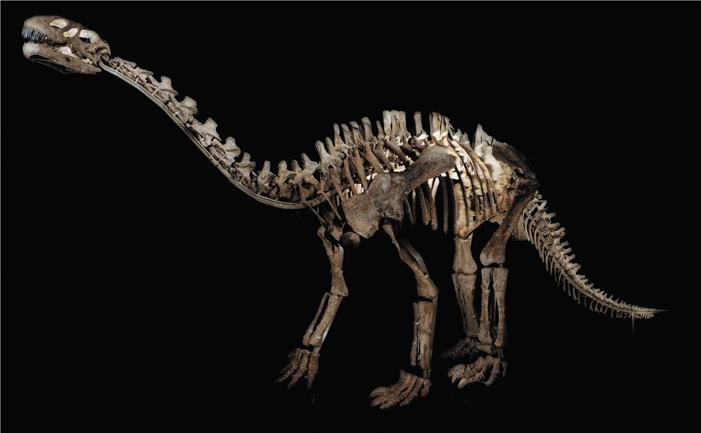
Шунозавр – один из самых известных архаичных завропод. При длине около 10 м он был намного больше большинства ранних завроподоморфов и мог лучше поддерживать большой вес. Более поздние завроподы стали еще крупнее
К ранней и средней юре появились привычные нам завроподы: например, вулканодон из Зимбабве и шунозавр из Китая. Шунозавр необычен своей шипастой «булавой» на кончике хвоста. Такие дубинки или шипы на хвосте были у нескольких видов. Возможно, с их помощью завроподы защищались от теропод, а может быть, сражались друг с другом за самок или территорию. Следы изношенности показывают, что зубы ранних завропод касались друг друга кончиками. Вообще это не типично для динозавров, но свойственно завроподам. Предположительно, этот признак закрепился, так как он позволял быстро срывать много растительного материала.
Туриазавры, мамэньсизавриды и цетиозавры
В течение средней и поздней юры от похожих на шунозавра предков произошло множество новых групп. Их шеи стали больше, системы воздушных мешков в костях скелета усложнились, а сами они увеличились в размерах. Они распространились по всему миру и в поздней юре стали доминирующими крупными травоядными. С исторической точки зрения одним из наиболее значимых из этих животных является цетиозавр, которого обнаружили в среднеюрских породах Англии. Название ему дал сам Ричард Оуэн в 1841 г. Позднее в США нашли и более полные скелеты, но цетиозавр по-прежнему остается важным для понимания эволюции завропод. У него было 12 или 13 позвонков в шее, и форма тела и шеи предполагает, что он был универсальным травоядным и мог находить пищу на вершинах деревьев, на земле и между ними.

Цетиозавр известен по двум неполным скелетам и многочисленным фрагментам. По-видимому, это был «обычный» завропод без каких-либо особенностей, характерных для более поздних групп
Цетиозавр близок к предкам нескольких других завропод. Среди них туриазавры, которые пока что найдены только в Западной Европе. Их отличительная черта – крупные ложковидные зубы с коронкой в форме сердца, а также мощные передние конечности. Некоторые из них были огромными, наравне с самыми большими завроподами. Например, туриазавр, открытый в 2006 г., возможно, был более 25 м в длину и весил более 40 т.

Мамэньсизавр – один из самых длинных завропод. В некоторых музеях (как на фото, например) его скелеты установлены с длинным и плоским черепом, как у диплодока. Это неверно. На самом деле его морда была гораздо короче и выше
У небольшой группы в основном восточноазиатских завропод были особо длинные шеи, состоящие из длинных низких позвонков. Это мамэньсизавриды, названные по типовому роду мамэньсизавра из поздней юры Китая. У них часто встречаются длинные и стройные конечности, и у них больше шейных позвонков, чем у других групп (более 15 против обычных 12 или около того). Это значит, что они объедали высокие кроны деревьев, намного выше, чем могли дотянуться другие завроподы и прочие травоядные динозавры. Опять же, некоторые мамэньсизавриды достигали исполинских размеров, больше 30 м в длину, и, возможно, весили больше 75 т. У самых крупных видов шея могла достигать невероятных 17 м. Другая особенность – небольшая «булава» на хвосте у нескольких видов. Зачем она была нужна, опять-таки точно неизвестно.
Диплодокоиды и макронарии
Большинство видов завропод относятся к одной из двух основных групп, которые появились в середине юрского и сохранились до конца (или почти до конца) мелового периода. Это диплодокоиды и макронарии. К диплодокоидам относится несколько групп, у которых обычно длинные прямоугольные морды, зубы-колышки и длинные хвосты, кончик которых похож на хлыст. Также у них короткие, мощные передние конечности. Диплодок, апатозавр и бронтозавр, найденные в знаменитой верхнеюрской формации Моррисон в США, – диплодокоиды. Они входят в кладу под названием диплодоциды.
К диплодокоидам также относятся сравнительно небольшие, короткошеие дикреозавриды и диковинные реббахизавриды из мелового периода. Сюда входят самые странные завроподы. Например, у амаргазавра – дикреозаврида из раннего мела Аргентины – были длинные парные шипы, растущие назад и вверх из позвонков шеи. А нигерзавр – маленький реббахизаврид из раннего мела Нигера – обладал хрупким черепом, на котором ряды зубов повернуты так, что располагаются вдоль переднего края челюсти, а не по бокам.

Черепа диплодокоидов – как у этого диплодока – необычны по сравнению с другими завроподами. Отверстия для ноздрей расположены высоко на лбу. Однако следы кровеносных сосудов и другие структуры показывают, что мясистые ноздри, скорее всего, располагались ближе к краю пасти
Длинные шеи, необычные морды и зубы-колышки диплодоцид вроде диплодока и апатозаврапредполагают, что у них был свой особый образ жизни, ведь они сильно отличаются от других завропод. Одна из гипотез состоит в том, что они объедали кроны деревьев, просовывая шею в самую гущу листвы, куда другие не могли дотянуться, причем, возможно, даже приподнимались на задние лапы. Другая заключается в том, что они тянулись к земле и поедали стелющиеся растения. Мы еще поговорим о питательных стратегиях диплодоцид в главе 4.
В отличие от диплодокоидов, макронарии обычно имели большие носовые отверстия (это объясняет их название, которое означает «большие носы») и более округлую морду. Позднеюрские завроподы камаразаври брахиозавр из США и жираффатитан из Танзании относятся к макронариям, как и титанозавры – самые разнообразные и долгоживущие завроподы. Раньше считалось, что макронарии были необычными завроподами, чья анатомия черепа сильно отличается от предков. Теперь, когда мы знаем, что ранние завроподы вроде шунозавра обладали тупыми мордами и крупными ноздрями, стало ясно, что как раз черепа макронариев обычные, а вот низкие черепа диплодоцид с маленькими ноздрями – исключение из правил.

На черепе камаразавра можно увидеть типичные признаки макронариев: короткие мощные челюсти и огромное отверстие для ноздрей (большая дыра спереди)
В последнее время произошел всплеск находок титанозавров в Америке, Европе, Африке, Азии и на Мадагаскаре. Оказалось, в течение своей истории они породили огромное разнообразие видов. Некоторые были гигантскими, наравне с самыми большими завроподами, а следовательно, самыми крупными наземными животными всех времен. Аргентинозавр из позднего мела Аргентины, возможно, был 30 м длиной, а весил более 70 т. Патаготитан – тоже из позднего мела Аргентины – был примерно такой же длины и почти наверняка весил столько же. Многие другие титанозавры были «среднего размера» по завроподовым меркам, скажем, 14 м или меньше в длину и менее 15 т весом. Другие же, такие как мадьярозавр из Румынии, были совсем крошечные (для завропод) – всего 8 м длиной, а весили, возможно, не более тонны.
Форма и пропорции титанозавров тоже были разнообразны. К примеру, у позднемелового сальтазавраиз Аргентины были короткие мощные конечности и обычная шея. Другие были более стройными, а шея их была чрезвычайно длинной. Таким, например, был рапетозавр из позднего мела Мадагаскара. Так как титанозавры относятся к макронариям, изначально у них были широкие, относительно короткие морды и зубы с широкой коронкой, по крайней мере у некоторых. У других же морды были длинные, а коронка зубов – узкая. У некоторых титанозавров с удлиненной мордой пасть была расширена, почти как утиный клюв. Все это значит, что в течение мелового периода титанозавры приспосабливались к разным стратегиям питания и, возможно, заняли некоторые ниши, которые раньше занимали диплодокоиды – еще один пример эволюционной конвергенции.
Совсем недавно, еще в 1990-х гг., считалось, что завроподы жили в основном в юрском периоде, а к меловому, по сути, исчезли. Теперь мы знаем, что это мнение было ошибочным и что завроподы обитали на многих континентах на протяжении большей части мелового периода. При этом вместо застоя или стагнации они постоянно развивали новые анатомические особенности и новые способы срывания растений.
Назад: Тероподы: хищные динозавры и птицы
Дальше: Птицетазовые динозавры: бронированные, утконосые, рогатые и их родственники

