Книга: Лекции о работе больших полушарий головного мозга
Назад: Лекция шестая
Дальше: Лекция восьмая
Лекция седьмая
Анализаторная и синтезирующая работа больших полушарий: а) первоначальный обобщенный характер условных раздражителей; б) дифференцировочное торможение
Мм. гг.! Условно-рефлекторные агенты, беспрерывно сигнализируя непосредственно благоприятствующие и разрушающие влияния окружающей среды на организм, представляют собой, в соответствии с бесконечным разнообразием и колебаниями этой среды, как мельчайшие ее элементы, так и то большие, то меньшие комплексы из них. А это делается возможным только благодаря тому, что нервная система обладает как механизмами, выделяющими для организма из сложности среды отдельные ее элементы, – анализаторными механизмами, так и механизмами, соединяющими, сливающими эти элементы для организма в те или другие комплексы, – механизмами синтезирующими. Следовательно, при изучении высшей нервной деятельности, деятельности коры больших полушарий, мы должны встретиться с явлениями как нервного анализа, так и нервного синтеза, к чему мы теперь и приступаем.
Нервная система есть всегда больший или меньший комплекс анализаторных приборов, анализаторов. Оптический отдел выделяет для организма световые колебания, акустический – воздушные и т. д. В свою очередь каждый из этих отделов дробит соответствующую часть внешнего мира на длиннейший ряд отдельных элементов. К каждому данному анализатору должны быть отнесены как периферические приборы всевозможных афферентных нервов (трансформаторы, из которых каждый превращает в нервный процесс только определенную энергию), так и сами нервы и клеточные мозговые концы. Понятно отсюда, в анализаторной работе участвуют как те, так и другие. Более низкие степени анализа свойственны, конечно, и низшим отделам нервной системы (как и малодифференцированному веществу животных без нервной системы), так как и организм, лишенный головного мозга, отвечает очень различно на различные по месту, интенсивности и качеству раздражения его внешней поверхности. Но высший тончайший анализ, на который способно данное животное, достигается только при помощи больших полушарий. Ясно, что усложнение связи животного организма с окружающим миром, все более разнообразное и более точное приспособление к внешним обстоятельствам, более совершенное уравновешивание организмов внешней среды идет параллельно и неразрывно с этой все прогрессирующей анализаторной деятельностью нервной системы. В современной физиологии изучение анализаторной деятельности составляет очень важный отдел так называемой физиологии органов чувств. Этот отдел представляет огромный интерес и является в высшей степени содержательным, отчасти, может быть, потому, что имел привилегию привлекать на себя внимание величайших физиологов, с гениальным Гельмгольцем во главе. В этом отделе заложены прочные основы учения о деятельности как периферических конструкций анализаторов, так и их мозговых концов, указаны многие пределы анализаторной работы (у человека), разъяснены некоторые сложные случаи этой работы и установлены многие частные правила, относящиеся к ней. Но весь этот огромный материал, субъективный в преобладающей его части, построен на наших ощущениях, этих наипростейших субъективных сигналах объективных отношений организма к внешнему миру. И это составляет значительный недостаток этого отдела. Из него таким образом исключено все то, что должно дать физиологии изучение разнообразия и развития анализаторной деятельности в животном царстве; вследствие этого же натуралист лишен неисчерпаемой возможности экспериментирования, которая представляется ему опытами над животными. Условные рефлексы все изучение важнейшей функции нервной системы, функции анализа, отдают в руки чистой физиологии, строго объективного, естественно-научного исследования. С помощью них удобно и точно определяются объем и предел анализаторной деятельности разных животных и выясняются законы этой деятельности. Пусть сейчас физиология анализаторов у животных еще очень скудна, но ее содержание все растет и будет расти все стремительнее, расширяя и углубляя наше знание о связи животного организма с окружающим миром.
«Нервная система есть всегда больший или меньший комплекс анализаторных приборов, анализаторов»
Как можно объективно, по видимым реакциям, изучать у животных работу анализаторов? Как уже мной кратко указывалось раньше, всякое колебание окружающей животное среды влечет за собой если не специальные – врожденные или приобретенные реакции, – то вообще ориентировочную реакцию, исследовательский рефлекс. Он, этот рефлекс, прежде всего и может служить для констатирования того, в какой степени нервная система данного животного может отличить одно от другого. Если, положим, в обычной среде между прочим имеется определенный звук – тон, то хотя бы и небольшое изменение его высоты должно вызвать и вызывает на себя ориентировочную реакцию, т. е. определенную постановку животным в отношении звука ушного аппарата. То же самое относится и ко всяческим другим раздражениям, простым или сложным, при малейших их изменениях. Но это, конечно, только в том случае, если анализаторный аппарат, по тонкости его конструкции, в состоянии отметить происшедшую разницу. В таком случае исследовательский рефлекс может служить для исследования или сам по себе, или, вследствие влияния его на условные рефлексы как чувствительнейшие реакции организма, в виде то тормозящего, то растормаживающего агента. Однако, как постоянный прием для изучения анализаторной деятельности нервной системы, исследовательский рефлекс, несмотря на его часто большую чувствительность, представляет и много неудобств. Из них главнейшее то, что он при некоторых слабых раздражениях очень летуч, не может быть повторяемым, чем существенно и подрывается возможность точного исследования с его помощью. Наоборот, условный рефлекс сам по себе является объектом, в высшей степени соответствующим цели данного изучения. Из строго определенного внешнего агента мы делаем обыкновенной процедурой специальный условный раздражитель и повторениями постоянно утверждаем его в этой роли, а ближайший к нему по интенсивности, месту или качеству агент, при известных условиях, может оставаться постоянно без этой специальной реакции и таким образом легко и точно отличается от него.
Так как в действительности анализаторная и синтезирующая работы нервной системы постоянно встречаются и перемежаются между собой и так как анализу внешних агентов с помощью условных рефлексов предшествует особенное нервное явление – род синтентической деятельности, то им прежде всего и приходится заняться.
«Пусть сейчас физиология анализаторов у животных еще очень скудна, но ее содержание все растет и будет расти все стремительнее, расширяя и углубляя наше знание о связи животного организма с окружающим миром»
Всякий агент, из которого мы образуем наш условный раздражитель, непременно носит обобщенный характер. Если нами сделан условный раздражитель, например из тона в 1000 колебаний в секунду, то вместе с тем и многие другие тоны сами по себе получают то же условное действие, но тем меньшее, чем они дальше отстоят по высоте вверх и вниз от тона в 1000 колебаний. Точно так же, если механическое раздражение определенного места кожи сделано условным раздражителем, то такое же действие принадлежит затем и другим местам кожи тем в меньшем размере, чем дальше эти места находятся от места, на котором вырабатывался условный рефлекс. Тот же факт повторяется при раздражениях других рецепторных поверхностей.
Описанное явление с биологической точки зрения можно было бы понять так, что натуральные агенты большей частью не оказываются строго определенными, а скорее постоянно колеблются, меняются, переходя из одной родственной группы в другую. Возьмем для примера звук, издаваемый животным-врагом и являющийся условным раздражителем оборонительного рефлекса животного-жертвы. Он может представлять большие колебания в высоте, силе и составе в зависимости от напряжения голосового аппарата, расстояния, резонанса.
Дальше мы встретились при условных рефлексах и с таким обобщением, жизненное значение которого не было ясно прямо. До сих пор мы имели дело с обобщением условного раздражителя в пределах одного анализатора. В случае условных следовых раздражителей, именно образованных на поздних следах (1–3 минуты спустя после прекращения действия внешнего агента), приходится видеть совсем другое. Здесь имеется обобщение универсальное. Мы образовали условный раздражитель, например из механического раздражения кожи, а у нас в качестве условных раздражителей начинают действовать агенты, относящиеся к другим анализаторам и не стоящие ни в какой связи с нашим рефлексом. Я позволю себе остановить ваше внимание на этом факте несколько дольше, так как он представляет интерес в отношении его разработки.
Вот примеры этих опытов из работы Гроссмана. Механическое раздражение кожи – условный следовой (через минуту по прекращении наличного раздражения) кислотный раздражитель. Холод и тон применяются впервые.
Опыт 6.II.1909
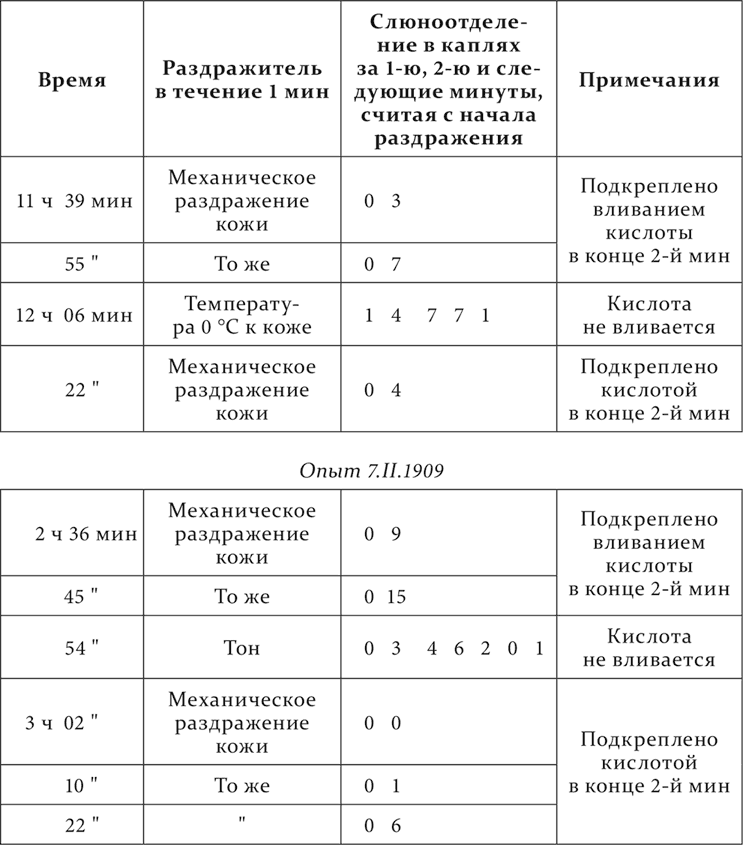
Мы видим, что и агенты, никогда ранее не приходившие в связь с действием кислоты, оказались возбудителями кислотной реакции. Мы замечаем вместе с тем, что впервые испробованные агенты действуют, так сказать, по той же программе, как и выработанный следовой раздражитель, т. е. действуют не во время их применения, а лишь главным или исключительным образом по прекращении их. Это, конечно, с самого начала склоняет к заключению, что здесь дело идет об обобщенных рефлексах, воспроизводящихся по типу выработанного следового рефлекса. Но этого, разумеется, мало, и, ввиду особенности факта, мы подвергли его многократному переисследованию. Принимая во внимание все нам известное относительно условных рефлексов, мы сделали еще два возможных предположения о ближайшем механизме разбираемого факта.
Так как следовой рефлекс на поздних следах образуется нелегко и нескоро, то нельзя при этом не заметить, что до образования его очень быстро образуются условные рефлексы на посторонние раздражения, случайно совпадающие с применением безусловного раздражителя и идущие от самого экспериментатора. Здесь эта опасность особенно велика. Поэтому эти опыты были повторены с удалением экспериментатора из комнаты, где находилась собака, и в новом специальном здании, чем было совершенно исключено всякое несознаваемое влияние со стороны экспериментатора на животное. Но чрезвычайная обобщенность следового рефлекса оставалась и тут.
Второе предположение, казавшееся тоже очень серьезным, состояло в следующем. При образовании у собак в лаборатории в первый раз условных рефлексов соответствующий рефлекс прежде всего образуется на всю лабораторную обстановку, начиная с привода животного в экспериментальную комнату. Его можно было бы назвать условным обстановочным рефлексом. Затем, когда начинает появляться рефлекс на применяемый нами специальный агент, элементы обстановки постепенно теряют свое условное действие, очевидно, в силу развития на них постепенно внутреннего торможения. Но это торможение, особенно на первых порах, легко и растормаживается всякими экстренными посторонними агентами. Вот относящийся сюда яркий случай, очень обычный в лаборатории при старой манере, когда экспериментатор и собака находились вместе в одной комнате. Обстановочный рефлекс, когда слюна текла сама по себе, миновал, и теперь на фоне покоя железы имеется свой специальный условный рефлекс. Но стоило мне впервые зайти в комнату для ознакомления с положением дела, и у собаки наступало сплошное слюноотделение, пока я находился в комнате. Я, как экстренный посторонний агент, растормаживал уже заторможенный обстановочный рефлекс. Мы предположили: не есть ли то, что мы называем обобщенным следовым рефлексом, растормаживание обстановочного рефлекса? Но при проверке и это предположение должно было отпасть. Во-первых, чрезвычайное обобщение следовых раздражителей получается легко и постоянно у собак, у которых обстановочный рефлекс давно и прочно заторможен, так что почти невозможно его растормозить. Во-вторых, нами проконтролировано особенно строго следующее обстоятельство. Прошу припомнить, что при запаздывании растормаживание недеятельной фазы рефлекса происходило сейчас же, как пускался в ход посторонний раздражитель. Здесь же при следовых рефлексах этот раздражитель производит эффект почти всегда только по его прекращении. Если, настаивая на рассматриваемом предположении, принимать, что в данном случае все посторонние раздражители действуют почему-то всегда как очень сильные и потому не растормаживают, а еще глубже при их наличии тормозят обстановочный рефлекс (как это было и на запаздывающих рефлексах) и растормаживают только своими следами как слабеющим раздражением, то против такого изысканного допущения говорит следующее. При повторном применении сильных посторонних раздражителей их тормозящее действие непременно постепенно падает, и затем наступает фаза, когда они прямо только растормаживают, что мы и видели при запаздывающих рефлексах. Здесь при следовых рефлексах этого никогда не наблюдается, и многократно повторяющиеся посторонние раздражители производят слюноотделение только после. Наконец, в-третьих, при следовых рефлексах после применения посторонних раздражителей, которые, конечно, не сопровождаются безусловным раздражителем, специальный условный раздражитель временно слабеет, даже доходит до нуля, что было бы совершенно непонятно, если бы здесь дело шло о растормаживании обстановочного рефлекса. При обобщенном следовом рефлексе это ослабление – простой случай угасания после неподкрепления.
«Так как следовой рефлекс на поздних следах образуется нелегко и нескоро, то нельзя при этом не заметить, что до образования его очень быстро образуются условные рефлексы на посторонние раздражения, случайно совпадающие с применением безусловного раздражителя и идущие от самого экспериментатора»
Итак, при образовании условных рефлексов сперва определенный безусловный рефлекс связывается с массой внешних агентов, один раз благодаря внешней среде, действующей на организм совокупностью ее элементов (условный синтетический обстановочный рефлекс), в другой раз в силу свойств самой нервной системы, когда внешнему элементарному агенту ею придается в качестве условного раздражителя обобщенный в большей или меньшей мере характер. Этот факт, как это частью ясно прямо, частью указано выше, конечно, оправдывается действительными отношениями животного к окружающему миру. Но вместе с тем очевидно, что тот же факт должен иметь лишь ограниченное и временное значение, когда при известных условиях возможная, приблизительная и общая связь заменяется действительной, точной и специальной связью.
Как же происходит специализация условного раздражителя, дифференцирование внешних агентов? Сначала нам казалось, что здесь имеют место два приема. Один – это только многократное повторение определенного агента в качестве условного раздражителя с постоянным подкреплением безусловным рефлексом. Другой – перемежающееся противопоставление этого определенного, постоянно подкрепляемого, условного раздражителя с близким к нему агентом, но не сопровождаемым безусловным раздражителем. В настоящее время мы склонны признавать действительность только последнего приема. С одной стороны, мы имели условные раздражители, повторенные тысячекратно и которые, однако, через это одно не делались узко специализированными. С другой – было замечено, что даже однократная проба каждого из родственных агентов без подкрепления и редкое (через дни и даже недели) такое же (т. е. без подкрепления) применение ряда их (каждый раз все нового агента), при повторениях с подкреплением основного условного раздражителя, ведет, однако, к специализированию его. Поэтому мы постоянно пользовались вторым приемом, во всяком случае несравнимо скорее приводящим к цели дифференцировки агентов.
«При повторном применении сильных посторонних раздражителей их тормозящее действие непременно постепенно падает, и затем наступает фаза, когда они прямо только растормаживают»
Мы прежде всего остановимся на подробностях хода процесса дифференцировки внешних агентов при помощи условных рефлексов. Здесь на первом месте должен быть поставлен факт, казавшийся нам в свое время довольно долго загадочным. Когда по образовании условного раздражителя из определенного внешнего агента впервые пробуют другие агенты, очень близкие к нему, например тоны, соседние с тоном-раздражителем, то часто получают от них условно-рефлекторный эффект, резко меньший, чем от выработанного условного раздражителя. Но затем при повторении их, конечно без подкрепления, этот эффект быстро увеличивается, сравнивается с эффектом условного раздражителя и потом только постепенно начинает уменьшатся, доходя до нуля. Значит, сперва разница между агентами обнаруживается сразу, затем она почему-то исчезает и потом только снова медленно проявляется и, наконец, становится абсолютной. Как понимать этот факт? Мы воспользовались для этого сходством теперешнего факта с подобным фактом, ранее нами анализированным. Вы помните, что при выработке условного торможения наблюдалось такое же отношение. Когда впервые к условному раздражителю присоединяется новый агент, из которого потом образуется условный тормоз, то эта комбинация производит или эффект, свойственный условному раздражителю, но небольшой, или даже остается совсем бездейственной. Но затем, несмотря на то, что мы не сопровождаем ее безусловным рефлексом, она снова и быстро обнаруживает полное действие и лишь позже при повторении постепенно доходит до нуля. При условном торможении мы объяснили – и с полным правом – это явление так, что прибавочный агент вызвал ориентировочную реакцию и сейчас же затормозил действие условного раздражителя, с которым он был комбинирован. При повторении ориентировочная реакция быстро ослабела, и действие условного раздражителя временно восстановилось, чтобы при дальнейших повторениях постепенно быть подавленным развивающимся торможением. Мы и здесь можем представить себе, что сходственные агенты, близкие к агенту условного рефлекса, как бы состоят из двух частей (как и тормозная комбинация) – одной общей с условным раздражителем и другой особенной. Первая есть причина того, что близкие агенты действуют подобно выработанному условному раздражителю. Вторая сначала дает основание для временного ориентировочного рефлекса, тормозящего условный эффект первой части сейчас же и тоже на время, а впоследствии – для развития постоянной и окончательной дифференцировки близких агентов.
Веским доказательством основательности нашего понимания явлений при начале выработки дифференцировок является то, что сходство в ходе этих явлений при развитии условного торможения и выработке дифференцировок простирается и на имеющиеся здесь подробности. Как в том, так и в другом случае встречаются одни и те же вариации. Первоначальное ослабление эффекта близких агентов иногда переходит на короткое время в усиление сравнительно с тем, что дает специальный условный раздражитель, чтобы затем, вернувшись к величине последнего, постепенно уменьшаясь, кончить нулем. Всего чаще за первоначальным ослаблением следует возврат почти к величине условного раздражителя, а потом развитие дифференцировки. Редко без этих колебаний прямо наступает постепенное образование дифференцировки и, наконец, тоже редок непосредственный переход первоначального ослабления в прогрессирующее дифференцирование.
Так как при описании выработки условного торможения я только сообщил об этих вариациях, не приводя протоколов опытов, то здесь могу с избытком пополнить этот дефект. В данном случае (работа М. М. Губергрица) эти вариации были прослежены в особенности систематично.
Ряд опытов на одном и том же животном. Вертящийся в направлении часовой стрелки предмет – условный пищевой раздражитель, тот же предмет, вертящийся в обратном направлении, – дифференцируемый агент.
Опыт 15.II.1917

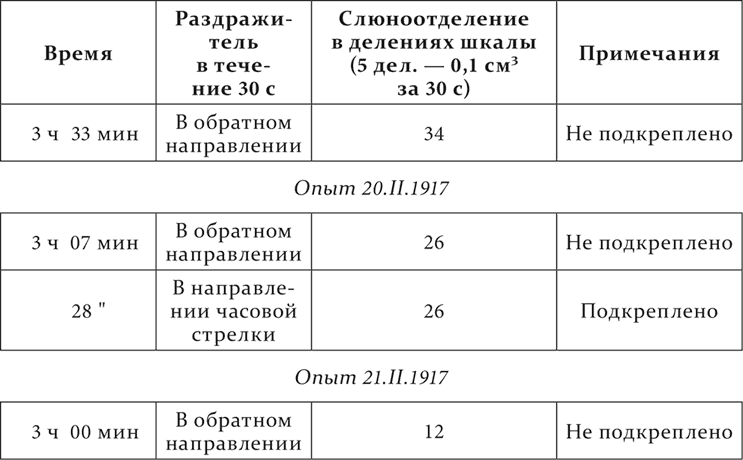
Затем с колебаниями дифференцируемый агент доходит до нуля действия.
Другая собака. Тон – условный пищевой раздражитель, полутон к нему – дифференцируемый агент.
Опыт 12.X.1917
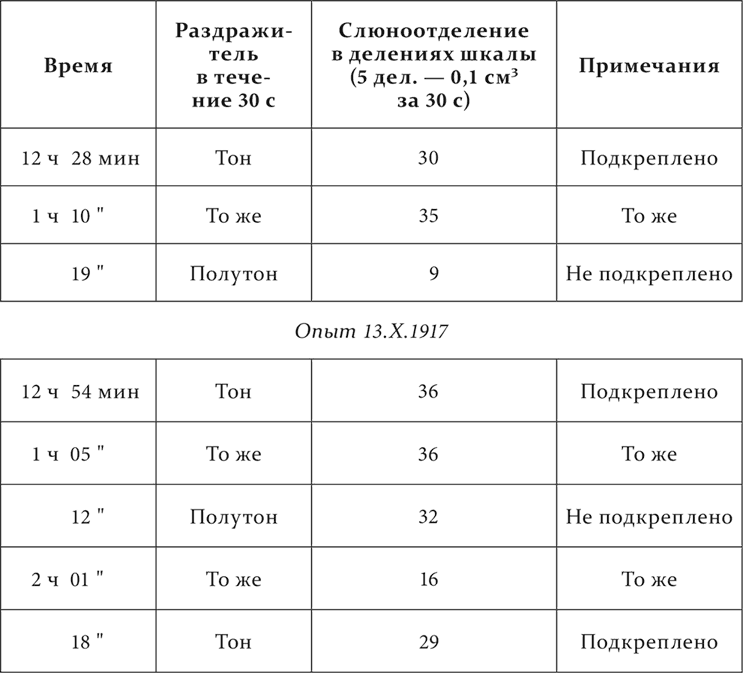
Далее с колебаниями к тринадцатому повторению полутон доходит до нуля.
Та же собака, что в первых опытах. Появление круга перед глазами собаки – условный пищевой раздражитель, появление квадрата той же площади и того же освещения – дифференцируемый агент.
Опыт 28.XI.1916
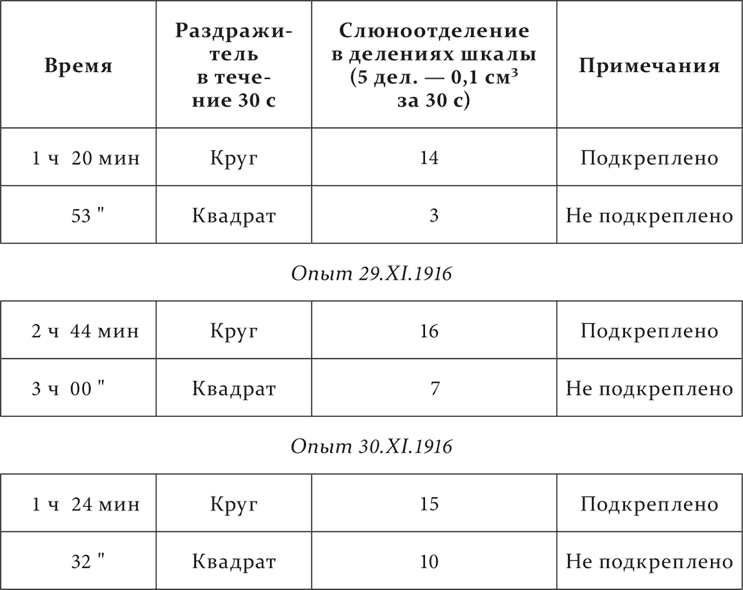
Затем с колебаниями к одиннадцатому повторению квадрат становится совершенно недействительным.
При дифференцировании внешних агентов с помощью условных рефлексов, помимо только что описанной начальной стадии, связанной с вмешательством ориентировочной реакции, привлекли к себе наше внимание и многие другие стороны этого нервного акта.
Можно, конечно, производить сразу дифференцировку агента условного рефлекса от очень близкого к нему агента, но можно подходить к ней и постепенно, начиная сперва с дифференцирования отдаленных агентов. Оказалась огромная разница между одним и другим приемом. Мы начинаем прямо с дифференцирования близкого агента и, несмотря на значительное число противопоставлений одного агента другому, искомой разницы не оказывается. Переходим тогда к более отдаленным и потом постепенно возвращаемся к первому – теперь через этапы достигнутых дифференцировок на более отдаленных агентах, в целом в гораздо более короткий срок, имеем диференцировку близкого агента.
Вот подтверждающие это отношение опыты, тоже из работы Губергрица.
У одной собаки условным пищевым раздражителем был сделан круг из белой бумаги, появляющийся перед ее глазами. От него начали дифференцировать круг из бумаги № 10 Циммермановской коллекции (50 оттенков от белого цвета до черного); 75 применений серого круга (№ 10), конечно, без подкрепления, чередуемых с белым, постоянно подкрепляемым, не обнаружили ни малейших признаков различения. Тогда перешли к дифференцированию сперва более темного круга № 35, а затем последовательно к кругу № 25 и кругу № 15 и, наконец, вернулись к кругу № 10. Теперь полная дифференцировка была достигнута всего через 20 применений этих кругов, считая все вместе.
«Наши неоднократно повторенные опыты показали, что дифференцирование различных агентов достигает приблизительно одинаковых степеней, применяются ли они в качестве положительных условных раздражителей или отрицательных, тормозных»
То же было проделано на другой собаке также с глазным раздражителем, но в другой форме. У нее условным пищевым раздражителем был сделан также круг, но от него дифференцировался эллипс, из той же бумаги и той же величины площади с отношением полуосей 8:9. Несмотря на 70 противопоставлений, никакой дифференцировки не обнаружилось. Когда же после этого стали последовательно дифференцировать эллипсы, начиная с эллипса с отношением осей 4:5 и переходя через эллипсы 5:6, 6:7, 7:8, то уже после 18 применений, считая вместе все, потребовавшиеся для выработки каждой из них, имелась дифференцировка и на эллипс 8:9.
Если мы производим дифференцировку постепенно, начиная с отдаленных агентов, то скоро бросается в глаза следующий постоянный факт. При выработке дифференцировки, хотя бы и грубой, в первый раз дело идет довольно медленно, особенно если настаивать на абсолютном дифференцировании, т. е. на нуле эффекта от дифференцируемого агента. Но раз на первом этапе достигнута абсолютная или близкая к ней дифференцировка, то процедура дальнейшего дифференцирования происходит все быстрее, но опять несколько замедляясь, когда мы подходим к предельной дифференцировке.
Привожу один из примеров.
У собаки белый круг определенного диаметра – условный пищевой раздражитель, тоже белые эллипсы той же величины площади с разными отношениями их полуосей – дифференцируемые агенты. Первый эллипс с отношением полуосей 4:5 потребовал 24 применений (34 деления за 30 секунд действия дал круг, а эллипс только 4). При следующем эллипсе 5:6 нужно было только троекратное его повторение, чтобы его эффект оказался даже нулевым. То же и со следующим эллипсом 6:7.
Ход дифференцирования, выключая начальную фазу с участием ориентировочной реакции, может быть как почти прямолинейным, так и волнообразным, и последнее не всегда может быть отнесено к каким-нибудь внешним нарушающим влияниям, а, вероятно, связано с колебаниями внутреннего нервного процесса, при этом происходящего. Прочность дифференцировки определяется по длине промежутка между применением условного раздражителя и отдифференцированного от него агента, при котором дифференцировка сохраняется. Сначала эти промежутки короткие, затем они становятся все длиннее. Мы считали дифференцировку окончательно выработанной, когда она через сутки, поставленная в опыте на первом месте, т. е. ранее других раздражений, оказывалась полной.
Наши неоднократно повторенные опыты показали, что дифференцирование различных агентов достигает приблизительно одинаковых степеней, применяются ли они в качестве положительных условных раздражителей или отрицательных, тормозных.
Так же могут и в той же степени дифференцироваться агенты и в роли следовых условных раздражителей, причем тоже как положительных, так и тормозных.
Вот пример дифференцировки следового тормоза из работы Фролова.
У собаки удары метронома (104 удара в минуту) – условный пищевой раздражитель. Определенный тон органной трубы (№ 16), продолжавшийся 15 секунд и прекращаемый за минуту до ударов метронома, – условный тормоз. Такая же комбинация ударов метронома с соседним тоном той же органной трубы (№ 15) сопровождалась безусловным рефлексом и стала положительным раздражителем. Дифференцировка следового тормоза представилась в следующем виде.
Опыт 25.IV.1922
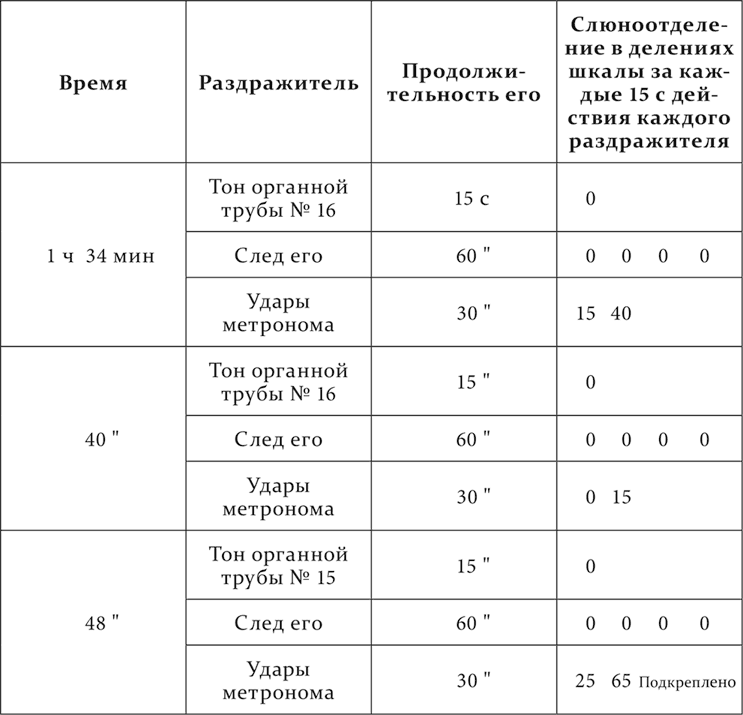
Надо прибавить, что эта дифференцировка была достигнута только благодаря крайней постепенности при ее выработке. Приходилось начинать с секундных следов и с удаленных тонов.
«Между констатированием нервной системой разницы между внешними агентами вообще и дифференцированием тех же агентов при помощи условных рефлексов есть существенная разница»
Теперь пора определенно поставить вопрос: каким нервным процессом обусловливается переход обобщенного характера условного раздражителя к его крайней специализации, или, иначе говоря, дифференцирование близких агентов? После знакомства с внутренним торможением и после того, что уже сообщено выше в этой лекции, нетрудно догадаться, в чем тут дело, – и наши дальнейшие многочисленные опыты не оставляют сомнения в том, что в основании дифференцирования лежит тормозной процесс, т. е. как бы постепенное заглушение сначала широко возбужденного мозгового конца анализатора, исключая его мельчайшую часть, отвечающую данному условному раздражителю. Вот первое фактическое оправдание этого вывода. Указанной выше процедурой мы один агент – наш условный раздражитель – отдифференцировали от близкого к нему агента: он дает постоянный и полный условный эффект, отдифференцированный же является, по-видимому, совершенно недействительным. Теперь после условного раздражителя применим этот последний, а за ним сейчас же или через некоторый промежуток повторим опять наш условный раздражитель – он будет иметь временно маленькое или никакого действия.
Вот пример из работы В. В. Белякова.
У собаки определенный тон органной трубы – условный пищевой раздражитель, от него прочно отдифференцирован тон на 1/8 часть выше.
Опыт 14.II.1911
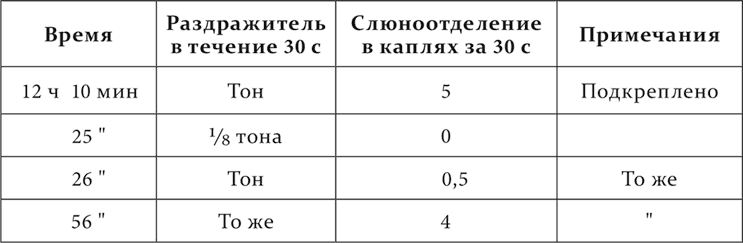
Следовательно, после применения отдифференцированного тона в нервной системе остается на некоторое время тормозной процесс, который дает себя знать на испытанном последовательно условном раздражителе, уменьшая или совершенно уничтожая его эффект, иначе говоря, перед нами хорошо нам знакомое по ранним лекциям последовательное торможение.
Таким образом, мы имеем четвертый случай внутреннего торможения, которое мы соответственно назвали дифференцированным торможением. Может быть, было бы допустимо его отождествить и объединить общим прилагательным (именно, дифференцировочное) с условным торможением, так как в обоих случаях идет дело об устранении с помощью торможения действия посторонних (простых или сложных) агентов, получивших это их действие без специальной выработки, только вследствие той или другой связи их с выработанным условным раздражителем.
Последовательное торможение, с которого мы начали при процессе дифференцирования, представляется одинаковым с последействием при условном торможении в том отношении, что оно также с повторениями делается все более во времени ограниченным, простираясь на десятки минут вначале и, наконец, сокращаясь до секунд.
Здесь заслуживает еще особенного упоминания тот факт, что чем выше степень дифференцировки, т. е. чем более близки друг к другу дифференцируемые агенты, тем значительнее последовательное торможение при прочих равных условиях.
Вот иллюстрация этого из работы Белякова.
Определенный тон – условный пищевой раздражитель, ½ и 1/8 того же тона – дифференцируемые агенты.
Опыт 19.III.1911
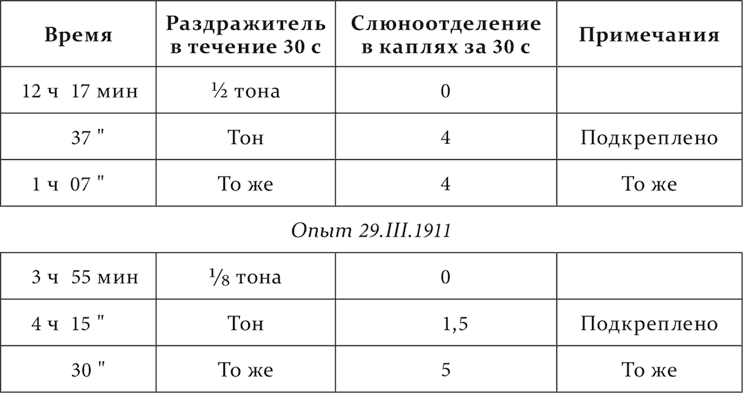
Но и помимо последействия дифференцировочное торможение также вполне сходствует с ранними случаями торможения.
Оно также суммируется при повторениях.
Вот опыты также из работы Белякова, поставленные на эту тему.
Другая собака. Тон – условный пищевой раздражитель, полутон ниже – дифференцируемый агент.
Опыт 8.VI.1911
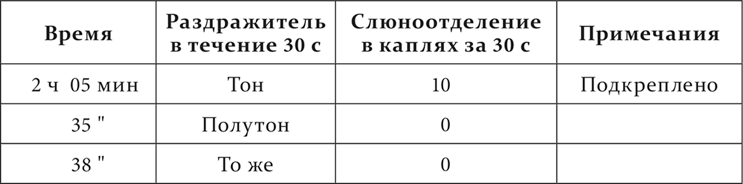
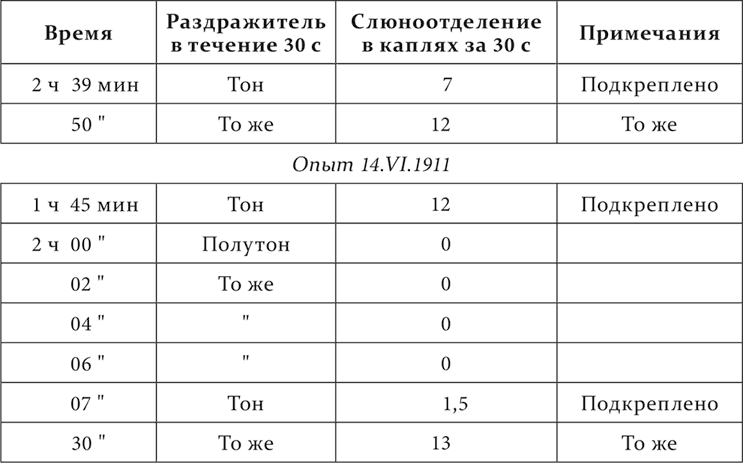
Дифференцированное торможение так же, так сказать, отмеривается по размеру того раздражительного процесса, с которым имеет дело при выработке, и потому нарушается при увеличении интенсивности дифференцируемого агента и при изменении общей и местной возбудимости центральной нервной системы.
Если мы пользуемся пищевым рефлексом, то стоит только повысить пищевую возбудимость, делая опыт в более поздний час дня, т. е. ближе к сроку обычного корма собаки, или же заставить голодать животное лишний день, как ранняя вполне выработанная дифференцировка оказывается уже недостаточной.
Точно так же, если общую возбудимость центральной нервной системы повысить впрыскиванием кофеина, совершенная ранее дифференцировка нарушается. Вот опыт, сюда относящийся, из работы П. М. Никифоровского. Механическое раздражение кожи на передней лапе – пищевой условный раздражитель, такое же раздражение кожи на спине – отдифференцированный агент.
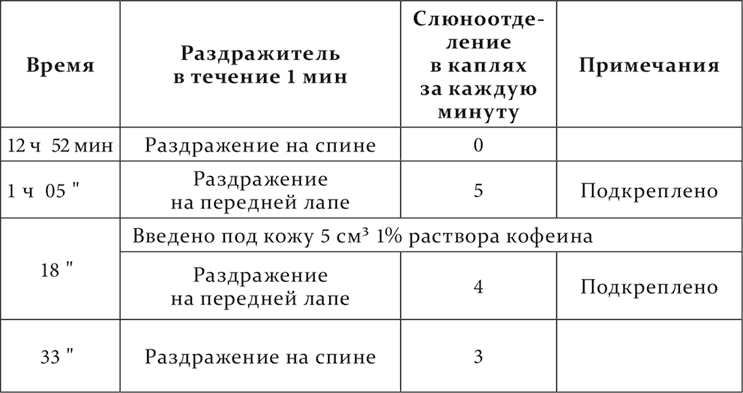
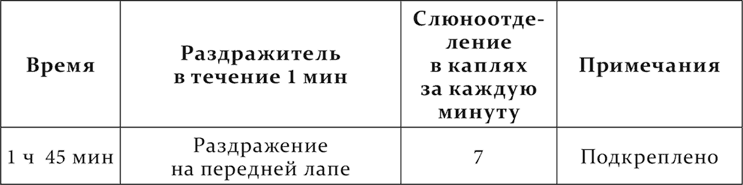
Наконец, дифференцировочное торможение, как и все остальные случаи внутреннего торможения, подвергается растормаживанию, т. е. и это торможение на короткое время устраняется, уступая место положительному эффекту под влиянием посторонних раздражений. Вот два опыта из работы Белякова на одном и том же животном с разными посторонними раздражителями.
Тон (800 колебаний в секунду) – условный пищевой раздражитель, 1/8 тона (812 колебаний в секунду) – основательно отдифференцированный агент. Оба посторонние раздражителя, бульканье и запах уксусного амила, испытанные отдельно, не сопровождались слюноотделением.
Опыт 18.VI.1911
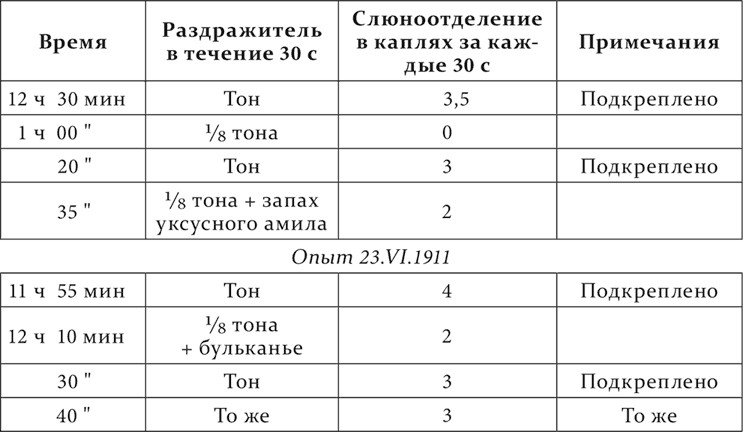
Интересно, что растормаживание можно констатировать и в фазе последействия дифференцировочного торможения.
Опыт Белякова на том же животном, у которого никогда не наблюдалось случайного слюноотделения.
Опыт 17.V.1911
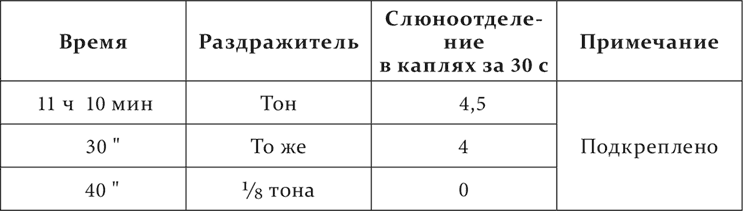
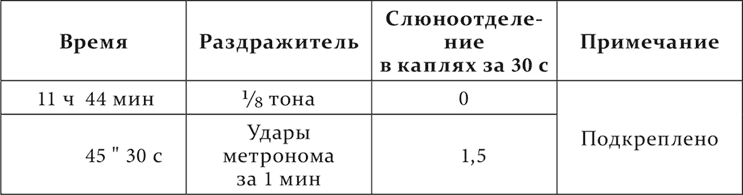
Между примененными нами посторонними раздражителями были и такие, которые вызывали не ориентировочную реакцию, но специальный рефлекс, более сложный и резкий. В таком случае растормаживающее действие давало себя знать еще долго по прекращении самого раздражителя.
Опыт Белякова на том же животном. Посторонним раздражителем была игрушечная труба, производившая резкие разнообразные и в высшей степени неровные звуки. Собака при этом лает, дрожит, рвется из станка.
Опыт 9.V.1911

Все сообщенные опыты делают несомненным, что выработка дифференцировок основана на развитии внутреннего торможения в отношении дифференцируемых агентов.
На основании разных фактов, описанных в последних лекциях, необходимо прийти к заключению, что между констатированием нервной системой разницы между внешними агентами вообще и дифференцированием тех же агентов при помощи условных рефлексов есть существенная разница. Первое обнаруживается раздражительным процессом в виде ориентировочной реакции, исследовательского рефлекса, только вторично влияющим на условные рефлексы или тормозящим или растормаживающим образом. Второе выражается в развитии тормозного процесса, являющегося в результате, так сказать, борьбы между раздражением и торможением. Как увидим это позже, эта борьба часто бывает очень трудна. Мыслимо, что иногда она и непосильна, и тогда при этой борьбе не всегда дело может доходить до полной утилизации для общей деятельности организма результата действительного анализа внешних агентов. Если это так, то изучение анализаторной деятельности нервной системы помощью условных рефлексов также будет иметь свой недостаток. Во всяком случае это – интересный вопрос.
Назад: Лекция шестая
Дальше: Лекция восьмая

