Книга: Вопрос жизни. Энергия, эволюция и происхождение сложности
Назад: Часть I Проблема
Дальше: Глава 2 Что значит жить?
Глава 1
Что такое жизнь?
Сорок два радиотелескопа в горах на севере Калифорнии не спускают глаз с неба. Проект SETI (Search for Extraterrestrial Intelligence, “поиск внеземного разума”) полвека – безуспешно – ловит сигналы от других цивилизаций. Даже подвижники уже не особенно надеются на успех. Впрочем, несколько лет назад, когда у SETI иссякли деньги, после обращения к публике средства были собраны и Антенная решетка Аллена вновь заработала. Мне кажется, в этом проекте воплощено неясное, тревожное чувство потерянности человека во Вселенной. Его работа наводит на мысли о хрупкости научного подхода как такового. Он дает нам иллюзию всеведения. Современные технологии настолько сложны, что почти уже неотличимы от научной фантастики. Тем прочнее укореняется наивная надежда, что мы не одни во Вселенной (ведь существа с других планет – частый атрибут жанра). Однако не стоит забывать, что эта надежда не имеет под собой научной основы.
Но даже если телескопы никогда не обнаружат жизнь вне нашей планеты, они не бесполезны. Может быть, вообще невозможно смотреть в эти телескопы неправильным образом и все способы верны, потому что настоящая их польза в другом. Что мы ищем в космосе? Найдутся ли где-нибудь разумные формы жизни, настолько похожие на нас, что они также будут пользоваться радиосвязью? Считаем ли мы, что основой живой материи обязательно должен быть углерод? Является ли вода необходимой составляющей жизни? А кислород? Эти вопросы на самом деле должны быть обращены не к инопланетной жизни, а к земной – почему она именно такая, какой мы ее видим. Но до биологов эти вопросы доходят, лишь отразившись от тарелок радиотелескопов. Проблема в том, что суть науки – это предсказания. Главные вопросы физики – почему физические законы таковы, какими мы их знаем, какие фундаментальные принципы определяют устройство Вселенной. В биологии нет законов, сопоставимых по предсказательным способностям с физическими. Даже эволюционная биология в этом отношении ужасающе неточна. Мы располагаем подробнейшими сведениями о молекулярных механизмах эволюции и об истории жизни на нашей планете. Но при этом мы не знаем, что в истории земной жизни – следствие случайности, а что, напротив, обусловлено физическими законами.
Нельзя сказать, что за эти проблемы никто не брался. На этом поле ведут игру крупнейшие ученые, в том числе нобелевские лауреаты, но, несмотря на остроту их ума и познания, им так и не удалось прийти к согласию. Сорок лет назад, в начале эры молекулярной биологии, французский биолог Жак Моно написал знаменитую книгу “Случайность и необходимость”, полную доводов в пользу того, что жизнь возникла благодаря редчайшему стечению обстоятельств и что мы, скорее всего, одиноки в мертвой Вселенной. Последние строки этой книги – близкий к поэзии сплав науки и метафизики:
Древние скрижали разбиты. Человек знает наконец, что одинок в равнодушной необъятности Вселенной, в которой появился благодаря случайности. Древние заветы ничего не скажут ни о его судьбе, ни о его предназначении. Царствие Небесное или тьма под ногами – выбирать ему самому.
Тогда же появились люди, которые придерживались противоположной точки зрения. Они считали, что происхождение жизни – это неизбежный результат космохимических процессов. Согласно этой гипотезе, жизнь способна зарождаться почти везде и довольно быстро. Но предположим, что на какой-нибудь планете начинает развиваться жизнь: к чему приведет это развитие? И на этот вопрос у нас нет четкого ответа. Может быть, разные пути развития жизни, подчиняясь физическим ограничениям, независимо приведут примерно к одним и тем же результатам. Так, гравитация вынуждает способных к полету существ быть легкими и обладать чем-то вроде крыльев. Еще более сильное заключение – что эволюция неизбежно должна привести к возникновению клеток: блоков, содержимое которых отделено от окружающей среды. Если такие закономерности универсальны, то инопланетная жизнь должна напоминать земную. С другой стороны, развитие жизни зависит и от случайности. Например, в результате глобальных катаклизмов наблюдается эффект “бутылочного горлышка”. Именно такой эффект проявился после падения метеорита, из-за которого вымерли динозавры. Мысленно пустим время вспять и перенесемся на полмиллиарда лет, в кембрийский период, когда в палеонтологической летописи появляются животные. Если позволить миру повторить свое развитие начиная с этого момента, будет ли он похож на наш? Кто знает! Возможно, в параллельной реальности Земля населена гигантскими сухопутными осьминогами.
Одна из причин направлять телескопы в космическое пространство состоит в том, что здесь, на Земле, мы имеем дело только с земными процессами. Мы располагаем единственным примером того, как может выглядеть жизнь, а значит, не можем предсказывать закономерности развития, опираясь на статистические данные. Мы не можем сказать, что именно определяло (если определяло) ход эволюции жизни, поскольку нам не с чем ее сравнить. Правда, если сказать об этом было бы совсем нечего, написание моей книги (как и многих других) потеряло бы смысл. Законы физики повсюду во Вселенной одни и те же. То же касается химических элементов и их свойств – и, значит, химические процессы также универсальны. У земной жизни много странных свойств, например половое размножение и старение. Если бы мы знали, каким образом химический состав и другие базовые свойства Вселенной влияют на ход событий, это заменило бы большую выборку: мы сумели бы указывать, исходя из базовых принципов, вероятность приобретения тех или иных свойств и, в конечном счете, смогли бы объяснить, почему жизнь такова, какой мы ее знаем. Впрочем, жизнь на Земле трудно счесть единственным примером: это бесконечное многообразие организмов, эволюционирующих в неограниченной временной перспективе. Но все же теория эволюции не способна объяснить, почему развитие жизни пошло именно этим путем. Я вовсе не хочу сказать, что теория эволюции неверна. Она верна, однако не обладает достаточной предсказательной силой. Идея этой книги следующая: есть действительно важные факторы, определяющие ход эволюции, и это – энергетические ограничения. Рассматривая жизнь с позиции этих ограничений, мы можем вывести из базовых принципов некоторые из ее фундаментальных свойств. Прежде чем перейти к рассмотрению этих ограничений, стоит обсудить, почему эволюционная биология обладает слабой предсказательной силой и почему мы долгое время едва ли замечали существование проблемы энергетических ограничений. Лишь в последние годы стало ясно (пока только тем, кто следит за развитием эволюционной биологии), что в центре биологической науки – ошеломительный пробел.
В этом состоянии дел мы можем винить ДНК. Забавно, что эра современной молекулярной биологии со всеми ее впечатляющими технологиями, по сути, началась с физики, точнее, с книги Эрвина Шредингера “Что такое жизнь?” (1944). Шредингер осветил два самых существенных момента. Во-первых, жизнь каким-то образом противостоит общей тенденции распада – повышению энтропии, предписываемого вторым началом термодинамики. Вторая очень важная идея Шредингера такова: жизнь смогла уклониться от этого закона благодаря генетическому материалу. Шредингер предугадал, что генетический материал представляет собой “апериодический кристалл”, структура которого не является строго повторяющейся и поэтому может функционировать как “код”. (Считается, что это первое употребление данного термина в биологической литературе.) Шредингер, подобно большинству биологов того времени, считал, что обсуждаемый квазикристалл должен представлять собой белок, однако Уотсон и Крик обнаружили, что ДНК подходит на эту роль куда лучше. В 1953 году в своей второй статье в журнале “Нейчур” они указывали: “Таким образом, вероятно, определенная последовательность оснований является кодом, который содержит генетическую информацию”. Эта фраза – краеугольный камень современной биологии. Биология в нынешнем мире – это информация, геномные последовательности, хранящиеся в компьютерах, а жизнь – преобразования информации.
Изучение геномов открыло нам целый мир, чудесный и таинственный. Бесконечные строки кода (в случае нашего генома – 3 млрд символов) можно читать, как постмодернистский роман. Местами он представляет собой нарратив, перемежающийся повторами, стихами, пустыми страницами, потоком сознания, своеобразной пунктуацией. Лишь менее 2 % нашего генома кодирует белки. Несколько больше отведено под регуляторные механизмы. Рассуждение же о функциях остальной части генома может подвигнуть моих любезных коллег на нескончаемый спор, поэтому я не буду об этом говорить (кроме того, для нас это не так важно). Мы знаем, что геномы могут кодировать десятки тысяч генов и хранить информацию об огромном числе изощренных регуляторных механизмов, контролирующих такие сложные процессы, как превращение гусеницы в бабочку или взросление человека. Сравнительный анализ геномов животных, растений, грибов и одноклеточных амеб показывает, что процессы в этих организмах очень схожи. Несмотря на то, что их геномы чрезвычайно различаются по размеру и структуре, мы имеем дело с вариантами одних и тех же генов, регуляторных элементов, эгоистичных репликаторов (например вирусов) и одними и теми же участками некодирующих повторов. Геномы лука, пшеницы и амебы размерами превосходят наш и содержат больше генов. Размеры геномов амфибий, например лягушек и саламандр, могут отличаться друг от друга более чем на два порядка: геном некоторых видов саламандр больше нашего в 40 раз, а размер генома некоторых лягушек едва достигает трети размера генома человека. Если коротко описать наложенные на геном архитектурные ограничения, то получится: “Ничего невозможного”.
Это очень важно. Ведь если геном – это информация и не существует фундаментальных ограничений его структуры и размера, – значит, у закодированной в геноме информации ограничений также нет. Разумеется, это не означает, что на геном не наложены никакие ограничения. На него воздействует естественный отбор. В геноме происходит множество случайных процессов: генные, хромосомные и полногеномные дупликации, инверсии, делеции и встраивание паразитической ДНК. Во что все это выльется, зависит от многих факторов, например от экологической ниши организма, межвидовой конкуренции и размера популяции. Нам кажется, что влияние этих факторов нельзя предугадать, что они просто часть среды. Если бы все ее параметры были точно известны, то, может быть, мы смогли бы предсказать размер генома некоторых организмов. Но на деле мы сталкиваемся с бесконечным множеством видов, обитающих в бесконечно разнообразных условиях: внутри чужих клеток, в городах, в океанских глубинах под огромным давлением. Так что ограничения, наложенные на геном, лучше сформулировать так: “Возможно все”. Видимо, разнообразие геномов определяется разнообразием факторов среды. Структура геномов не позволяет увидеть будущее, но может рассказать о прошлом: с чем пришлось столкнуться организму за историю его развития.
Вернемся к иным мирам. Если основа жизни – это информация и она ничем не ограничена, то невозможно предсказать, как может быть устроена жизнь на другой планете – за исключением того, что она не должна противоречить законам физики. Стоит появиться некоему носителю наследственной информации (ДНК или нечто похожее), как на эволюцию перестают действовать информационные ограничения. И тогда определить ее путь становится невозможным: его будут определять условия среды, предшествующие случайные события и давление отбора. Но если мы вернемся с небес на Землю, то увидим, что это утверждение справедливо, когда мы говорим о невероятном разнообразии современных организмов, но не годится для большей части истории планеты. Судя по всему, существовали ограничения, которые невозможно сформулировать исходя из данных геномов, предшествующей истории и условий среды, но они миллиарды лет контролировали развитие жизни.
Краткая история первых двух миллиардов лет жизни
Нашей планете около четырех с половиной миллиардов (иначе говоря, 4500 млн) лет. В юности ей пришлось нелегко: не исполнилось ей и 700 млн лет, как она пережила Позднюю метеоритную бомбардировку (тогда Солнечная система приходила в себя после формирования). Земля испытала колоссальной силы столкновение с астрономическим объектом размером с Марс, в результате которого, вероятно, появилась Луна. Земная кора непрерывно трансформируется из-за геологической активности, но нетронутая древняя поверхность Луны хранит свидетельства бомбардировки в виде кратеров. Это было выяснено по датировке осколков лунных скал, привезенных астронавтами проекта “Аполлон”.
Хотя на Земле нет горных пород сопоставимого возраста, есть и другие способы получить представление об условиях на древней Земле. Так, состав цирконов (крошечные, меньше песчинок, кристаллы силиката циркония, нередкие в горных породах) позволяет предположить, что океаны появились на Земле гораздо раньше, чем считалось. На основании датировки уран-свинцовым методом можно сказать, что некоторые из этих удивительно прочных кристаллов образовались 4,4–4 млрд лет назад, а затем в виде зерен вошли в состав осадочных пород. Кристаллы цирконов надежно сохраняют внутри химические вещества, которые попали туда еще на стадии формирования кристаллов, а значит, можно определить, в каких условиях это происходило. Химический состав древних цирконов указывает на то, что они образовались при относительно низких температурах и в присутствии воды. Не слишком похоже на одну из тех жутких картин вулканического ада, каким художники любят изображать гадейский период (катархей). Кристаллы циркона говорят обратное: это был относительно спокойный водный мир с редкими островами.
Результаты изучения цирконов не подтверждают и ту давнюю гипотезу, что атмосфера древней Земли была наполнена метаном, водородом и аммиаком и они, реагируя друг с другом, формировали органические молекулы. Некоторые редкоземельные элементы, например церий, представлены в составе цирконов в основном своей окисленной формой. Высокое содержание церия свидетельствует о том, что атмосфера в то время состояла главным образом из окисленных газов вулканического происхождения: диоксида углерода, водяного пара, азота и диоксида серы. Эта смесь по составу не отличалась от современной атмосферы – за исключением того, что в ней вплоть до возникновения фотосинтеза отсутствовал кислород. Реконструкция облика давно исчезнувшего мира опирается лишь на состав песчинок, но это гораздо лучше, чем совсем ничего. Это довод в пользу того, что древняя Земля была удивительно похожа на нынешнюю. Внезапное вмешательство астероида могло повлечь частичное испарение океанов, но вряд ли это могло сильно повредить бактериям, живущим в глубине (если, конечно, к тому времени они успели эволюционировать).
Самые ранние свидетельства жизни столь же эфемерны: едва различимые ее следы обнаружены в горных породах комплексов Исуа и Акилия (около 3,8 млрд лет) на юго-западе Гренландии (рис. 2). Эти следы представляют собой не окаменевшие организмы и даже не сложные молекулы организменного происхождения (биомаркеры). Это просто графит со смещенным соотношением типов углеродных атомов. Углерод состоит из двух стабильных типов атомов – изотопов, которые немного отличаются друг от друга по массе. Ферменты (белки, катализирующие реакции в клетках) охотнее взаимодействуют с легким изотопом углерода 12С, поэтому именно он накапливается в живых организмах. Можно представить, что атомы углерода двух типов – это мячики для пинг-понга, которые немного отличаются по массе. Более легкие мячики 12С отскакивают чуть быстрее и поэтому имеют больше шансов столкнуться с ферментами и войти в состав органических веществ. Содержание тяжелого 13С в природном углероде составляет лишь 1,1 %. Этот изотоп хуже усваивается живыми организмами и поэтому накапливается в океанах и включается в карбонатные осадочные породы, например в известняк. Эти мелкие различия изотопного состава используются как диагностический признак для обнаружения следов жизни. Изотопному фракционированию в живых клетках подвергаются не только углерод, но и железо, азот и сера. В углистых включениях в Исуа и Акилии обнаружено смещение изотопных составов и этих элементов.
Каждый пункт этих исследований можно поставить под сомнение: от датировки горных пород до самого существования графитовых гранул, в которых видят следы жизни. Более того, выяснилось, что смещение изотопного состава, вероятно, связано не только с деятельностью живых организмов. Фракционирование изотопов, которое мы принимаем за проявление жизни, может быть следствием геологических процессов в гидротермальных источниках. И даже если возраст горных пород Гренландии определен верно и они действительно содержат углерод со смещенным изотопным составом, то это еще не доказывает, что тогда существовала жизнь. Впрочем, не стоит расстраиваться: по большому счету, никакой разницы нет. Беру на себя смелость утверждать, что различие между “живой”, геологически активной планетой и живой клеткой – лишь вопрос установления того, что такое “жизнь”. Никаких четких границ между ними нет: геохимия и биохимия плавно переходят друг в друга. Можно считать, что указанные горные породы заполняют пропасть между геологией и биологией. Живая планета и произведенная ею жизнь представляют собой единое целое.

Рис. 2. Этапы развития жизни на Земле. На шкале отражены примерные даты ключевых событий (млрд лет). Многие датировки остаются спорными, но ясно, что бактерии и археи появились на 1,5-2 млрд лет раньше эукариот.
Если мы продвинемся по временной шкале на несколько сотен миллионов лет вперед, то увидим уже более отчетливые следы жизни, столь же надежные и весомые, как запечатлевшие их камни Австралии и Южной Африки. Во-первых, это микроокаменелости, очень похожие на клетки. Впрочем, пытаться отнести эти окаменевшие клетки к одной из современных групп живых организмов – занятие неблагодарное. Многие микроскопические окаменелости покрыты слоем углерода с характерным изотопным составом. Но эти изотопные следы жизни гораздо более достоверны, они определенно появились в результате биохимических, а не геохимических процессов. Во-вторых, это образования, похожие на строматолиты – каменные идолы мира бактерий. В строматолитах растущие клетки образуют слои. Нижележащие слои постепенно минерализуются, и образуются удивительные каменные столбы высотой до 1 м. Кроме окаменевших остатков бактерий, 3,2 млрд лет назад на Земле появляются огромные геологические образования площадью в несколько сотен квадратных километров и толщиной в десятки метров – полосчатые железистые формации и углистые сланцы. Мы привыкли считать, что бактерии и минералы принадлежат к двум разным мирам – живому и неживому, однако огромные залежи осадочных горных пород появились в результате жизнедеятельности бактерий. Так образовались и полосчатые железистые формации – удивительные по красоте минералы, расписанные красными и черными линиями: когда бактерии отнимают электроны у ионов двухвалентного железа, растворенных в воде океана (такая форма железа распространена при отсутствии кислорода), образуется нерастворимая ржавчина, оседающая на дно. До сих пор остается загадкой, как в этих горных породах образовались полосы, но смещение изотопного состава и здесь выдает следы жизни.
Обширные железистые отложения указывают на то, что жившие тогда организмы уже научились фотосинтезу. Но это не хорошо нам знакомый тип фотосинтеза, который мы можем наблюдать у растений и водорослей, а его примитивная предковая форма. Во всех формах фотосинтеза энергия света расходуется на то, чтобы оторвать электроны от субстрата, который не склонен их отдавать. Таким образом, энергия света преобразуется в энергию электронов. Затем эти электроны используются, чтобы включить углекислый газ в органические молекулы. В качестве источника электронов в фотосинтезе могут использоваться разнообразные субстраты, но чаще всего – растворимые в воде ионы двухвалентного железа, сероводород и вода. Когда от этих субстратов отрываются электроны, они превращаются в окисленные формы: осадки ржавчины, элементарную серу и кислород. Вода – это, пожалуй, наихудший источник электронов для фотосинтеза, и 3,2 млрд лет назад живые организмы предпочитали пользоваться другими субстратами. Как заметил биохимик Альберт Сент-Дьерди, жизнь – это скитание ищущих покой электронов. Точно неизвестен момент, когда живые организмы научились окислять воду (иначе говоря, применять оксигенный фотосинтез). Согласно наиболее аргументированной версии, оксигенный фотосинтез возник 2,9–2,4 млрд лет назад, незадолго до периода глобальных катаклизмов. Около 2,2 млрд лет назад, перед глобальным оледенением (“Земля-снежок”), происходил масштабный процесс окисления железа в горных породах на поверхности Земли. Так образовались красноцветные отложения, которые указывают на накопления в атмосфере кислорода: “Великое кислородное событие”. Глобальное оледенение также подтверждает, что в атмосфере стал накапливаться кислород. Метан порождает мощный парниковый эффект. Выделяющийся в атмосферу кислород стал реагировать с метаном, концентрация метана снизилась, и это спровоцировало глобальное похолодание.
С возникновением оксигенного фотосинтеза жизнь заполучила полный набор метаболических инструментов. Хотя наше стремительное путешествие сквозь 2 млрд лет истории (а это втрое дольше периода существования животных) не предполагает изложения всех подробностей, здесь мы все же остановимся и поговорим о том, что может рассказать нам временная шкала. Во-первых, жизнь возникла на Земле очень давно (4–3,5 млрд лет назад, если не раньше) в мире, который не слишком отличался от современного: во всяком случае, там было много воды. Во-вторых, 3,5–3,2 млрд лет назад бактерии уже нашли большую долю путей метаболизма, в том числе многочисленные формы дыхания и фотосинтеза. В течение миллиарда лет мир представлял собой огромный бурлящий котел с бактериями, являющий такую изощренность биохимических путей, что остается лишь диву даваться. Изотопное фракционирование свидетельствует о том, что главные энергетические циклы – углерода, серы, железа и т. д. – были в ходу уже 2,5 млрд лет назад. Но лишь с повышением концентрации кислорода, которое началось 2,4 млрд лет назад, жизнь преобразила планету настолько, что это стало заметно даже из космоса. Тогда в атмосфере стала накапливаться химически активная смесь газов, например кислорода и метана, непрерывно пополняемая живыми клетками (с их стороны это было предательство прямо-таки планетарного масштаба).
Среда или гены?
Великое кислородное событие долго считалось главным эпизодом в истории планеты, однако в последние годы его значение изменилось. Новое объяснение играет чрезвычайно важную роль в приведенных в этой книге доказательствах. В рамках прежнего взгляда кислороду отводится роль ключевого фактора среды, обусловливающего развитие жизни. Согласно новому взгляду, кислород не определяет конкретные результаты эволюции: он снимает прежние ограничения и делает возможным появление гораздо более сложных форм жизни. Так, жизнь животных неразрывно связана с активным перемещением в пространстве – поиском пропитания, бегством от хищников. Для этого требуется много энергии, и легко догадаться, что животные не смогли бы существовать без кислорода, использование которого позволяет получить на порядок больше энергии, чем от других форм дыхания. Это утверждение настолько банально, что не возникает и мысли оспаривать его. Но проблема отчасти именно в том, что этот тезис не обсуждается и потому не получает развития. Что ж, раз эту тему обсуждать не хочется, почему бы не принять за аксиому, что животным в любом случае требуется кислород (закрыв глаза на то, что это не всегда так), и не заняться по-настоящему важными вопросами эволюционной биологии, например строением и поведением животных и растений? Или все-таки лучше так не поступать?
Эта позиция в неявном виде поддерживается во всех учебниках. Мы привыкли думать о кислороде как о чем-то приятном и благотворном, однако в контексте древней биохимии это ядовитое и очень реакционноспособное вещество. “Школьный” взгляд гласит: когда уровень кислорода повысился, этот опасный газ выступил жестким фактором отбора в отношении микробного мира. С этим связаны наводящие ужас рассказы о массовом вымирании бактерий (Маргулис назвала это “кислородным холокостом”). Тот факт, что в палеонтологической летописи не осталось следов этого катаклизма, не должен нас беспокоить (мы знаем это наверняка): это было ужасно давно, а те создания были совсем крошечными. Кислород способствовал выработке новых взаимоотношений клеток – симбиоза и эндосимбиоза, в ходе которых те поддерживали взаимовыгодные отношения и с внешними партнерами, и с внутренними, приобретая все новые инструменты для выживания. За сотни миллионов лет появились сложные организмы, так как клетки научились не только справляться с кислородом, но и пользоваться его реакционной способностью – приобрели аэробное дыхание, дающее гораздо больше энергии. Крупные, сложно устроенные аэробные клетки таких организмов “упаковывают” ДНК в специализированный компартмент – ядро, тем самым оправдывая наименование эукариоты (то есть те, что “с настоящим ядром”). Повторяю: это история из учебника. Но в действительности (и я это докажу) дело обстояло иначе.
Сегодня все сложные живые организмы – растения, животные, водоросли, грибы и протисты (крупные одноклеточные, например амебы) – состоят из эукариотических клеток. Эукариоты шли к своему триумфу миллиард лет. Этот период известен теперь (забавно, правда?) как “скучный миллиард” – из-за ничтожного количества сохранившихся окаменелостей. Тем не менее находятся остатки отдельных клеток, относящиеся ко времени 1,6–1,2 млрд лет назад. Эти клетки очень напоминают эукариот: некоторые отлично бы вписались в современные группы вроде красных водорослей или грибов.
Затем, 750–600 млн лет назад, наступил период больших волнений: Земля попеременно покрывалась льдом и оттаивала снова. Вскоре после этого концентрация кислорода стремительно выросла, достигнув почти современного уровня. В это время в палеонтологической летописи появляется большое количество окаменелостей животных. Самые древние – группа симметричных листовидных организмов до 1 м в диаметре, которых большинство палеонтологов считает фильтраторами (хотя другие настаивают, что это лишайники). Речь идет об эдиакарской биоте, представителей которой также называют вендобионтами. Затем большинство этих существ (так же внезапно, как появились) в ходе массового вымирания исчезли, а на заре кембрийского периода (541 млн лет назад) их место заняли животные уже более привычного нам вида. Указанная дата – время “кембрийского взрыва”, заполнившего Землю разнообразными новыми формами, – столь же важна для биологов, как для западных историков 1066 или 1492 годы. Крупные и подвижные, со сложными глазами и чувствительными выростами на теле, эти устрашающие хищники и их не менее жуткие, закованные в броню жертвы вышли на арену эволюции и отыграли партию так, что Дарвин много бы дал, чтобы лично взглянуть, что творил на древней Земле естественный отбор.
Насколько этот сценарий близок к реальности? На первый взгляд, он довольно правдоподобен. Мне, впрочем, кажется, что суть его неверна, а добрая часть деталей сценария при ближайшем рассмотрении сомнительна. Суть относится к взаимодействию генов и среды. Целый сценарий завязан на кислороде, которому отводится роль ключевого фактора среды: кислород, устранив ограничения, дал зеленый свет генетическим преобразованиям. Концентрация кислорода повышалась дважды: 2,4 млрд лет назад, во время Великого кислородного события, и 600 млн лет назад, ближе к концу неимоверно долгого докембрия (рис. 2). Всякий раз при увеличении концентрации кислорода исчезали ограничения, сдерживающие развитие живых существ. После Великого кислородного события в новом мире, полном опасностей и возможностей, клетки приспособились оказывать друг другу услуги при помощи серийных эндосимбиозов, постепенно приближаясь к настоящей эукариотической сложности. Когда уровень кислорода поднялся во второй раз, перед “кембрийским взрывом”, физические ограничения были окончательно устранены. Это произошло будто по мановению волшебной палочки. Разумеется, организмы изменились не под прямым воздействием кислорода, а под влиянием обусловленного им преобразования селективного ландшафта. В новых условиях избавленные от ограничений геномы стали активно развиваться, и жизнь расцвела, породив великое множество форм и заполнив все возможные экологические ниши.
Такую точку зрения на эволюционный процесс можно интерпретировать в терминах диалектического материализма, принципов которого придерживались некоторые видные биологи-эволюционисты в первой половине ХХ века, когда формировался неодарвинизм. Гены и окружающая среда представляют собой взаимопроникающие противоположности, иначе известные как “природа и воспитание”. Всю биологию можно свести к изучению генов и того, как они реагируют на изменения окружающей среды. И что же: жизнь на этом заканчивается? Нет: жизнь включает не только взаимодействие генов и среды, но также клетки и ограничения, которые определяются их структурой. (Ниже я объясню, какие это ограничения и почему их нельзя свести к среде и генам.) Сценарии, предлагаемые в рамках двух противоположных мировоззрений, сильно различаются.
Рассмотрим первую позицию, трактующую эволюцию в терминах взаимодействия генов и окружающей среды. Самый существенный фактор, ограничивающий развитие жизни на древней Земле – отсутствие кислорода. Его накопление, соответственно, запустило бурный эволюционный процесс. Кислород вторгся в жизнь всех контактирующих с ним организмов, и тем пришлось приспосабливаться. Клетки, которые оказались лучше подготовлены к жизни в аэробной среде, выжили и размножились, а остальные погибли. Но на Земле возможно существование множества экологических ниш. Когда кислород накапливался, он не просто насыщал все возможные среды на планете. Он окислял минералы, содержащиеся в горных породах и растворенные в океанской воде, и продукты этого окисления накапливались и в анаэробных средах. Живые организмы получили доступ к нитратам, нитритам, сульфатам, сульфитам и другим окисленным соединениям. Клетки могут использовать эти вещества для дыхания вместо кислорода, и анаэробное дыхание получило широкое распространение в аэробном мире. Все это увеличивает число возможных экологических ниш в новом мире.
Представьте себе случайную смесь клеток. Некоторые, похожие на амеб, питаются путем фагоцитоза, то есть заглатывая другие клетки. Вторые способны фотосинтезировать. Третьи питаются осмотрофно: переваривают пищу вне тела, как грибы. Допуская, что структура клеток не ставит никаких непреодолимых препятствий на пути их развития, мы можем предположить, что эти разные типы клеток могли произойти от разных бактериальных предков. Некоторые бактерии были способны к примитивному фагоцитозу, другие – к простой форме осмотрофного питания, третьи умели фотосинтезировать. Их потомки стали более специализированными и довели свои умения до совершенства.
В общем, если бы повышение уровня кислорода ускоряло темпы появления новых организмов, следовало бы ожидать полифилетической радиации, когда неродственные (из разных групп) клетки или организмы, быстро эволюционируя, порождают новые виды, занимая свободные экологические ниши. В некоторых случаях так и происходит. Пример – “кембрийский взрыв”, в результате которого возникли десятки таксонов животных, среди которых губки, иглокожие, членистоногие и черви. Полифилетическая радиация животных в кембрии сопровождалась сходной радиацией среди водорослей, грибов и протистов, например инфузорий. Экологическая ситуация очень усложнилась, и это повлекло дальнейшие изменения в мире живых организмов. Мы не знаем, стал ли “кембрийский взрыв” следствием именно повышения концентрации кислорода, но ясно, что изменение окружающей среды способно радикально изменить механизмы отбора. Произошло нечто, что навсегда изменило мир.
Если же главный фактор, контролирующий развитие живых организмов, обусловлен их собственной структурой, мы наблюдали бы совершенно иную картину. Пока существуют такие структурные ограничения, они не позволяют организмам заметно изменяться в ответ на изменения среды. В таких условиях наблюдаются долгие периоды стазиса с очень редкими случаями монофилетической радиации. Иными словами, ситуации, когда живые существа преодолевают ограничения, наложенные их собственной структурой, чрезвычайно редки. Но если этот сценарий реализуется в каком-нибудь таксоне, происходит радиация этой группы организмов, и впоследствии они занимают все свободные экологические ниши (хотя, возможно, для этого им придется дождаться изменений среды). Разумеется, и такие процессы мы можем наблюдать. “Кембрийский взрыв” сопровождался радиацией групп животных, но сами животные в ходе эволюции появились всего единожды: все эти группы восходят к общему предку. Точно так же дело обстоит с растениями. Сложной многоклеточности с разделением клеток на соматическую и зародышевую линии достичь довольно тяжело. Отчасти это связано с тем, что требуются механизмы, способные очень точно управлять развитием организма, вплоть до каждой клетки. Но примитивные формы многоклеточности распространены очень широко и независимо появились примерно в 30 группах организмов (водоросли, грибы, слизевики и т. д.). Однако есть особый случай, когда ограничения, наложенные структурой клетки, преобладают над любыми другими ограничениями. Это происхождение эукариотической клетки от бактерий после Великого кислородного события.
Прореха в ткани биологии
Если бы сложные клетки появились как ответ на повышение содержания кислорода в атмосфере, можно было бы предположить полифилетическую радиацию: независимое происхождение эукариот от разных групп бактерий. Так, фотосинтетические бактерии могли бы дать начало более крупным и сложным водорослям, осмотрофные бактерии – грибам, подвижные хищные клетки – фагоцитам. Такое развитие сложности могло происходить путем обычных мутаций, генетического обмена и естественного отбора. Другой вариант – слияние и вхождение в эндосимбиотические отношения, в соответствии с теорией серийных эндосимбиозов Линн Маргулис. В любом случае, в отсутствие ограничений касательно структуры клетки повышение концентрации кислорода должно было сделать возможным приобретение сложности вне зависимости от конкретного эволюционного пути. Можно предположить, что кислород снял ограничения для всех клеток, делая возможной полифилетическую радиацию – независимое усложнение всевозможных бактерий. Но этого мы не наблюдаем.
Я расскажу об этом подробнее: важно в этом разобраться. Если сложные клетки образовались под давлением классического естественного отбора (генетические мутации обеспечивают разнообразие форм, а те попадают под действие отбора), то стоило бы ожидать множества вариантов внутренней структуры – столько же, сколько существует вариантов внешнего облика клеток. Эукариотические клетки демонстрируют удивительное разнообразие форм и размеров – от гигантских листовидных клеток водорослей до веретенообразных нейронов и растекающихся амеб. Если эукариоты приобрели большую часть сложности в рамках адаптации к конкретным способам существования в расходящихся популяциях, это должно было проявиться в виде соответствующих различий во внутренней структуре. Но заглянув внутрь клетки, мы увидим, что эукариоты, по сути, сложены из одинаковых компонентов. Большинство из нас не сможет отличить растительную клетку от клетки печени или пойманного в ближайшем пруду протиста, даже рассмотрев их под электронным микроскопом: все они поразительно похожи. Попробуйте-ка сами (рис. 3). Если увеличение концентрации кислорода устранило преграды на пути к сложности, то, согласно классической теории естественного отбора, приспособление популяций к различным способам существования должно было привести к полифилетической радиации. Однако ничего подобного мы не наблюдаем.
В конце 60-х годов Линн Маргулис заявила, что происхождение эукариотических клеток путем классического естественного отбора – это заблуждение и эукариоты появились в результате серии эндосимбиозов, в рамках которых бактерии вступали в настолько тесную кооперацию, что некоторые даже физически проникали внутрь партнеров. Эта идея восходит к началу XX века. Рихард Альтман, Константин Мережковский, Жорж Портье, Иван Валлин и другие ученые считали, что сложные клетки появились в результате симбиоза более простых. Их идеи были услышаны, однако считались “слишком фантастичными, чтобы обсуждать их в хорошем обществе”. К началу молекулярно-биологической революции 60-х годов Линн Маргулис еще прочнее утвердилась в своих взглядах, хотя почва под ее ногами оставалась зыбкой. Сейчас мы знаем, что минимум два компонента эукариотической клетки произошли от эндосимбиотических бактерий: митохондрии (преобразователи энергии в сложных клетках) – потомки альфа-протеобактерий, и хлоропласты (фотосинтетические машины растений), произошедшие от цианобактерий. Почти все специализированные органеллы эукариотической клетки хоть раз рассматривались в качестве эндосимбионтов, включая ядро, реснички, жгутик (извивающийся отросток, ритмичное биение которого направляет движение клетки) и пероксисомы (“фабрики токсичного производства” клетки). Таким образом, теория последовательных эндосимбиозов гласит, что эукариоты являют собой плод совместных усилий бактерий, трудившихся сотни миллионов лет, со времени Великого кислородного события.

Рис. 3. Сложность эукариот.
Четыре эукариотические клетки с равной морфологической сложностью.
А. Клетка животного (плазмоцит) с крупным ядром (Я), протяженными внутренними мембранами (эндоплазматический ретикулум, ЭР), на которых располагаются рибосомы, и митохондриями (M).
Б. Одноклеточная водоросль Euglena. Видны хлоропласты (Х), ядро (Я) и митохондрии (М).
В. Растительная клетка с клеточной стенкой, вакуолью (В), хлоропластами (Х), ядром (Я) и митохондриями (М).
Г. Зооспора хитридиомицета, виновного в исчезновении 150 видов лягушек: ядро (Я), митохондрии (М), жгутик (Ж) и гамма-тельца (ГТ), функция которых неясна.
Это поэтический взгляд, но теория последовательных эндосимбиозов в неявном виде подразумевает то же, что и классический отбор. Если бы она была верна, можно было бы ожидать независимого появления множества вариантов внутренней структуры, столь же многообразных, как внешний вид клеток. Рассмотрение любой серии эндосимбиозов, где сотрудничество основано на обмене метаболитами в определенной среде, наводит на мысль, что в разных средах должны реализоваться разные типы взаимодействия клеток. Гипотеза предсказывает, что если такие клетки становятся органеллами сложных эукариотических клеток, у разных эукариот должны появиться разные наборы компонентов внутренней структуры. Тогда в местах вроде стоячих илистых водоемов таились бы организмы всевозможных промежуточных форм, совсем не родственные друг другу, воплощающие разные варианты развития. До самой смерти в 2011 году Маргулис была твердо убеждена, что эукариоты – это мозаика, собранная из более простых организмов в результате цепи эндосимбиозов. Она видела в эндосимбиозе путь жизни, неизведанный “женственный” путь эволюции, на котором процветало “налаживание связей”, – кооперация, которая оказалась гораздо важнее, чем соперничество хищника и жертвы, по сути маскулинное. Но, при всем благоговении перед живыми клетками, Маргулис все же возвратилась к более сухой вычислительной филогенетике – изучению генных последовательностей и целых геномов, которое может рассказать, какие именно родственные отношения связывают разные группы эукариот. А это другая, гораздо более захватывающая история.
Эта история разворачивается вокруг крупной (более 1 тыс. видов) группы простых одноклеточных эукариот, у которых нет митохондрий. Эту группу когда-то считали примитивным “недостающим звеном” между бактериями и более сложными эукариотами, а именно – той самой промежуточной формой, существование которой предполагает теория серийных эндосимбиозов. К этой группе относится, например, пренеприятный кишечный паразит лямблия (Giardia). По словам Эдмунда Юна, она напоминает злобную слезинку (рис. 4). Ее образ жизни вполне соответствует внешнему виду: лямблия вызывает диарею. У нее не одно ядро, а целых два: казалось бы, не должно возникать сомнений в ее “эукариотичности”. Однако у лямблии нет некоторых других базовых эукариотических свойств (особенно заметно отсутствие митохондрий). В середине 80-х годов известный сокрушитель устоев Томас Кавалье-Смит утверждал, что лямблия и другие относительно простые эукариоты появились в самый ранний период эукариотической эволюции, еще до митохондрий, и сохранились в таком виде до наших дней. Хотя Кавалье-Смит и принял тот факт, что митохондрии произошли от бактериальных эндосимбионтов, у него не нашлось времени на детальное изучение теории серийных эндосимбиозов Маргулис. Вместо этого он представил (и до сих пор держится этого представления) самых древних эукариот в виде примитивных фагоцитов, вроде современных амеб, которые питаются, заглатывая другие клетки. Кавалье-Смит утверждает, что клетки, которые приобрели митохондрии, уже имели ядро, динамический внутренний скелет (который позволял им изменять форму и передвигаться), белковые механизмы (позволяющие загружать пищевые частицы внутрь клетки и перемещать их внутри), специализированные компартменты для внутриклеточного переваривания пищи и т. д. Приобретение митохондрий определенно пошло этим клеткам на пользу. Если представить, что клетка сродни автомобилю, то это нововведение сопоставимо с установкой турбокомпрессора. Но даже если машина начинает ездить быстрее, ее устройство в основе остается прежним: двигатель, коробка передач, тормоза, турбина лишь придает энергии. То же самое и примитивные фагоциты Кавалье-Смита. Все части механизма уже были на месте, за исключением митохондрий, которые снабдили клетку большим количеством энергии. Если и существует взгляд на происхождение эукариот, который можно назвать каноническим, – вот он.
С легкой руки Кавалье-Смита этих примитивных эукариот стали называть архезоями, чтобы подчеркнуть их предполагаемое древнее происхождение (рис. 4). Поскольку некоторые архезои паразитируют на человеке, вызывая болезни, их биохимия и структуры геномов представляют интерес для медицины. Исследования в этой области хорошо финансируются, поэтому мы знаем об этих существах чрезвычайно много. За 20 лет изучения геномных последовательностей и биохимии архезоев стало ясно, что на самом деле они вовсе не промежуточное звено между бактериями и эукариотами. Более того, они сами произошли от сложных эукариот, которые некогда обладали всеми компонентами эукариотических клеток, в том числе митохондриями. При переходе к паразитизму архезои утратили эту сложность. Но все они сохранили структуры, которые, как сейчас известно, являют собой упростившиеся митохондрии: гидрогеносомы или митосомы. Внешне эти структуры не похожи на митохондрии (правда, и те, и другие покрыты двумя мембранами, поэтому считалось, что митохондрий у архезоев нет), но из совокупности молекулярных и филогенетических данных точно следует, что гидрогеносомы и митосомы произошли от митохондрий, а не от других бактериальных эндосимбионтов (как считала Маргулис). Таким образом, у всех эукариот имеются либо митохондрии, либо произошедшие от них органеллы. Напрашивается вывод: митохондрии имелись уже у последнего общего предка эукариот, как в 1998 году предсказал Билл Мартин (гл. 1). Тот факт, что у эукариот имеются митохондрии, может показаться тривиальным, но в сочетании со знанием о том, как распространяются геномные последовательности по огромному микробному миру, он когда-то перевернул наше понимание эукариотической эволюции.
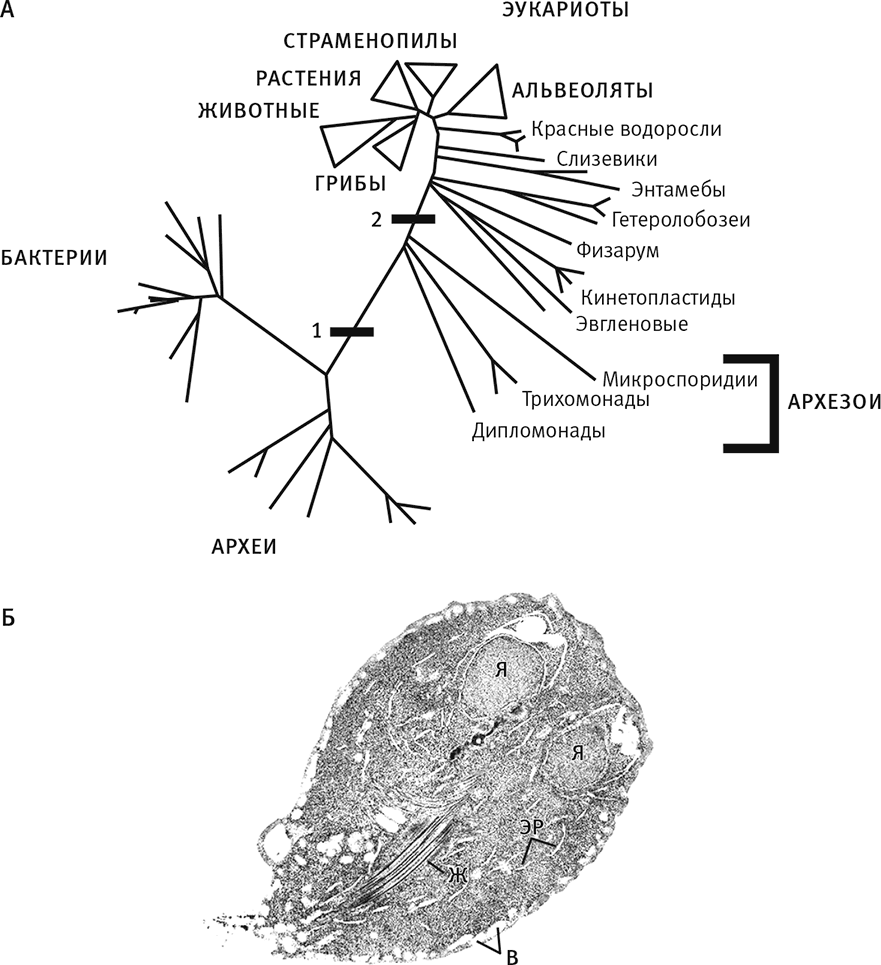
Рис. 4. Архезои – знаменитое якобы недостающее звено.
А. Устаревшая, вводящая в заблуждение филогенетическая схема, построенная на основе сравнения рибосомальной РНК. Она включает три домена: бактерии, археи и эукариоты. Перемычка № 1 обозначает предполагаемое раннее возникновение ядра, а № 2 – предполагаемое приобретение митохондрий, произошедшее позднее. Три группы, ответвившиеся в промежутке между полосками – это архезои, считавшиеся примитивными эукариотами, которые еще не приобрели митохондрии (к ним относится, например, лямблия).
Б. Теперь мы знаем, что архезои вовсе не примитивны: они произошли от организмов, у которых уже имелись митохондрии. В действительности ответвление архезоев произошло от главной части эукариотического дерева. (Я – ядро, ЭР – эндоплазматический ретикулум; В – вакуоль; Ж – жгутик.)
Известно, что все эукариоты восходят к общему предку, который лишь однажды возник в ходе эволюции. Абсолютно все эукариоты (растения, животные, водоросли, грибы, протисты) восходят к общему предку. Иными словами, это монофилетическая группа. Растения, животные и грибы произошли не от разных групп бактерий, а от одной популяции эукариотических клеток со сложной морфологией, единожды появившихся в истории Земли. Общий предок по определению может быть лишь один. В нашем случае под общим предком подразумевается не одна клетка, а одна популяция идентичных клеток. Само по себе это не означает, что появление сложных клеток – очень редкое событие. Вполне возможно, они возникали несколько раз, но выжили потомки лишь одной группы, а все остальные по каким-либо причинам вымерли. Я докажу, что к эукариотам это не относится, но сначала мы должны рассмотреть строение эукариотических клеток.
От общего предка эукариот довольно скоро отделилось пять “супергрупп” организмов с разной клеточной морфологией. Большинство этих групп неизвестны даже людям с классическим биологическим образованием. Примеры таких групп – Unikonta (к ним относятся животные и грибы), Excavata, Chromalveolata и Plantae (последние включают наземные растения и водоросли). Важны два момента. Во-первых, генетическое разнообразие внутри каждой группы гораздо выше, чем между предками этих групп (рис. 5). Это наводит на мысль, что в момент формирования эукариотических супергрупп происходила “взрывная” эволюция, точнее – монофилетическая радиация, которая могла быть связана с преодолением структурных ограничений. Во-вторых, общий предок эукариот уже был чрезвычайно сложной клеткой. Сравнивая признаки каждой супергруппы эукариот, можно попытаться восстановить облик общего предка. Признаки, присутствующие почти у всех видов внутри всех супергрупп, вероятнее всего, унаследованы от него. Признаки, которые встречаются лишь в одной-двух группах, скорее всего, приобретены ими независимо и позднее. Хорошая иллюстрация последнего варианта – хлоропласты, которые встречаются только у растений и хромальвеолят. Они возникли в результате эндосимбиоза, но у общего предка эукариот их не было.
Филогенетика говорит нечто поразительное о том, что было внутри клетки общего предка: было все, кроме хлоропластов. С вашего позволения, я кратко освещу несколько моментов. Мы знаем, что общий предок имел ядро, где хранилась ДНК. Ядро обладало сложной структурой, которая затем стала общей для всех эукариот. Оно окружено двойной мембраной, даже целым рядом уплощенных полостей, которые переходят в другие клеточные мембраны. Ядерная мембрана пронизана замысловатыми белковыми порами и покрыта эластичной сетью, которая поддерживает форму ядра. Внутри ядра присутствуют структуры, универсальные для эукариот, например ядрышко. Стоит подчеркнуть, что множество ключевых белков этих комплексов консервативны – они почти не отличаются у разных супергрупп организмов. Таковы, например, гистоновые белки, на которые накручиваются молекулы ДНК. Все эукариоты имеют линейные хромосомы с колпачками теломеров на концах, которые не дают хромосомам “растрепаться”, как случается с кончиками шнурков. Гены эукариот имеют фрагментарное строение: короткие участки кодирующей ДНК перемежаются длинными некодирующими последовательностями – интронами. Интроны вырезаются перед тем, как с РНК считывается белок. Удаление этих фрагментов осуществляется с помощью механизма, общего для эукариот. Даже расположение интронов нередко консервативно: вставки обнаруживаются на одном и том же месте внутри последовательности общего для разных групп эукариот гена.
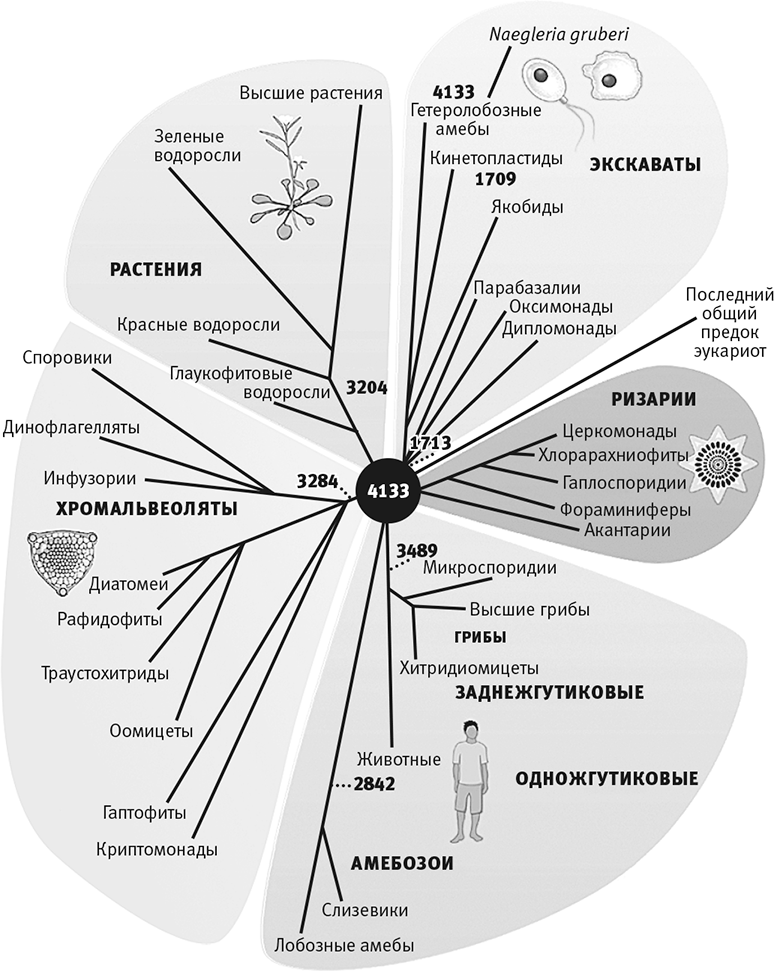
Рис. 5. “Супергруппы” эукариот.
Дерево эукариот, построенное на основе сравнения тысяч общих генов, включает пять “супергрупп”, которые были описаны Евгением Куниным в 2010 году. Цифры означают число генов, общих для данной группы и для Последнего общего предка эукариот. Каждая группа независимо потеряла или приобрела множество других генов. Сильнее всего здесь различаются одноклеточные протисты. Все животные попадают в группу метазои. Заметьте, что различия между представителями одной супергруппы гораздо заметнее, чем между предками эти групп, что позволяет предположить их интенсивное эволюционирование на ранних этапах. Мне нравится символическая черная дыра в центре: Последний общий предок эукариот уже обладал всеми характерными для эукариот свойствами, но филогенетика мало что может сказать о том, как бактерии или археи передали ему эти свойства.
За пределами ядра ситуация сходная. Эукариотам свойственна, по сути, одинаковая клеточная машинерия – за исключением архезоев (разбросанных по всем пяти супергруппам, что свидетельствует о независимой утрате присущей им прежде сложности). У всех эукариот имеются сложные внутренние мембранные структуры, например эндоплазматический ретикулум и аппарат Гольджи, приспособленные для упаковки и выведения наружу белков. У всех эукариот есть динамический внутренний цитоскелет, способный принимать любую форму в соответствии с потребностями клетки. У всех эукариот есть моторные белки. Все эукариоты обладают митохондриями, лизосомами, пероксисомами, транспортными системами для перемещения веществ внутрь и наружу, а также общими сигнальными системами. И список на этом не заканчивается! Все эукариоты делятся митозом, в ходе которого хромосомы расходятся к полюсам клетки, увлекаемые микротрубочками веретена. Набор участвующих в митозе ферментов один и тот же. Эукариоты размножаются половым путем, а их жизненный цикл включает мейоз (редукционное деление), в результате которого формируются гаметы – сперматозоиды и яйцеклетки, которые впоследствии сливаются друг с другом. Те редкие эукариоты, которые отказываются от полового размножения, как правило, быстро вымирают (“быстро” в данном случае – за несколько миллионов лет).
Почти все это давно известно из исследований микроскопической структуры клеток, но наступление новой эры филогеномики прояснило два момента. Во-первых, стало понятно, что перечисленные структурные гомологии обусловлены не поверхностным сомнительным сходством. Все эти признаки закодированы в последовательностях генов миллиардами нуклеотидов, и по этим последовательностям можно выстраивать очень точные филогенетические деревья. Во-вторых, с развитием методов высокопроизводительного секвенирования значительно упростился процесс обнаружения и исследования новых организмов. Отпала необходимость в таких долгих и трудоемких процедурах, как культивация клеток и приготовление микропрепаратов. Новый метод надежен и скор. Так было открыто несколько необычных групп эукариот, например экстремофилы, способные переносить высокие концентрации ядовитых металлов, и пикоэукариоты – крошечные, размером с бактерии, клетки, обладающие полным эукариотическим набором: ядром и митохондриями. Так расширилось наше представление о разнообразии эукариот. Все недавно обнаруженные эукариоты относятся к пяти супергруппам, существование которых четко установлено: на филогенетическом дереве эукариот уже не вырастает крупных ветвей. Поразительно, что при всем разнообразии эукариоты похожи друг на друга. Нам не удается обнаружить ни переходных форм в эволюции эукариот, ни боковых ветвей. То, о чем говорила теория серийных эндосимбиозов, не сбывается.
Это порождает другую проблему. Успехи филогенетики и биоинформатики столь ошеломительны, что мы легко забываем: эти методы имеют свои ограничения. Проблему являет собой филогенетический горизонт событий, скрывающий момент появления эукариот. Ведь все их геномы восходят к Последнему общему предку эукариот (LECA), строение которого уже было очень сложным. Но откуда произошли все его структуры? Есть ощущение, будто предок эукариот родился уже зрелым, как Афина в полном вооружении из головы Зевса. О происхождении почти всех компонентов эукариотических клеток не известно почти ничего. Как и почему появилось ядро? Как возникло половое размножение? Почему у эукариот два пола? Как появилась изумительная система внутренних мембран? Как цитоскелет приобрел пластичность и динамику? Почему при мейозе число хромосом удваивается перед тем, как уменьшиться вдвое? Почему мы стареем, болеем раковыми заболеваниями и умираем? Увы, молекулярная филогенетика, несмотря на свою изощренность, почти ничего не может рассказать об этих биологических процессах. Подавляющее большинство генов, управляющих этими процессами, присуще лишь эукариотам и у прокариот не встречается. У бактерий, в свою очередь, почти не наблюдается тенденции к приобретению сложных эукариотических черт (рис. 6). История появления этих признаков – тайна.
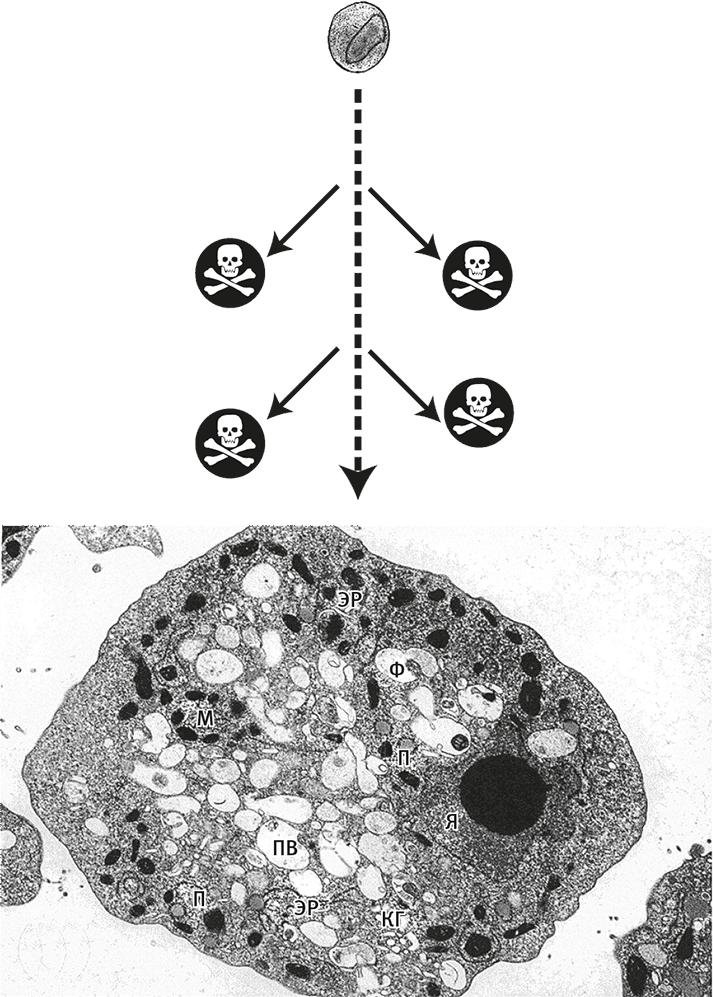
Рис. 6. Грандиозный пробел в знаниях.
Нижний рисунок – электронная микрофотография клетки амебы Naegleria (организма, напоминающего сложностью и размерами Последнего общего предка эукариот). У этой клетки есть ядро (Я), эндоплазматический ретикулум (ЭР), комплекс Гольджи (КГ), митохондрии (М), пищеварительная вакуоль (ПВ), фагосомы (Ф) и пероксисомы (П). Вверху, для сравнения, – электронная микрофотография сравнительно сложной бактерии планктомицета. Разумеется, эукариоты происходят не от планктомицетов, но на этом примере видно, как велика пропасть между самыми сложными из прокариот и примитивнейшими эукариотами. И нам не известны выжившие промежуточные формы, которые могли бы перебросить мост через эту пропасть (возможные переходные звенья обозначены черепом с костями).
Недостающие шаги на пути к сложности
Эволюционная теория предполагает: сложные свойства приобретаются в результате ряда маленьких шагов, и каждый шаг позволяет получить небольшое преимущество. Отбор адаптивных свойств предполагает потерю менее адаптивных, поэтому промежуточные формы непрерывно отсеиваются. С течением времени высота пиков адаптивного ландшафта меняется, и пики, соответствующие самым полезным приобретениям, заслоняют соседние. Так, мы знаем глаз во всем его совершенстве, а менее совершенные промежуточные этапы его развития нам неизвестны. Дарвин в “Происхождении видов” высказал мысль: естественный отбор действительно предполагает утрату промежуточных форм. Поэтому неудивительно, что не сохранилось живых переходных форм между бактериями и эукариотами. Куда удивительнее то, что удачные приспособления не продолжают улучшаться – как в случае глаза.
Мы не можем непосредственно наблюдать этапы эволюции глаза, зато можем видеть экологический спектр его форм у разных организмов. Из зачаточного светочувствительного пятна древнего червеобразного существа глаза независимо появлялись внутри разных групп. Как раз это предсказывает теория естественного отбора. Всякое небольшое изменение обеспечивает небольшое преимущество, которое зависит от среды. В различных условиях появлялись различные по морфологии типы глаз. Некоторые различаются очень сильно (например сложные глаза мухи и зеркальные глаза гребешка), а некоторые схожи – как глаза осьминога и человека, которые в результате конвергенции приобрели почти одинаковое строение. У животных можно найти все мыслимые промежуточные стадии, от зрительных ямок до аккомодационных линз. Даже у протистов иногда встречаются миниатюрные глаза с “хрусталиком” и “сетчаткой”. Эволюционная теория предсказывает, что свойства у разных групп организмов должны были развиваться независимо (полифилетически), и каждый следующий шаг предоставлял новое преимущество. Теоретически этот принцип применим ко всем свойствам – и обычно так и происходит. Так, способность летать при помощи крыльев независимо появлялась минимум 6 раз: у летучих мышей, птиц, птерозавров и разнообразных насекомых. Многоклеточность возникала около 30 раз. Различные формы эндотермности (теплокровности) появились у нескольких групп, включая млекопитающих и птиц, а также некоторых рыб, насекомых и даже, в некотором смысле, у растений. “Сознательную осведомленность” приобрели в большей или меньшей степени независимо птицы и млекопитающие. На примере глаз мы наблюдаем мириады форм, соответствующих условиям, в которых они развивались. Разумеется, существуют физические ограничения, но они недостаточно жестки для того, чтобы исключить многократное возникновение признаков.
А что можно сказать о половом размножении, ядре и фагоцитозе? В этих случаях следует рассуждать таким же образом. Если каждое из этих приобретений появилось в результате действия естественного отбора (несомненно, так оно и было), то мы должны наблюдать многократное возникновение эукариотических признаков у бактерий. Но это не так. Недалеко и до эволюционного “скандала”! У бактерий есть лишь зачатки эукариотических черт. Например, секс. Можно возразить, что бактерии практикуют конъюгацию, а это, по сути, то же самое: передача собственной ДНК партнеру. Правда, в этом случае передача происходит путем горизонтального переноса. У бактерий есть все необходимые механизмы для рекомбинации ДНК, что позволяет им формировать разнообразные новые хромосомы, а это считается главным преимуществом полового размножения. Но все же у этих процессов множество существенных различий. Секс предполагает слияние двух гамет, каждая из которых содержит половину набора генов. Затем происходит их взаимная рекомбинация и объединение в новый целый геном. Горизонтальный перенос происходит редко, случайным образом, задействуя лишь небольшую часть генома и далеко не всегда предполагая взаимный обмен. Если коротко, то эукариоты занимаются сексом по-настоящему, а у бактерий смутное его подобие. Понятно, что при этом у бактерий должно иметься преимущество перед сластолюбивыми эукариотами. Однако тогда можно ожидать, что бактерии по крайней мере некоторых типов делают такое, пусть и отличающееся в деталях. Но нет: насколько известно, они никогда этим не занимались. Так же дело обстоит с ядром, фагоцитозом и почти всеми эукариотическими признаками. Первые шаги на пути к сложности сделаны. Есть бактерии со складчатыми внутренними мембранами. Есть бактерии без клеточной стенки и с более или менее подвижным цитоскелетом. Есть бактерии с линейными хромосомами, с множеством копий генома, с клетками гигантского размера. Все это намеки на эукариотическую сложность. Но бактерии всегда останавливаются, не достигнув уровня эукариот, и очень редко (если такое вообще бывает) одна бактериальная клетка сочетает одновременно несколько эукариотических черт. Самое простое объяснение глубоких различий между бактериями и эукариотами – конкуренция. Утверждается, что, когда появились первые эукариоты, они были настолько конкурентоспособными, что заняли нишу морфологически сложных организмов и никто не мог с ними соревноваться. Если какие-нибудь бактерии и предпринимали робкие попытки посягнуть на указанную нишу, прочно обосновавшимся там эукариотам это было безразлично. То есть бактерии не представляли для них серьезной угрозы. Все мы знаем о массовом вымирании динозавров и других крупных животных и растений, и в этом свете объяснение выглядит вполне правдоподобным. Миллионы лет динозавры притесняли предков современных млекопитающих – мелких пушистых зверьков. Лишь после вымирания динозавров эти зверьки дали начало современным группам млекопитающих. Тем не менее есть веские причины подвергнуть сомнению эту удобную, но обманчивую в своей привлекательности идею. Микроорганизмы – далеко не то же самое, что крупные животные: численность их популяций несравнимо больше, и они способны путем горизонтального переноса передавать друг другу полезные гены (например гены устойчивости к антибиотикам), что сильно снижает угрозу вымирания. Не известно ни одного массового вымирания микробов. Этого не произошло даже после Великого кислородного события. Нет доказательств того, что “кислородный холокост”, в ходе которого, как предполагают, погибло большинство анаэробных клеток, действительно случился: это не подтверждается ни филогенетическими, ни геохимическими данными. Напротив, анаэробы процветали.
Еще важнее вот что. Существует убедительное свидетельство в пользу того, что промежуточные формы на самом деле не были обречены на вытеснение более сложными эукариотами. Доказательство простое: они до сих пор существуют. Мы уже встречались с ними. Это архезои – большая группа примитивных эукариот, которая когда-то ошибочно считалась “недостающим звеном”. Они не являются истинной промежуточной формой в эволюционном смысле, но в экологическом отношении они действительно в промежуточном положении – занимают ту самую нишу. Эволюционная промежуточная форма – это, например, тиктаалик, рыба с ногами, или покрытый перьями крылатый динозавр археоптерикс. Экологическая промежуточная форма – это не настоящее звено, однако ее возникновение позволяет убедиться в том, что определенная экологическая ниша пригодна для жизни. Белка-летяга не является близким родственником других летающих позвоночных, например летучих мышей, тем более птиц, однако она демонстрирует, что планировать с дерева на дерево можно и без покрытых перьями крыльев. Значит, предположение, что механический полет мог начаться подобным образом, не будет пустой выдумкой. И в этом настоящая ценность архезоев: они – экологическая промежуточная форма, своим существованием подтверждающая, что такой образ жизни возможен.
К архезоям относят более тысячи видов. Эти организмы – эукариоты, которые приспособились к “промежуточной” нише путем упрощения, – а вовсе не бактерии, строение которых немного усложнилось. Позвольте это подчеркнуть. Данная ниша пригодна для жизни, и она многократно заселялась морфологически простыми клетками. Простые клетки не были вытеснены эукариотами. Напротив, они процветали, и именно потому, что пошли по пути упрощения. Статистически (при прочих равных условиях) вероятность захвата этой ниши только простыми эукариотами (а не сложными бактериями) в 1 тыс. случаях составляет 1 на 10300: соотношение, которое могло бы получиться в результате работы генератора бесконечной невероятности Зафода Билброкса. Даже если архезои 20 раз (возьмем самую скромную оценку) возникали независимо и всякий раз возникшая форма порождала множество дочерних видов, вероятность все же остается очень небольшой: один из миллиона. Или выпал тот самый единственный шанс, или же вмешалось что-то кроме статистической вероятности. Наиболее правдоподобное объяснение состоит в том, что в строении эукариот было нечто, способствующее освоению ими “промежуточной” ниши, – а строение бактерий, напротив, отличалось чем-либо таким, что мешало приобрести морфологическую сложность.
Эта идея не выглядит чересчур смелой. Она не противоречит ничему из того, что мы знаем. В этой главе я говорил лишь о бактериях, но существует два домена одноклеточных организмов без ядра, которые поэтому называют прокариотами (“доядерными”). Эти группы – бактерии и археи (не путать с архезоями). Я могу лишь извиниться за путаницу. Научная терминология иногда выглядит так, будто придумана специально для того, чтобы никто ничего не понял. Запомните: археи и бактерии – это прокариоты и не имеют ядра, а архезои – примитивные эукариоты, у которых имеется ядро. На самом деле архей до сих пор иногда называют архебактериями – “древними бактериями”, в противоположность эубактериям, “настоящим бактериям”, так что представителей обеих групп можно вполне законно называть бактериями. Для простоты я продолжу использовать термин “бактерии” для обозначения обеих этих групп, за исключением случаев, когда придется подчеркнуть важные различия двух доменов.
Указанные два домена – бактерии и археи – в высшей степени различны в отношении генетики и биохимии, однако морфологически почти не отличаются друг от друга. Организмы, принадлежащие к обеим группам – это маленькие простые клетки, не имеющие ядра и многих других эукариотических черт, характеризующих “сложную” жизнь. Ни одной из этих групп, несмотря на поразительное генетическое разнообразие и биохимическую пластичность, не удалось выработать сложную морфологию. Это обстоятельство можно расценивать так, будто существует некий внутренний физический запрет, не позволяющий прокариотам достичь сложности, а эукариоты его каким-либо образом обошли. В гл. 5 я покажу, что эта преграда была устранена в результате редчайшего события: единичного эндосимбиоза двух прокариот. А пока отметим, что некий запрет вынуждал оба домена прокариот (и бактерий, и архей) оставаться простыми в морфологическом отношении непостижимо долгие 4 млрд лет. Только эукариотам удалось открыть мир сложности, а вступили они в него в результате стремительного лавинообразного развития – монофилетической радиации. Она подразумевает, что эукариоты освободились от всех структурных ограничений, которые только могли им мешать. Судя по всему, это случилось лишь однажды: все эукариоты родственны друг другу.
Неверный вопрос
Это и есть новый взгляд на историю развития жизни. Здесь она изложена очень кратко. Древняя Земля не слишком отличалась от нашей: она была покрыта водой, обладала умеренным климатом и атмосферой, состоящей главным образом из азота и углекислого газа. В то время в составе атмосферы было мало подходящих для органической химии газов, например водорода, метана и аммиака. Это исключает идею “первичного бульона”. Так или иначе, жизнь возникла настолько рано, насколько это вообще возможно. Бактерии на протяжении 2 млрд лет изменяли облик земного шара, преобразуя океаны, атмосферу и континенты. Они становились причиной климатических катастроф – Земля раз за разом покрывалась льдами, а затем оттаивала вновь. Они отравили мир, наполнив химически активным кислородом атмосферу и океаны. И все же за все это время ни бактерии, ни археи не превратились во что-либо иное – они упрямо сохраняли простоту своего строения и образа жизни. В эти бесконечные 4 млрд лет, полные перемен, происходивших и с условиями, и со средой, бактерии изменяли свои гены и биохимию, но никогда – свою форму. Они так и не дали начало более сложным формам жизни – кроме одного-единственного раза, – таким, которые мы надеемся найти на других планетах.
Единственный раз, здесь, на Земле, бактерии дали начало эукариотам. Нет ни палеонтологических, ни филогенетических данных, указывающих на то, что сложная жизнь возникала неоднократно и выжила лишь одна группа: эукариоты. Напротив, то, что эукариоты – монофилетическая группа, наводит на мысль, что их появление было продиктовано необходимостью избавиться от внутренних физических ограничений, которые никак не помогали в периоды экологических потрясений, например во время Великого кислородного события. В части III этой книги мы увидим, какими могли быть ограничения. А сейчас заметим, что любой заслуживающий внимания рассказ на эту тему должен содержать объяснение, почему сложная жизнь возникла лишь однажды. Наше объяснение достаточно убедительно для того, чтобы в него можно было поверить – но недостаточно убедительно для того, чтобы перестать задаваться вопросом, почему это не происходило многократно. Любая попытка объяснить причину уникального события упирается в счастливую случайность. Как мы можем подтвердить то или иное предположение? Само событие мы, конечно, не можем наблюдать непосредственно, но оно могло оставить следы. Как только оковы “бактериальности” были сброшены, эукариоты приобрели удивительную сложность и морфологическое многообразие. Путь наращивания сложности вовсе не был прямым и простым: эукариоты приобрели целый ряд черт (от полового размножения и старения до видообразования), ни одна из которых не встречалась у бактерий или архей. Все эти древнейшие эукариотические черты объединились в единственном и неповторимом общем предке. Мы не знаем переходных форм между морфологической простотой бактерий и чрезвычайной сложностью общего предка, никаких существ, которые могли бы заполнить эту пропасть. Это открывает перед нами захватывающие перспективы: главные вопросы биологии еще предстоит решить! Содержат ли эти черты нечто, что позволило бы понять, как они эволюционировали? Я думаю, да.
Эта загадка возвращает нас к вопросу, поставленному в начале главы. В какой степени, исходя из базовых принципов, можно предсказать свойства жизни и ход ее истории? Я предположил, что жизнь подчиняется таким ограничивающим факторам, которые непросто объяснить с точки зрения генетики, истории и экологии. Идею рассматривать жизнь исключительно в информационном ключе я не поддерживаю, так как этот взгляд не позволяет предсказать ни единого эпизода этой непостижимой истории. Почему жизнь зародилась так рано? Почему она на миллиарды лет застыла на одном уровне морфологической сложности? Почему изменения условий и среды столь мало повлияли на бактерий и архей? Почему сложная жизнь за 4 млрд лет появилась лишь однажды? Почему прокариоты вновь и вновь не порождают новые, более сложные клетки и организмы? Почему этого даже иногда не происходит? Почему свойственные эукариотам специфические черты, например половое размножение, наличие ядра и способность к фагоцитозу, не возникают у бактерий или архей? Почему эукариоты обладают всеми этими чертами?
Если жизнь сводится к информации, то эти вопросы – глубочайшие тайны. Я не верю, что на них можно найти ответ, опираясь исключительно на информацию. Возникновение причудливых особенностей жизни пришлось бы списать на случайность, дело рук беспечной и безжалостной Фортуны. Мы бы не имели возможности предсказывать свойства живого на других планетах. Все же вышло так, что ДНК, будто бы обещающая дать ответ на любой вопрос, заставила нас забыть о втором центральном принципе, который выдвинул Шредингер: жизнь сопротивляется энтропии. В своей книге “Что такое жизнь?” Шредингер заметил, что, если бы сочинял книгу для физиков, то должен был бы вместо термина “энтропия” использовать другой термин: “свободная энергия”.
Слово “свободная” здесь имеет специфическое значение, которое мы обсудим в следующей главе. Сейчас скажу лишь, что энергия – это как раз то, чего не хватает в этой главе и, как верно заметил Шредингер, в его книге. Ее знаменитое заглавие представляет собой неправильно поставленный вопрос. Если учесть, что жизнь – это процесс, требующий активного участия, то есть энергии, вопрос станет гораздо содержательнее: “Что значит жить?” Но оставим Шредингера в покое. Когда он писал свою книгу, о биологическом потоке энергии было известно очень мало. Сейчас мы знаем, как это устроено, вплоть до атомного уровня. Оказывается, сложные механизмы получения энергии являются общими для всех организмов – так же, как является общим для всех генетический код. Этими механизмами определяются действующие на клетки фундаментальные структурные ограничения. Но мы не знаем ни того, как они возникли, ни того, как именно биологическая энергия ограничивала и направляла развитие жизни. Это и есть вопрос, которому посвящена книга.
Назад: Часть I Проблема
Дальше: Глава 2 Что значит жить?

