Лекция шестая
2) Внутреннее торможение: в) запаздывание
Мм. гг.! Перехожу к нашему третьему случаю внутреннего торможения, называемому у нас запаздыванием.
Понятно, что комбинирование условного и безусловного раздражителей при выработке условного рефлекса можно очень разнообразить в отношении промежутка между началом условного раздражителя и моментом присоединения к нему безусловного. Промежутки эти могут быть короткие, 1–5 секунд, может быть, даже и части секунды (но должно, как это уже было указано, непременно начинать с условного раздражителя), а также и более длинные, до нескольких минут. Таким образом, происходят разные условные рефлексы, которые мы отличаем разными названиями: то совпадающие, точнее говоря – почти совпадающие, то отставленные, или запаздывающие. Промежутки играют очень важную роль, так как ими, во-первых, обусловливается то торможение, которым мы теперь будем заниматься; во-вторых, они же определяют окончательную судьбу каждого условного рефлекса, как увидим это позже.
Если промежуток постоянно берется малый, 1–5 секунд, то при выработке условного рефлекса и слюноотделение тоже начинается быстро, сейчас же после начала условного раздражителя. При увеличении промежутка слюноотделение все более и более запаздывает, отодвигается от начала условного раздражителя. Это запаздывание соответственно доходит до минут. Запаздывания можно достигать разными приемами. Один раз мы начинаем опыт с почти совпадающего рефлекса. Условная реакция наступает быстро – через 1–3 секунды. Затем мы постепенно увеличиваем промежуток, положим, каждый день на 5 секунд. Соответственно и условный эффект постепенно запаздывает, пока мы не остановимся на определенном промежутке и таким образом на определенном запаздывании. В другой раз мы сразу переходим с почти совпадающего рефлекса на очень отставленный. Тогда выработанная реакция сначала совершенно исчезает, и на большой срок происходит длинный период нулей, как выражались некоторые мои сотрудники в своих работах. А кончается тем, что поблизости момента присоединения безусловного раздражителя появляется слюноотделение, которое с повторением так расположенного опыта все растет, подаваясь постепенно назад, т. е. к началу условного раздражителя, и останавливаясь, наконец, на значительном от него расстоянии. Начинать прямо с очень значительного отставления едва ли возможно, по крайней мере у огромного большинства собак, потому что собаки в таких случаях скоро погружаются в сонливое состояние, и тогда получить у них условный рефлекс не удается. Из-за указанной трудности этот последний прием не был у нас достаточно исследован. Вот образец запаздывающего рефлекса из работы Завадского, которому принадлежит заслуга особенно тщательного обследования данного случая внутреннего торможения.
У собаки свисток – условный кислотный раздражитель. Начало условного раздражителя от начала безусловного отделено промежутком в 3 минуты.
«Комбинирование условного и безусловного раздражителей при выработке условного рефлекса можно очень разнообразить в отношении промежутка между началом условного раздражителя и моментом присоединения к нему безусловного»
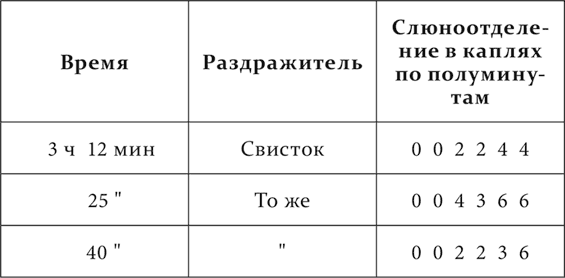
Скорость развития запаздывания чрезвычайно разнообразна. Прежде всего она находится в очень резкой зависимости от индивидуальности животного, характера его нервной системы. У одних собак запаздывание происходит очень быстро, у других, наоборот, очень медленно: слюноотделение упорно не отходит от начала условного раздражителя. У одних оно обозначается в течение одного опыта при повторении нескольких более отставленных рефлексов, у других для этого мало месяца и более. У первых запаздывание часто довольно скоро переходит в сонное состояние, и поэтому у некоторых из этих собак приходится при систематической работе над ними ограничиваться коротко отставленными рефлексами, т. е. довольствоваться меньшей величиной слюноотделения, чем у других. Затем отчетливо дает себя знать род условных раздражителей. Механические и термические раздражения кожи и световые раздражения скорее ведут к развитию запаздывания, чем звуковые. При прочих равных условиях, это идет рядом вообще с меньшим условным эффектом первых раздражений сравнительно со вторыми. Вот опыты из работы В. В. Яковлевой, иллюстрирующие эти отношения.
У собаки три условных пищевых раздражителя: удары метронома, механическое раздражение кожи и вспыхивание лампочки перед глазами.
Опыт 9.IV.1924. Безусловный раздражитель отставлен от условного на 30 секунд.
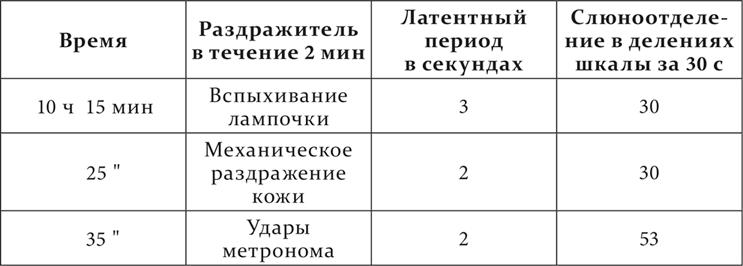
Опыт 24.IV.1925. Рефлексы, многократно применявшиеся для других целей за годовой промежуток между этим и предшествующим опытом, затем были сделаны более отставленными: сперва на минуту, а потом на две. Все рефлексы повторялись отставленными одинаковое число раз.
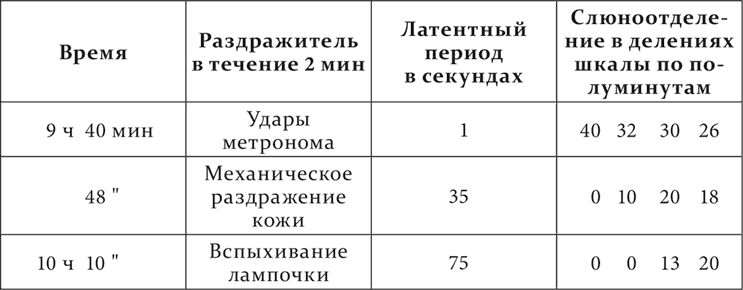
Большое влияние имеет длительная практика короткого отставления, которое иногда упорно не допускает развития более значительного запаздывания при большем отставлении безусловного рефлекса от начала условного.
Наконец, по-видимому, при раздражениях одного рода, например звуковых, запаздывание развивается скорее или медленнее в связи с тем, является ли раздражение сплошным или прерывистым. В случае сплошного – оно наступает скорее.
Итак, при большом отставлении во времени безусловного раздражителя от начала условного эффект условного состоит из двух фаз, часто одинаковых по продолжительности: начальной недеятельной и второй деятельной. Что значит недеятельная фаза? Нарастает ли, постепенно суммируется раздражительный процесс, чтобы через очень большой период проявиться в видимом эффекте, или он, достаточный по силе, существует гораздо раньше, но до поры до времени благодаря чему-то маскируется, заслоняется? Уже обычные приемы при получении запаздывающих рефлексов говорят против первого предположения. При первоначальном коротком отставлении мы уже имеем вполне проявившийся эффект условного раздражения. Почему же тот же раздражитель при большей его продолжительности требовал бы большего суммационного времени? Если бы это было утомлением, то оно должно было бы вести к постепенному уменьшению эффекта. А этого нет. Эффект, хотя и запаздывая, при первом приеме выработки запаздывания (прием приведен в начале лекции) все увеличивается, а при втором – сначала исчезает, потом появляется вновь и, долго возрастая, наконец, становится постоянным на максимуме. Ввиду этого, остается второе предположение, что здесь имеет место замаскирование раздражения, его временное задерживание. И это доказывается фактом, что раздражительный процесс в первой фазе можно быстро выявить.
Если мы в недеятельной фазе подействуем на наше животное каким-нибудь посторонним агентом, не имевшим до сих пор никакого отношения к работе слюнных желез, то мы получим сейчас же слюноотделение, и часто обильное, вместе с соответствующей данному условному раздражителю двигательной реакцией, т. е. условный рефлекс выступает целиком.
Вот опыты Завадского.
Механическое раздражение кожи – условный кислотный раздражитель. Начало безусловного раздражителя от начала продолжающегося все время условного раздражителя отделено промежутком в 3 минуты. Удары метронома никакого отношения к кислотному рефлексу доселе не имели.
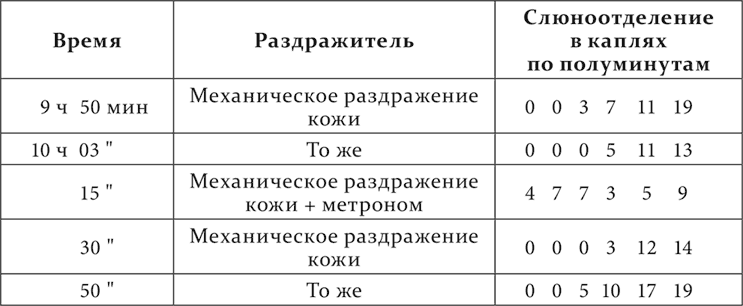
Другой опыт с другим посторонним раздражителем на том же животном; посторонний раздражитель – бесшумно вертящийся предмет.
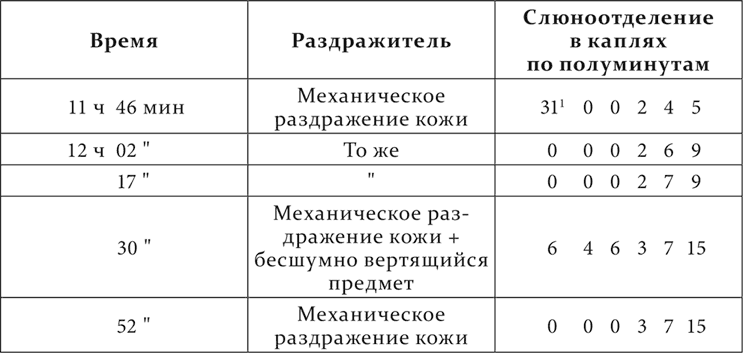
Происходит очень интересное и неожиданное явление: условный выработанный раздражитель один не действует 1–1½ минуты, а индифферентный агент, впервые к нему в это время прибавленный, тотчас же обусловливает закономерную реакцию данного условного раздражителя.
Очевидно, что мы здесь имеем дело с тем же явлением, с которым познакомились раньше и которое было названо растормаживанием. Так как в нашей лаборатории впервые оно наиболее систематически было исследовано Завадским, то я и остановлюсь более подробно на его опытах.
Как вы могли заметить, я в трех лекциях подряд говорю о растормаживании и привожу относительно него много протокольного материала. Оправданием для этого, как мне кажется, должна служить относительная физиологическая своеобразность явления, хотя и для него, конечно, имеются аналогии в теперешней нервной физиологии, и затем полная темнота, что касается понимания ето внутреннего механизма. Можно надеяться, что изобилие фактических подробностей может когда-нибудь пособить в понимании этого механизма.
Обращаю ваше внимание на первый из только что приведенных двух опытов. Мы видим в нем отчетливо, что присоединенные к механическому раздражению кожи удары метронома не только вызвали слюноотделение в недеятельной фазе, но вместе с тем резко уменьшили слюноотделительный эффект во второй, деятельной фазе. Во всех случаях одного механического раздражения кожи за 1½ минуты выделилось слюны от 29 до 46 капель; при раздражении кожи совместно с ударами метронома – 17 капель. Таким образом, мы наблюдаем двойное действие постороннего агента, растормаживающее в недеятельной фазе и тормозящее в деятельной.
«Мы быстро привыкаем к запахам, звукам и освещению, т. е. ощущаем их все иначе. А это значит, что нервные клетки, на которые падают раздражения, переживают различные последовательные состояния. И только определенное состояние, совпадающее с безусловным рефлексом, становится условным раздражителем»
Если присоединить к условному раздражителю в запаздывающем рефлексе различные посторонние агенты из группы внешних тормозов, то получается очень разнообразная картина изменений в течении запаздывающего рефлекса, в которой, однако, легко открывается определенная закономерность. Все посторонние агенты, применяемые в этих наших опытах, могут быть расположены в следующий ряд по их влиянию на запаздывающие рефлексы.
I. Температура 5 °C и 44 °C, приложенная к коже, и слабый запах камфоры.
II. Температура 0,5 °C и 50 °C.
III. Бесшумно вертящийся перед глазами собаки предмет, удары метронома, механическое раздражение необычных мест (если механическое раздражение одного места кожи есть условное раздражение), свисток умеренной силы, запах уксусного амила.
IV. Интенсивный запах камфоры, сильные свистки и звонки.
Первый ряд раздражителей не оказывает никакого влияния на ход запаздывающего рефлекса.
Второй ряд обнаруживает свое действие только на первой фазе рефлекса, вызывая во время нее отделение слюны.
Третий ряд нарушает обе фазы: в первой вызывая слюноотделение, во второй уменьшая его сравнительно с нормой.
Наконец, четвертый ряд почти или совсем не изменяет первую фазу, но вполне или почти подавляет слюноотделение во второй.
Нужно прибавить, что эти опыты при соблюдении определенных предосторожностей идут часто с поражающей точностью.
Вот ряд опытов, иллюстрирующих указанные отношения.
Механическое раздражение кожи – условный кислотный раздражитель. Начало условного раздражителя от начала безусловного отделено промежутком в 3 минуты. Посторонними раздражителями служат: температура в 44 °C, приложенная к коже, температура в 0,5 °C, запах уксусного амила и звонок.
Опыт 13.X.1907
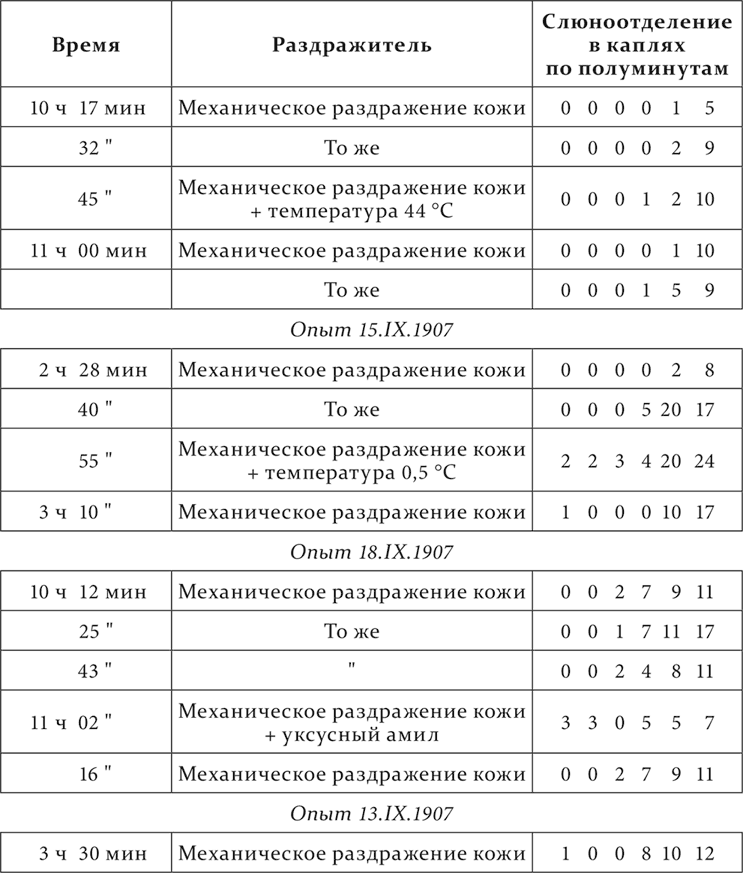
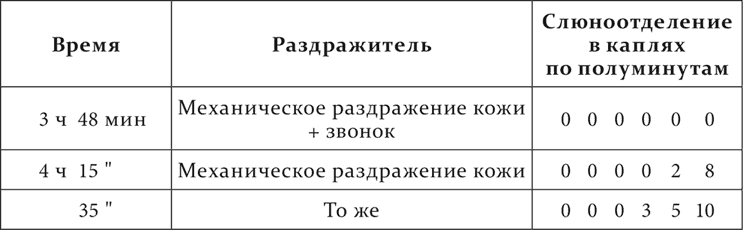
Совершенно те же в существенном результаты получаются, если посторонние раздражители присоединяются к условному раздражителю не на всем его протяжении от его начала до присоединения безусловного раздражителя, а отдельно то в недеятельную фазу (первые 1½ минуты), то в деятельную (вторые 1½ минуты). Вот примеры.
То же животное и те же отношения.
Опыт 23.VII.1907
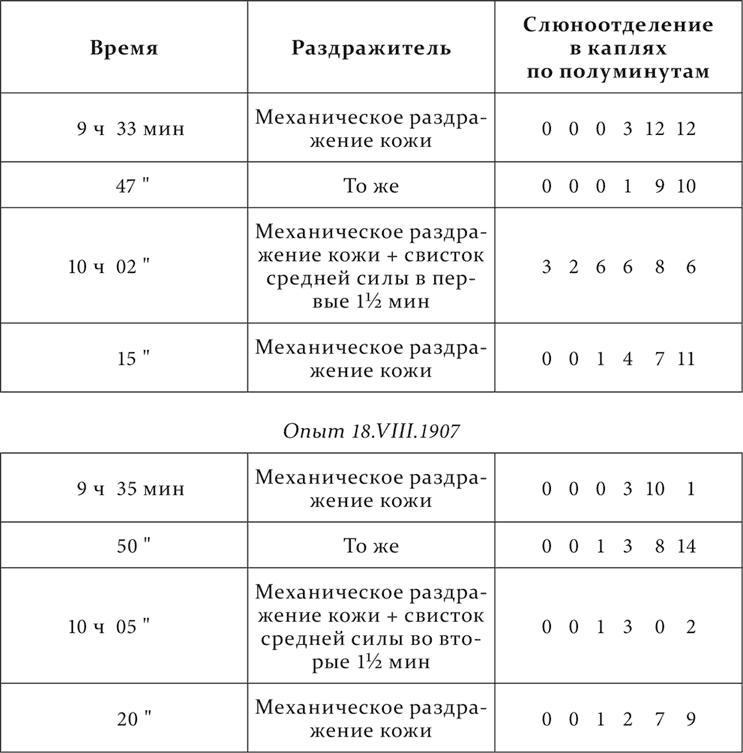
Посторонний раздражитель из третьей группы отдельно вызвал слюноотделение в первой фазе и уменьшил его во второй; в первом опыте некоторое уменьшение его во второй фазе, конечно, произошло от следов раздражения.
«Для цели определения истинного латентного периода условных рефлексов необходимо пользоваться совпадающими рефлексами, когда безусловный раздражитель возможно скоро, через секунду или даже части секунды, следует за началом условного»
Теперь поднимается вопрос: что это за серия посторонних раздражителей? Что лежит в ее основании? По многим данным становится ясным, что здесь идет дело об относительной физиологической силе посторонних раздражителей, о величине реакций на них животного. Приведенный ряд есть ряд все более и более увеличивающихся по силе действия раздражителей. В некоторых случаях это прямо видно в таблице, когда применяются разные интенсивности одних и тех же агентов и когда, стало быть, соответственно изменяются и реакции на них животного. А затем и во всех случаях это обнаруживалось в двигательной реакции животного. При первых членах ряда часто не замечалось ни малейшей реакции. При дальнейших членах ряда реакция становилась постепенно все энергичнее и продолжительнее. Наконец, применяемые нами теперь посторонние раздражители суть агенты так называемого нами внешнего торможения, и на деятельной фазе анализируемых рефлексов вполне отчетливо и тем резче сказывается их тормозящее действие, чем более вниз мы подвигаемся по приведенной таблице, кончая полным торможением. Следовательно, наш ряд посторонних раздражителей есть действительно ряд по нарастающему их действию на организм, начиная с легкого исследовательского рефлекса.
Что это понимание фактов правильно, вполне подтверждается и следующей вариацией опытов. Если посторонний раздражитель повторно присоединять к запаздывающему рефлексу, то его тормозящее действие на деятельную фазу постепенно раз от разу падает. А это есть совершенно понятное свойство посторонних раздражителей, как гаснущих тормозов.
Вот опыты с тем же животным, что выше.
Опыт 13.XI.1907
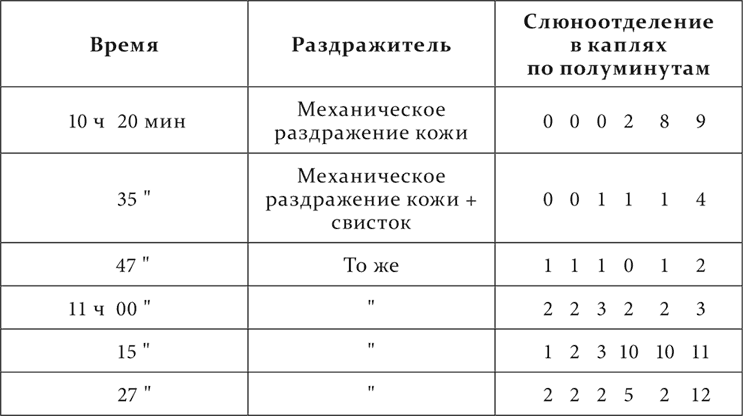
Опыт 20.XI.1907
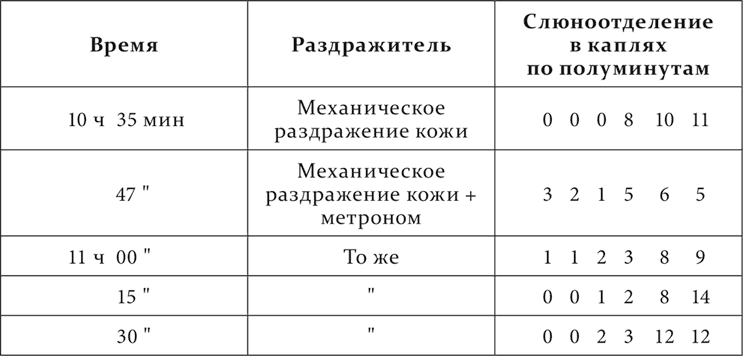
В этих опытах мы видим, как тормозящее действие постороннего раздражителя на деятельную фазу с повторением постепенно падает, особенно правильно во втором опыте. Отношения, выступившие во всех приведенных опытах, проделанных на одном и том же животном, повторились и на других животных. Разница оказалась только в некотором изменении состава рядов посторонних раздражителей, члены которых располагались отчасти в другом порядке, чтó и понятно. Реакция животных на разные посторонние раздражители очень, даже чрезвычайно часто колеблются в интенсивности в зависимости от индивидуального характера нервной системы и от прошлого животного, от его встреч в жизни с теми или другими раздражителями, как это каждый из нас знает хорошо по себе.
«Если с условными тормозными раздражителями одновременно на короткие сроки, но повторно, применять индифферентные агенты, то эти последние приобретают также тормозную функцию, т. е. и сами по себе вызывают в коре процесс торможения»
Таким образом, мы должны признать, что указанный выше порядок изменений эффекта запаздывающего рефлекса под влиянием посторонних раздражителей действительно определяется различной физиологической силой этих раздражителей. При малой силе их незаметно никакого изменения на нашем рефлексе. При некотором увеличении ее терпит изменение только недеятельная фаза, превращаясь в деятельную. При еще более значительном увеличении одновременно изменяется и деятельная фаза, очень теряя в своем размере, часто сравниваясь, а иногда даже и уступая первой фазе, ставшей теперь деятельной. Наконец при высшей силе некоторых посторонних раздражителей запаздывающий рефлекс делается нулевым на всем своем протяжении.
В описанных изменениях запаздывающего рефлекса под влиянием посторонних раздражителей особенно резко подчеркиваются два важных факта, относящихся к деятельности центральной нервной системы. Первый, что посторонний раздражитель, действуя на положительный условный раздражитель, его тормозит, а падая на отрицательный, тормозной, его растормаживает; иначе говоря, в обоих случаях изменяет наличный центральный нервный процесс в обратный. А второй, что тормозной процесс сравнительно с раздражительным процессом лабильнее, так как более легко, под влиянием более слабых посторонних раздражителей, испытывает изменение.
Относительно этого второго пункта у меня имеется следующий случайный, но поучительный опыт. Я читал публичные лекции об условных рефлексах в новом отдаленном месте (собак пришлось перевозить) и при вообще необычной для наших животных обстановке. Все опыты с положительными условными рефлексами в числе 5–6, демонстрированные на первой лекции, протекли вполне хорошо. Все опыты на тех же животных с тормозными условными рефлексами (тоже в числе 5–6) на следующей лекции были неудачны, т. е. во всех них наступило растормаживание. Таким образом, одна и та же обстановка нисколько не нарушила положительных условных рефлексов, но, даже повторенная, резко повлияла на тормозные условные рефлексы.
К обсуждению этих фактов мы вернемся в более поздней лекции в связи с многочисленными фактами, относящимися к тому же пункту взаимоотношения между раздражительным и тормозным процессами.
Так как в первой фазе запаздывающего рефлекса мы имеем совершенно тот же тормозной процесс, что при угасании и условном торможении, то и в подробностях относительно него мы должны ожидать аналогичных отношений.
При угасании и условном торможении мы видели, что торможение, достигнутое для определенного условного раздражителя, простирается само собой и на другие условные раздражители, но в мере, которая определяется относительной физиологической силой этих раздражителей. Если другие условные раздражители слабее того, на котором было развито или выработано торможение, то вторичное последовательное торможение этих других раздражителей полное; если же они сильнее, то торможение только частичное, т. е. вырабатывающееся торможение как бы точно отмеривается по размеру того условного раздражителя, на котором оно вырабатывалось. То же самое наблюдается и на тормозном процессе запаздывающего рефлекса. С изменением силы условного раздражителя этого рефлекса установившееся отношение между недеятельной и деятельной фазами резко нарушается то в одну, то в другую сторону, соответственно изменению силы раздражителя.
Вот относящиеся сюда опыты на том же животном.
Опыт 22.X.1907
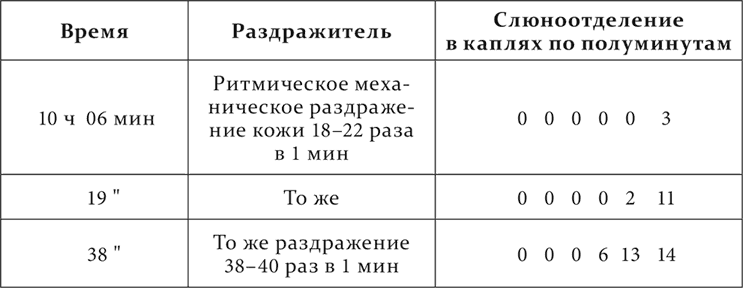
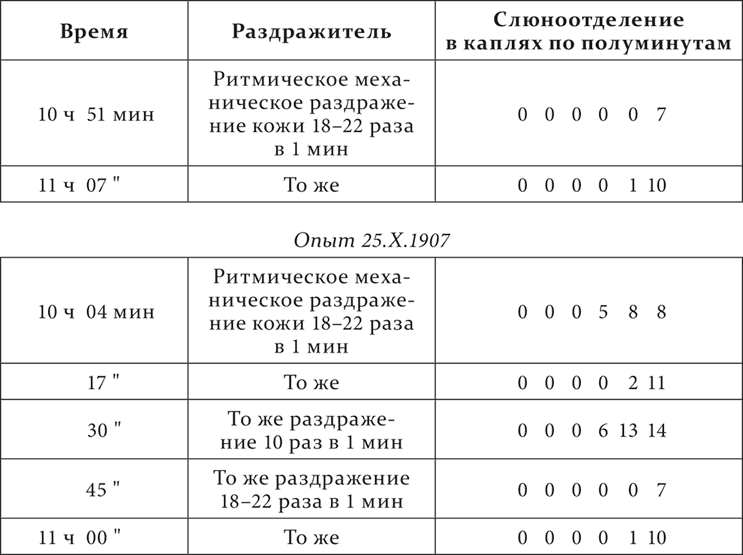
В опытах, если не отчетливо увеличение деятельной фазы с усилением условного раздражителя, то резко уменьшение ее с ослаблением раздражителя.
Точно таким же образом на запаздывании сказывается и сила безусловного раздражителя. Это удобно демонстрируется на условном пищевом раздражителе. Стоит только сравнить распределение фаз запаздывающего рефлекса на обычно питаемом животном и подвергнутом некоторому голоданию.
Опыт. Другое животное. Свисток средней силы – условный пищевой раздражитель. Отставление – 3 минуты.
Опыт 13.XII.1907 при обычном питании.
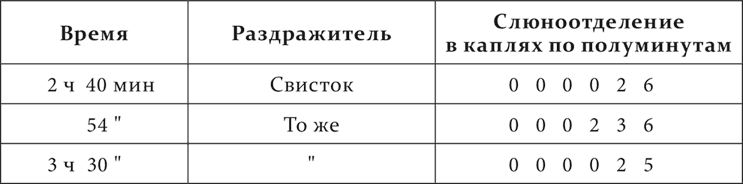
Опыт 15.XII.1907 на том же животном, но голодавшем 2 дня.
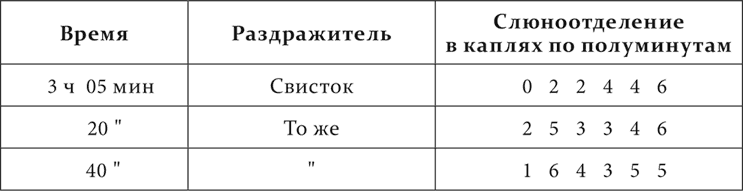
При голодном состоянии недеятельная фаза, тормозной процесс почти исчезли.
При запаздывании наблюдается такая же суммация торможения, какую мы видели при угасании и при условном торможении. Это явственно выступает на следующем опыте.
Механическое раздражение кожи – условный раздражитель, отставление – 3 минуты.
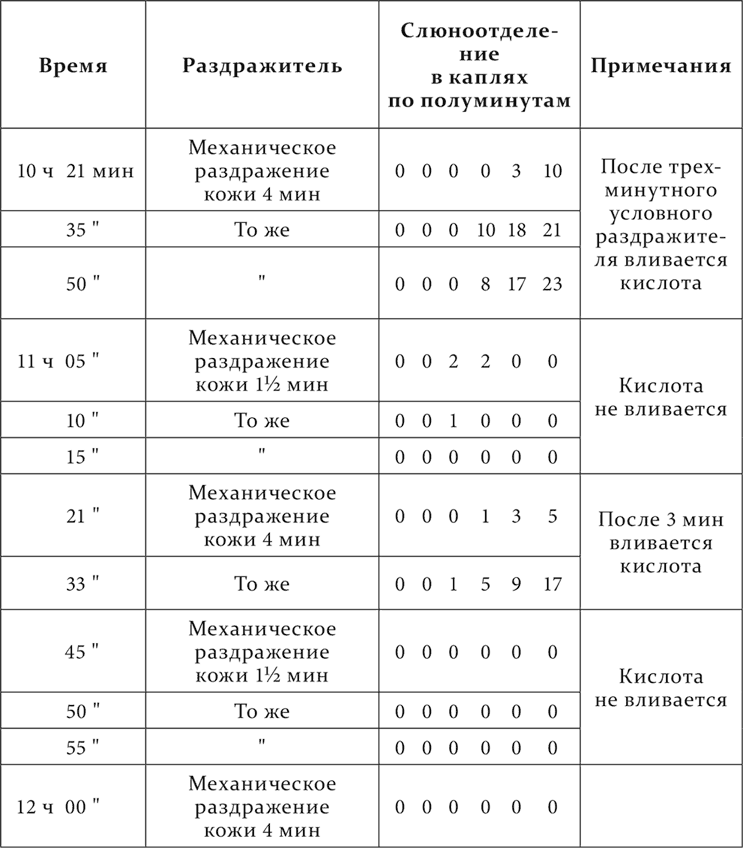
Механическое раздражение кожи в первые 1½ минуты, обусловливающее торможение, повторенное несколько раз без продолжения его во вторые 1½ минуты, повело к такому усилению тормозного процесса, что после этого примененное раздражение кожи во вторые 1½ минуты в первом случае сильно потеряло в своем положительном эффекте, а при повторении этой процедуры еще раз и совсем лишилось своего действия.
Как понимать, что в запаздывающем рефлексе один и тот же раздражитель сначала действует тормозящим образом, а затем возбуждающим? Что обусловливает противоположное действие одного и того же раздражителя, по-видимому, при совершенно одинаковых условиях? Можно ли привести это явление в согласие с уже нам известными фактами? Я думаю, что да. В третьей и пятой лекциях мы познакомились с агентами, из которых могут быть образованы как положительные, так и тормозные условные раздражители. Между ними упоминалось время как реальный агент. Там было разъяснено, как его надо понимать физиологически, и были приведены опыты, иллюстрирующие его роль как раздражителя. Особенно прошу вспомнить вариацию опыта, когда был применен суммарный раздражитель, состоящий из внешнего агента и из определенного момента времени. Тогда один внешний агент без надлежащего момента времени не действовал, а его действие появлялось и постоянно возрастало лишь по мере того, как приближался надлежащий момент времени. В запаздывающем рефлексе совершенно то же самое. В приведенных опытах безусловный раздражитель присоединялся к внешнему агенту только по истечении третьей минуты, т. е. только конец третьей минуты вместе с внешним агентом составляли тот настоящий суммарный агент, за которым непосредственно следовал безусловный раздражитель, и он-то (этот суммарный агент) специально и должен был сделаться условным раздражителем. Внешний же агент в соединении с другими моментами времени, отдаленными от конца третьей минуты, был другим суммарным агентом, который систематически не сопровождался безусловным рефлексом, и на него должен был вырабатываться тормозной рефлекс, как на всякий несколько отличающийся агент, что мы уже и видели при случае условного торможения и с чем мы в еще более резких примерах познакомимся в следующей лекции на случае так называемого нами дифференцировочного торможения. В запаздывающем рефлексе значение момента времени как раздражителя может быть представлено еще конкретнее и проще. Мы продолжаем избранный для образования условного рефлекса внешний агент известное время, а в каждый момент его продолжения он оказывается для организма другим. Мы знаем, как быстро мы привыкаем к запахам, звукам и освещению, т. е. ощущаем их все иначе. А это, конечно, значит, что нервные клетки, на которые падают раздражения, переживают различные последовательные состояния. И только определенное состояние, совпадающее с безусловным рефлексом, становится условным раздражителем. В следующей же лекции мы увидим, до какой степени простирается дифференцирование интенсивностей одного и того же агента, причем одна интенсивность становится положительно, а другая отрицательно действующим раздражителем. Таким образом, мы имеем несколько несомненных фактов, которые делают понятным факт запаздывающего рефлекса.
«Если индифферентные раздражения попадают в кору полушарий, когда там господствует тормозной процесс, то они хронически получают тормозную функцию, т. е., когда ими потом действуют на деятельные пункты коры, они производят в них тормозной процесс»
Факт развития запаздывания имеет очень большое значение в методическом отношении, и при разнообразном исследовании условных рефлексов с ним необходимо постоянно считаться. Конечно, для того, чтобы иметь данные о размере условного рефлекса и его многочисленных и тонких вариациях при разных условиях, мы должны отодвигать на больший или меньший период времени момент присоединения безусловного раздражителя от начального момента условного. Но это влечет за собой в большей или меньшей мере сейчас же развитие запаздывания, т. е. при этом рядом с раздражительным процессом возникает тормозной. Естественно, это осложняет исследование, потому что приходится иметь дело с двумя процессами, хотя бы вопрос касался одного раздражительного. Указанное обстоятельство, например, совершенно исключает возможность при обыкновенной форме наших опытов с условными рефлексами определения истинного латентного периода этих рефлексов. То, что мы обыкновенно при наших исследованиях называем латентным периодом, и то, что вы видели при нашем первом опыте (помните, там прошло 8 секунд между первым ударом метронома и появлением первой капли слюны), есть запаздывание, вмешательство тормозного периода и заслуживало бы точного термина: предварительный тормозной период. Поэтому для цели определения истинного латентного периода условных рефлексов необходимо пользоваться совпадающими рефлексами, когда безусловный раздражитель возможно скоро, через секунду или даже части секунды, следует за началом условного. И только при таких рефлексах, главным образом пользуясь двигательным компонентом наших рефлексов, можно убедиться, что величина латентного периода условных рефлексов действительно падает в категорию величин обыкновенного рефлекторного времени. Этими определениями у нас занимаются только теперь. Мы не придавали фундаментального значения определению истинного латентного периода условных рефлексов при установлении их рефлекторной натуры, считая главным доказательством этого их непременность и закономерность при известных условиях, так как и при обыкновенных рефлексах их латентные периоды значительно колеблются, находясь в зависимости от тех или других центральных связей, или путей. Можно с правом допустить, не угрожая рефлекторной натуре условных рефлексов, что в больших полушариях эти связи сложнее.
«Посторонние раздражители, повторяемые без дальнейших последствий для животного, также и сами по себе одни ведут к развитию в коре тормозного состояния»
Решение многих других вопросов относительно условных рефлексов, например вопрос о подлинном, беспрепятственном течении при них раздражительного процесса с самого его начала и т. п., также более или менее затрудняется вмешательством запаздывания. В этом отношении много помогает делу характер нервной системы разных животных. У иных из них, как это было указано выше, запаздывание развивается с трудом, и тогда раздражительный процесс или совсем не нарушается, или очень мало. Но и в руках экспериментатора есть некоторое средство ослаблять влияние запаздывания, ограничиваясь возможно коротким оставлением безусловного раздражителя от начала условного, если величина условного рефлекса вообще велика, т. е. если и за короткий срок она достаточна для сравнения при различных вариациях опыта. Но, с другой стороны, есть случаи, когда предварительный тормозной процесс представляет некоторую выгоду, дает возможность для решения некоторых вопросов, как с этим мы встретимся в следующей лекции. Таким образом, приходится приспособляться, соответственно пользоваться имеющимися отношениями.
Познакомившись в последних трех лекциях с основным фактическим материалом относительно внутреннего торможения условных рефлексов, нетрудно усмотреть его огромную биологическую ценность. Этим торможением постоянно корригируется и совершенствуется сигнализационная деятельность больших полушарий. Если в данное время сигнальный, условный раздражитель повторно не сопровождается его, так сказать, безусловно деловым раздражителем, он, как убыточный для организма, напрасно вызывающий трату энергии, на некоторое, как правило короткое, время лишается своего физиологического действия. Если условно действующий агент, постоянно встречаясь с посторонним агентом, при условии этой встречи не сопровождается безусловным раздражителем, он также теряет свое условное значение, но только в этой комбинации, являясь в таком случае как бы другим агентом. Наконец, если условный агент всегда только в определенный момент после своего возникновения совпадает с действием безусловного раздражителя, то он при более или менее отдаленно предшествующих этому раздражителю моментах делается недействительным, проявляясь все в большей и большей степени по мере приближения к моменту присоединения безусловного раздражителя. Таким образом, постоянно обнаруживается высшее приспособление организма к окружающим условиям, или, иначе говоря, более тонкое и точное уравновешивание организма с внешней средой.
Как ясно из всего сообщенного в последних трех лекциях, мы при угасании, условном торможении и запаздывании имеем перед собой образование условных тормозных рефлексов. Но эти же рефлексы могут быть получены еще и при совсем иной процедуре. Если с условными тормозными раздражителями одновременно на короткие сроки, но повторно, применять индифферентные агенты, то эти последние приобретают также тормозную функцию, т. е. и сами по себе вызывают в коре процесс торможения. Этот предмет специально разработан Ю. В. Фольбортом. Прежде всего он делал посторонние агенты индифферентными, повторяя их перед животным до тех пор, пока они не переставали тормозить положительные условные рефлексы и растормаживать тормозные. Затем он присоединял их повторно на короткие сроки то к угашенным условным рефлексам, то к тормозным комбинациям. И после этого испытывал их действие, комбинируя их с положительными условными раздражителями, причем и обнаруживал теперь их роль как условных тормозов. Так как эти испытания естественно приходилось ради точности результата повторять не один раз, то Фольборт один раз сопровождал пробную комбинацию безусловным рефлексом, в другой – нет, делая это строго поочередно, чтобы при этих повторных испытаниях комбинация не получила благодаря им постоянного, определенного, то положительного, то отрицательного характера. Но, конечно, бесспорнее и убедительнее была та форма опыта, когда он при испытаниях всегда сопровождал пробную комбинацию безусловным рефлексом, а торможение, несмотря на это, при повторении опыта все же резко выступало.
Вот эти опыты. На собаке применялся натуральный условный пищевой рефлекс. Когда он угашался до нуля, к нему повторно присоединялись сделанные раньше индифферентными удары метронома, а затем при продолжении этого приема изредка пробовалась комбинация ударов метронома с положительным пищевым рефлексом, причем она всякий раз подкреплялась безусловным рефлексом, едой.
Опыт 5.IX.1911 после 10 совпадений ударов метронома с угашенным условным рефлексом.

Опыт 1.XII.1911 после 19 совпадений ударов метронома с угашенным условным рефлексом.
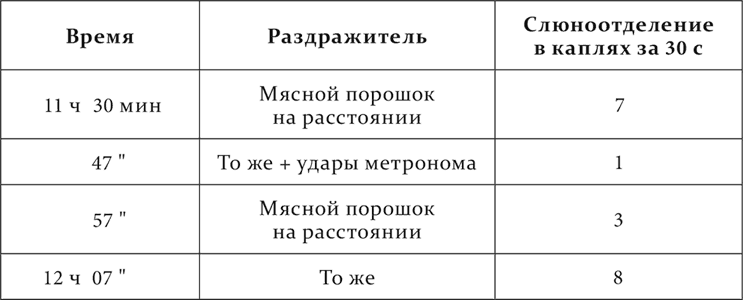
Опыт 18.XII.1911 после 26 совпадений ударов метронома с угашенным условным рефлексом.
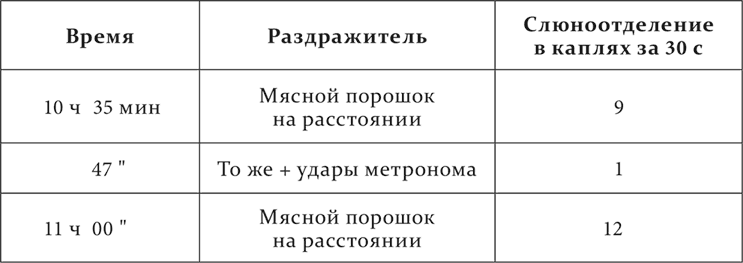
Совершенно очевидно, что ранее индифферентные удары метронома после совместных применений с угашенным условным раздражителем стали тормозящим агентом. Эти новые, иначе полученные условные тормозные раздражители оказались по своим свойствам вполне сходными с теми, которые мы изучили при угасании, условном торможении и запаздывании.
Новый условный тормоз, полученный с одним угашенным рефлексом, действует тормозящим образом и на другие условные рефлексы.
Опыт 15.III.1911. Другая собака. Удары метронома совпадали с угашенным условным натуральным пищевым рефлексом, а затем применялись вместе с запахом камфоры, искусственным условным пищевым раздражителем.
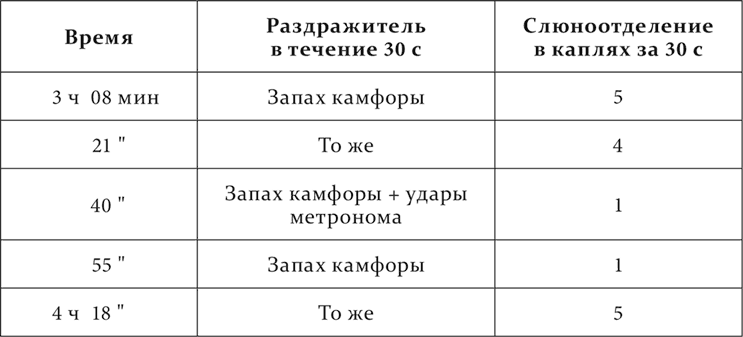
Новый условный тормоз обнаруживает свое действие не только при одновременном применении с положительными условными раздражителями, но и последовательно.
При его повторениях он также суммируется, и, наконец, его действие может растормаживаться.
Итак, оказывается: если индифферентные раздражения попадают в кору полушарий, когда там господствует тормозной процесс, то они хронически получают тормозную функцию, т. е., когда ими потом действуют на деятельные пункты коры, они производят в них тормозной процесс.
Однако не следует упускать из виду и то, что, как увидим это потом, посторонние раздражители, повторяемые без дальнейших последствий для животного, также и сами по себе одни ведут к развитию в коре тормозного состояния. Следовательно, при только что сообщенных опытах должно быть специально проконтролировано, насколько главное участие в образовании новых условных тормозов принимают именно условные тормозные рефлексы, с которыми повторно сочетаются посторонние раздражители.
Назад: Лекция пятая
Дальше: Лекция седьмая

