Глава 8
Мультирегиональное противостояние
Пока я занимался устройством нового института, а Матиас Крингс изо всех сил пытался прочитать мтДНК из пары-тройки дополнительных неандертальцев, в научном сообществе разгорелась битва вокруг наших результатов по типовому неандертальцу из долины Неандерталь. Адепты так называемой мультирегиональной эволюции наши результаты приняли в штыки; эта гипотеза, в частности, утверждала, что неандертальцы среди прочего являются предками современных европейцев. Напрасно мультирегионалисты так разволновались. Ведь в статье 1997 года мы специально отметили, что, несмотря на очевидные различия митохондриальной ДНК неандертальца и современных людей, неандертальцы могли все же оставить свои гены в наследство современным европейцам – но только ядерные гены. Складывалось такое ощущение, будто критика наших исследований мультирегионалистами – это безнадежное сопротивление загнанного в угол: все наши результаты сходились на том, что, по крайней мере для митохондриального генома, подтверждалась модель “из Африки”, а не модель мультирегиональной эволюции. Да и другие лаборатории, изучающие паттерны генетической вариабельности современного человека, также склонялись в сторону африканского сценария, а не мультирегионального. Например, группа Алана Уилсона с Линдой Виджилант и Марком Стоункингом в 1980- х работала с митохондриальным геномом, и наши заключения прекрасно соотносились с их результатами, так что мы оказались в превосходной компании. И даже больше: с начала работы в Германии они расширили свои исследования и на ядерный геном, и выводы из этих исследований были совершенно очевидны.
Изучение ядерного генома современных людей вел Хенрик Кессманн – наверное, самый талантливый аспирант из всех, кого я знал. Хенрик поступил к нам в лабораторию в 1997- м. Высокий, атлетически сложенный блондин, он очень серьезно относился к работе. Мы оба с удовольствием занимались бегом; бегали в Альпах в районе Мюнхена и особенно любили Хиршберг. (Похоже, этому месту отводилась какая-то особая роль в моей жизни.) После тяжелого подъема бегом по причудливым изгибам горной дороги и неторопливого спуска мы часто подолгу обсуждали научные темы, в особенности генетическую изменчивость у человека. Из работ Алана Уилсона и других мы знали, что изменчивость мтДНК у людей ниже, чем у человекообразных обезьян. Это показывает, что современные люди стартовали от небольшой популяции и тем отличны от обезьян. Но одновременно мы ясно понимали, что небольшой размер мтДНК и ее простое, прямолинейное наследование в материнской линии могут исказить реальную картину генетической истории человека и обезьян. Однако когда Хенрик пришел к нам в лабораторию, новые и быстрые способы секвенирования ДНК уже позволяли изучать куски ядерного генома, а не только митохондриального. Хенрика это заинтересовало – он решил исследовать вариабельность ядерной ДНК обезьян и человека. Встал вопрос, какой фрагмент генома взять в работу.
Нам известны функции только десяти процентов ядерного генома. Эти 10 процентов в основном кодируют белки. Тут индивидуальная вариабельность крайне невысока: подавляющее большинство мутаций в этих генах вредны. А если, положим, ген в прошлом изменился и изменил свою функцию и носитель новой вариации оказался лучше приспособлен и имел больше детей, то такой ген должен распространиться в популяции; в результате разница в геномах так или иначе отразит подобный процесс. Оставшиеся 90 процентов генома не так бдительно охраняются естественным отбором – скорее всего, именно из-за того, что их функции не столь существенны для выживания. А нас интересовали как раз случайные мутации и процесс их накопления по ходу эволюционного времени. Поэтому выбрать предстояло из этих 90 процентов. Мы решили сосредоточиться на определенном участке в 10 тысяч нуклеотидов в Х-хромосоме. В нем, насколько было известно, не содержалось генов белков или чем-то примечательных фрагментов ДНК.
Определив, что мы будем секвенировать, теперь предстояло выбрать кого секвенировать, то есть чьи исследовать гены. Понятно, что выбирать нужно из мужчин, так как у них только одна Х-хромосома, а у женщин две – иными словами, задача сразу облегчается в два раза. Но из каких мужчин, вот ведь вопрос! Многие просто работали с наиболее доступным материалом. Например, множество исследований, особенно в медицине, выполнялись на выборках европейцев. Какой-нибудь наивный пользователь базы данных генетической изменчивости человека может утвердиться во мнении, что у европейцев генетическая вариабельность выше, чем у других групп населения. А на самом-то деле их высокая вариабельность отражает лишь относительно бедную представленность других групп.
Создать более разумную выборку мы могли тремя способами. Во-первых, можно отбирать мужчин пропорционально численности населения в разных частях света. Эту идею пришлось отставить, поскольку образцы в таком случае будут в основном из Китая и Индии, где население сильно выросло за 10 тысяч лет в результате, в частности, развития сельского хозяйства. Другими словами, согласившись на этот вариант, о генетическом разнообразии говорить не придется. Во-вторых, можно собирать образцы, ориентируясь на территорию, то есть, например, по одному образцу с каждых двух-трех квадратных километров. Но это, вдобавок к баснословным транспортным расходам, приведет к чрезмерно раздутой выборке из областей с низкой плотностью населения, например из северного Заполярья. По третьему сценарию, на котором мы в конце концов остановились, выборка производится в соответствии с основными лингвистическими группами. Мы считали, что лингвистические группы (индоевропейская, финно-угорская и т. д.) в общем приближении отражают культурное разнообразие древности, и оно старше, чем 10 тысяч лет. Так что, если мы будем отбирать образцы по лингвистическим группам, у нас увеличатся шансы сделать выборку из групп населения с независимой генетической историей. И мы, хочется верить, получим более полную картину генетической изменчивости.
К счастью, мы были не первыми: образцы ДНК по этому принципу уже собрал известный генетик из Стэнфорда, итальянец Лука Кавалли-Сфорца, и мы могли воспользоваться его коллекцией. Хенрик отобрал шестьдесят девять образцов – мужских представителей основных языковых групп. Для каждого образца отсеквенировал назначенные цепочки из десяти тысяч нуклеотидов. Сравнив затем последовательности ДНК в случайно выбранных парах, он подсчитал, что среднее число отличающихся нуклеотидов равно 3,7. Так же как и в случае с митохондриальной ДНК, Хенрик отметил большую вариабельность ядерной ДНК у пар из Африки, чем у неафриканцев. Чтобы понять смысл полученных результатов, он обратился к исследованию ближайших родственников человека – шимпанзе.
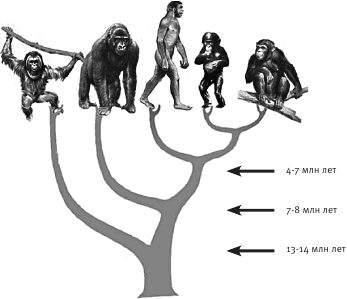
Нам известны два вида шимпанзе, оба из Африки. “Обычные” шимпанзе обитают в экваториальных лесах и саваннах, распределяясь пятнами от Танзании на востоке до Гвинеи на западе. Бонобо, которых иногда называют карликовыми шимпанзе, живут только к югу от реки Конго, в Демократической Республике Конго. Наши эволюционные линии разошлись, как показывает сравнительный анализ ДНК этих видов, примерно 4–7 млн лет назад. А еще раньше, 7–8 млн лет назад, жил общий предок человека, шимпанзе и гориллы, другой африканской человекообразной обезьяны. И все они вместе с орангутанами с Борнео и Суматры произошли от общего предка, существовавшего 12–14 млн лет назад (рис. 8.1).
Хенрик выбрал тридцать самцов шимпанзе (обыкновенного, не бонобо), представляющих популяции Восточной, Центральной и Западной Африки, и секвенировал тот же участок Х-хромосомы, с которым работал на человеческом материале. Он повторил сравнение у случайно выбранных пар. Получилось среднее различие в 13,4 нуклеотида для пары особей. Невероятный результат! Население в семь миллиардов против каких-то двух сотен тысяч шимпанзе… Население, занявшее каждый свободный клочок земли, против кучки особей, уцелевших на африканском экваторе! А генетических различий у любых двух шимпанзе в 3–4 раза больше, чем у двух людей.
Затем Хенрик составил такие же последовательности для бонобо, горилл и орангутанов. Он пытался проверить, люди ли так необычно похожи (генетически) или шимпанзе так необычно различаются. И выяснил, что изменчивость ДНК у горилл и орангутанов даже выше, чем у шимпанзе, и только у бонобо разница в нуклеотидных последовательностях такая же низкая, как у людей. Мы опубликовали результаты исследований в 1999–2001 годах в трех статьях в Nature Genetics и Science. В этих статьях мы продемонстрировали, что изменчивость ядерной ДНК на выбранном участке очень похожа на ту, которую группа Алана Уилсона выявила для мтДНК. По-видимому, этот паттерн изменчивости типичен для всего человеческого генома, и у меня все больше крепло убеждение, что верна как раз модель “из Африки”. А вот критические замечания мультирегионалистов я слушал-слушал, и как-то они меня совсем не впечатляли. Я не отвечал на критику или почти не отвечал: думал, время рассудит.
Армия мультирегионалистов состояла в основном из палеонтологов и археологов. Я бы ни за что не признался вслух, но в душе не очень-то верил, что они в принципе могут решать вопросы о взаимодействии двух групп: например, одна из двух групп заместила другую, перемешалась с ней или одна группа изменилась, превратившись в нечто новое? Эти палеонтологи даже не смогли между собой договориться, как отделить одну архаичную группу от другой. И в результате разгорелись – и поныне бушуют – яростные баталии между “дробителями”, различающими множество ископаемых видов гоминин, и “объединителями”, относящими ископаемые остатки всего к паре-другой видов. Кроме того, у палеонтологии есть и другие, так сказать, врожденные противоречия. Есть знаменитая фраза антрополога Винсента Сарича, работавшего с Аланом Уилсоном в 1980- х. Он сказал, что у ныне живущих людей предки были наверняка, иначе бы они, люди, сейчас не жили, а вот в обратную сторону – неизвестно: были у найденных ископаемых людей потомки или нет. И в самом деле, большинство ископаемых остатков из музеев выглядят вполне “по-человечески”, потому что у нас с ними в глубоком прошлом обязательно найдутся общие предки, но очень часто эти ископаемые люди не имеют прямых современных потомков, то есть они что-то вроде генеалогического тупика на нашем фамильном древе. И тем не менее многие мысленно помещают их в категорию “наших предков”. Временами в приступах мечтательного энтузиазма мне видится, как с помощью ископаемых ДНК мы навсегда избавляемся от этих привычных двусмысленностей.
Среди критиков-мультирегионалистов числился знаменитый палеонтолог Эрик Тринкаус. Он заметил, что в работе с неандертальцами мы, возможно, все время совершаем одну и ту же ошибку, отбрасывая как загрязнения любые цепочки ДНК, напоминающие ДНК современного человека. Он утверждал, что любые из отброшенных фрагментов могут оказаться своими, неандертальскими. И действительно, некоторые костные образцы неандертальцев давали на выходе только последовательности, похожие на современные. Но как раз про такие образцы я точно знал, что их неряшливо хранили, а потому не сомневался, что все эндогенные фрагменты в них давно разложились и к нам в пробирки попадают только привнесенные ДНК. И все же в рассуждениях Тринкауса логика имелась, и я понимал, что к его аргументам нужно отнестись серьезно.
К этой задаче обратился Давид Серр, французский аспирант из Гренобля. Голову его украшала невероятная копна густейших волос, зимой он носился как сумасшедший на горных лыжах, летом бесстрашно скатывался по каньонам с головокружительных водопадов. Мы решили, что если он останется жив после всех своих эскапад, то пусть выяснит, у всех ли неандертальцев митохондриальные ДНК такие же, как и у “типового” неандертальца, и есть ли они у ранних людей современного типа, живших в Европе одновременно с неандертальцами. Вот этот-то последний вопрос и являлся основным. Как уже говорилось, выживание того или иного варианта мтДНК – это дело случая. Можно себе представить, что ранние представители современных людей добрались до Европы, там перемешались с местными жителями, неандертальцами. Тогда некоторые (или даже многие) из них получили неандертальские мтДНК, но не передали их последующим поколениям. Так могло быть, если у мам с неандертальскими мтДНК не рождались дочки. Именно такой сценарий описал шведский биолог-теоретик Магнус Норборг вскоре после нашей публикации в Cell в 1997- м.
Подобные рассуждения ужасно меня раздражали, так как в них некорректно смешивались два разных вопроса. Первый вопрос – внесли ли неандертальцы генетическую лепту, которую можно отследить сегодня в составе мтДНК современного человека. Мы ответили на него отрицательно. И второй вопрос – смешивались ли люди с неандертальцами. На это ответа у нас не было. Тем не менее я считал первый вопрос более интересным и значимым. Мне важно было знать, есть ли у меня или у кого-то из моих современников неандертальская ДНК. Если мы не унаследовали ДНК неандертальцев, то любые межвидовые скрещивания или нескрещивания 30 тысяч лет назад с генетической точки зрения значения не имеют. В разговорах с журналистами я неизменно подчеркивал этот аспект. Чтобы выразиться еще яснее, я говорил, что меня совершенно не интересует сексуальное поведение в позднем плейстоцене, если только это поведение не оставило в нас сегодняшних какие-нибудь генетические следы. Иногда добавлял, что меня бы удивило, если бы те люди не воспользовались возможностью вступать в половые связи с неандертальцами. Но для нас важно только одно: появлялись ли у них после этого дети, чтобы передать родительские гены далеким нам.
Несмотря на раздражение, я поручил Давиду эту задачу: отследить неандертальскую мтДНК у ранних людей из Европы, пусть даже эта ДНК и исчезла из современных геномов. А если древние европейцы в какой-то момент получили неандертальскую мтДНК, то почему бы им не унаследовать от неандертальцев и ядерную ДНК? А это значит, что какие-то фрагменты неандертальской ядерной ДНК могли сохраниться у нас.
Мы разослали письма в несколько европейских музеев с просьбой прислать костный материал по неандертальцам и ранним представителям людей. Наш успех с исследованием “типового” неандертальца несколько смягчил сердца музейных хранителей и позволил набрать образцы костной ткани двадцати четырех неандертальцев и сорока ранних людей современного типа. Давид проанализировал аминокислоты из всех шестидесяти четырех образцов. Судя по степени сохранности, только в образцах четырех неандертальцев и пяти человек можно было предполагать присутствие мтДНК – такова была суровая, хотя и предсказуемая реальность. Давид выделил ДНК из девяти костей и запустил ПЦР, используя праймеры для амплификации мтДНК и человекообразных обезьян, и неандертальцев, и, естественно, людей. Он получил продукты амплификации для всех девяти образцов. Составленные последовательности оказались идентичными современным. Неужели Тринкаус все-таки прав?
Я велел Давиду повторить эксперимент, только теперь взять в оборот костный материал от пятерых пещерных медведей из Виндии и одного из Австрии. И что вы думаете! Амплификация вытяжек костей медведей тоже дала человеческую нуклеотидную последовательность. Мы опять – я еще больше уверился в этом – получали на выходе цепочки человеческой современной ДНК, занесенной при работе с образцами. Давид старательно составил праймеры, которые должны были амплифицировать исключительно неандертальскую или похожую на нее мтДНК, но никак не мтДНК человека. Мы проверили эти праймеры на разнообразных ДНК, а затем опробовали их на пещерных медведях. Ничего. Амплификация не дала ничего. Прекрасно: это значило, что праймеры и впрямь специально предназначены для амплификации неандертальской мтДНК. Он испытал эти праймеры на вытяжках из неандертальских и человеческих костей. Из всех неандертальских костей он получил последовательности, похожие на те, что мы получили из “типового” неандертальца, подтвердив еще раз, что у неандертальцев нет митохондриальных нуклеотидных последовательностей, свойственных современным людям. И напротив, ни один из образцов пяти представителей ранних людей не показал наличие “типовых” неандертальских последовательностей. Похоже, Тринкаус ошибался.
Тем не менее эту тему хорошо было бы развить, и мы обратились к теории. Мы смоделировали популяцию древних людей, в которой они скрещивались с неандертальцами, а затем их потомки просуществовали в течение 30 тысяч лет и дожили до сегодняшнего дня. Затем мы спросили себя, каков мог быть наибольший генетический вклад неандертальцев в генофонд, если ни у ныне живущих, ни у ранних людей неандертальской мтДНК не обнаружено (это мы приняли за данность). По этой модели (весьма упрощенной, так как мы не включали в нее, например, скорость роста современного населения) мт-геном и ядерный геном современного человека не могут содержать больше 25 процентов неандертальской ДНК. Но мы ведь не обнаружили прямых доказательств неандертальского генетического вклада в современный генофонд, значит, резонно допустить, что этого вклада просто не было.
Я считал, что такой результат прекрасно иллюстрирует сильные стороны нашего подхода в противоположность классическому палеонтологическому анализу. Мы пользовались четко сформулированными предпосылками и делали заключения с определенной вероятностью. Никакой морфологический анализ костных фрагментов не мог сравниться с нашим по отточенности и объективности. Многие палеонтологи говорили об объективности и действенности собственных методов, но эти методы почему-то уже два десятка лет не могут дать сколько-нибудь определенный ответ, передали нам неандертальцы гены или нет; так что палеонтологический подход, по всей видимости, имеет серьезные ограничения.
Мы опубликовали результаты работы Давида, и после этого группа теоретиков из Швейцарии, возглавляемая популяционным генетиком Лораном Эскофье, разработала более развернутую модель взаимодействия неандертальцев и представителей ранних людей. Они допустили, что когда люди, анатомически уже достигшие уровня современного человека, мигрировали в глубь Европы, любое смешение с неандертальцами происходило на границе миграционной волны. Для такой первой миграционной волны характерна небольшая численность популяции с быстрым последующим ростом. Швейцарская группа показала, что в соответствии с этой моделью даже редкие случаи скрещивания оставят след в митохондриальном генофонде, так как в среднем в растущей популяции у женщин чаще рождаются дочери, которые и передадут материнскую мтДНК. В этом случае, то есть в случае растущей популяции, у неандертальской мтДНК, попавшей в генофонд человека, меньше вероятность исчезнуть, чем при постоянной численности популяции. Так как мы не смогли обнаружить следов неандертальской мтДНК ни у представленных пятерых ранних людей, ни у тысяч живущих сейчас (по материалам наших и других исследований), группа Эскофье заключила, что все указывает на “практически полную нескрещиваемость неандертальских женщин и сапиентных мужчин и, следовательно, перед нами два разных биологических вида”.
По мне, такой вывод был вполне допустим, хотя, конечно же, оставалась вероятность, что обстоятельства, при которых неандертальцы и люди скрещивались, были какими-то особенными и наши модели не могли их учесть. Например, если все дети от таких смешанных связей оставались жить в неандертальских сообществах, их генетический вклад был бы равен нулю, и картина выглядела бы как результат “практически полной стерильности”, как выразилась швейцарская группа. Или представим себе, что скрещивание происходило между мужчинами-неандертальцами и женщинами-людьми, и тогда подобные события не оставили генетического следа в генофонде мтДНК, так как мужские особи не передают мтДНК своему потомству. Такой процесс оставит след только в ядерном геноме. Так что если мы хотим понять, как встречи наших предков с неандертальцами повлияли на состав нашего с вами генома, нам определенно нужно браться за ядерный геном неандертальцев.

