Книга: Лекции о работе больших полушарий головного мозга
Назад: Лекция пятнадцатая
Дальше: Лекция семнадцатая
Лекция шестнадцатая
Переходные фазы между бодрым состоянием и полным сном… животного (гипнотические фазы)
Мм. гг.! Прошлый год я привел внушительное количество фактов, доказывающих, что сон есть внутреннее торможение, сплошное (а не раздробленное, постоянно перемежающееся с процессом раздражения), распространившееся по всей массе полушарий и спустившееся также на некоторые нижележащие отделы головного мозга. Надо было ожидать, что, так как распространение торможения происходит постепенно, будет иметься различная экстенсивность сна, постепенное захватывание им то бóльших, то меньших районов. Следовательно, должны быть разные переходные формы к полному сну. Это и есть на самом деле, и мы наблюдали и производили их. При наших опытах мы имели дело не только с обыкновенной формой сна, характеризующейся при отсутствии нормальной деятельности полушарий расслаблением скелетной мускулатуры (закрытые глаза, низко опущенная голова, полусогнутые конечности и пассивно повисшее туловище на лямках, петлях для ног), но и с совершенно другой формой, что касается состояния скелетной мускулатуры. При ней деятельность полушарий также отсутствует, все условные раздражители остаются без действия, и на всякие посторонние раздражители, если они не достигают большой силы, тоже нет никакой реакции, но животное сохраняет вполне активную позу. Оно стоит с открытыми неподвижными глазами, с поднятой головой, на вытянутых конечностях, отнюдь не опираясь на петли для ног, стоит неподвижно минутами и часами. При изменении положения его конечностей оно удерживает то, которое им придано. Отдергивание лапы при прикосновении к подошвенной стороне ее принимает характер контрактуры. При подаче еды животное тоже не реагирует, оставаясь неподвижным, и еды не берет. Такая форма торможения встречается довольно редко, и мы пока не знаем, с какими специальными условиями в обстановке нашего экспериментирования или с какой особенностью нервной системы она связана. Наш сотрудник Н. А. Рожанский, тщательно наблюдавший у собак переход бодрого состояния в сонное, пришел к заключению, что описанное состояние существует всегда при этом, но обыкновенно как очень летучее, кратковременное явление. Физиологическое понимание этого состояния, как мне кажется, не представляет особенных затруднений. Перед нами заторможенная деятельность только больших полушарий, но торможение не спустилось ниже на центры, заведующие уравновешиванием, установкой тела в пространстве (центры Магнуса и Клейна), т. е. каталептическое состояние. Следовательно, в этой форме разграничительная линия между заторможенным отделом мозга и свободным от него лежит прямо под большими полушариями. Но она может разделять и большие районы самих полушарий. С этой новой формой мы встречаемся чаще и даже можем производить ее нарочно. Мы наблюдали ее впервые при следующих условиях (опыты Л. Н. Воскресенского). Одна из собак, с которой ранее работали без всякой помехи со стороны сна, вследствие того, что она в станке и одна в экспериментальной комнате была часто оставляема часами без всяких воздействий, начала впадать в сонное состояние. Очевидно, однообразные раздражения обстановки, как это указано в прошлой лекции, обусловили, наконец, сильное сплошное торможение мозга. Тормозящее действие обстановки сделалось таким сильным, что одно введение собаки в экспериментальную комнату сейчас же, видимо, делало ее другой, а тем более постановка в станок. Нужно было животное нарочно всячески раздражать, чтобы оно не оказалось заснувшим при окончании приготовлений к опыту. Когда затем экспериментатор уходил за дверь, чтобы оттуда вести опыт и сейчас же, не теряя минуты, начинал действовать условными пищевыми раздражителями, нормальный условный рефлекс был налицо: текла слюна, и собака тотчас принималась за подаваемую ей еду. Но если по выходе из комнаты пропускалось 4–5 минут, получался совершенно особенный результат: слюноотделительный эффект на условный раздражитель был, слюнная реакция при подаче еды увеличивалась, но животное еды не брало, и надо было ему вводить ее насильно. В это время расслабленной скелетной мускулатуры еще не было. Если по выходе из комнаты ждать 10 минут до применения условного раздражителя, то теперь уже на него не было никакого эффекта, и животное оказывалось вполне спящим, с расслабленной мускулатурой и храпом. Какое другое может быть объяснение этого факта кроме того, что при первоначальном распространении торможения одно время оно захватило только двигательную область коры, оставляя пока нетронутыми все остальные районы коры, откуда условные раздражения шли к органу (железе), не связанному с двигательной областью. И только несколько позже сплошное торможение оказалось на всей массе полушарий и спустилось на нижележащие отделы мозга, и вместе с этим наступил полный сон. В данном случае эта стадия развивающегося сонного состояния поручилась под влиянием индифферентных, долго действовавших на большие полушария раздражений. Обычно мы имеем ее при действии как отрицательных условных раздражителей, многократно повторяемых в одном и том же опыте, так и положительных условных раздражителей, специально слабых, тоже при частом и сплошном их применении. Привожу два примера.
«Сон есть внутреннее торможение, сплошное (а не раздробленное, постоянно перемежающееся с процессом раздражения), распространившееся по всей массе полушарий и спустившееся также на некоторые нижележащие отделы головного мозга»
Первый относится к собаке, упоминавшейся уже в ранних лекциях, у которой тон в 256 колебаний в секунду был условным пищевым раздражителем, а по 10 тонов вверх и вниз от него были отдифференцированы (опыты Бирмана).
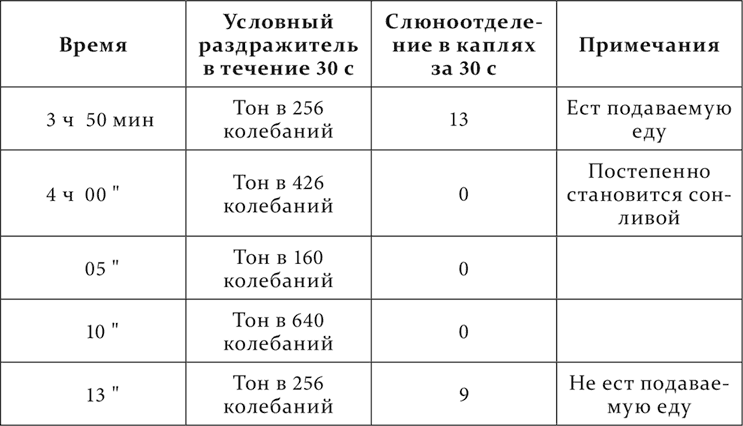
Другой пример беру из работы (Розенталя) над собакой, у которой сначала было много и постоянных условных пищевых рефлексов. Но когда у нее был выработан еще один рефлекс на появление перед ее глазами экрана из серой бумаги, и он повторялся много раз и часто сплошь один за другим в одном и том же опыте, то развилось следующее состояние собаки: часто на условные раздражители слюнная реакция была, и значительная, но к подаваемой еде собака не прикасалась. Вот такой пример.
При этом собака только малоподвижна, но еще нет никакого видимого сна. Без применения условных раздражителей и на том же станке собака с жадностью ест ту же еду.
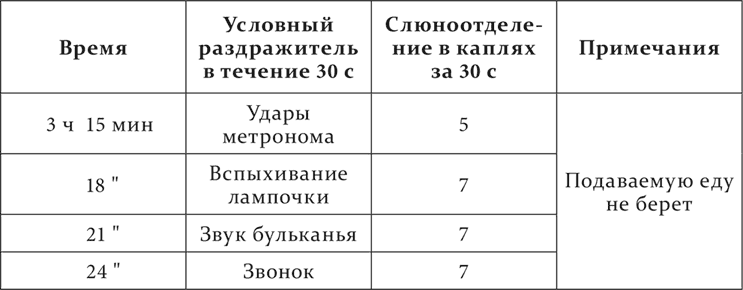
Сюда же относится и следующее случайное наблюдение. Одна из собак, давно служившая для опытов с условными рефлексами, никогда не представлявшая при условном пищевом рефлексе разъединения секреторной и двигательной реакций и никогда не спавшая в станке, была впервые поставлена перед многолюдной аудиторией для демонстрирования некоторых опытов с этими рефлексами. Необычная и очень сложная обстановка, видимо, оказала сильное действие на животное: оно оцепенело и слегка дрожало. При пробе условного раздражителя секреторный эффект получился, как обыкновенно, но поданную еду собака не взяла, а спустя довольно короткое время тут же в аудитории и в станке заснула, и с полным расслаблением скелетной мускулатуры. На этот раз, очевидно, сильный, необыкновенный раздражитель произвел прямо торможение больших полушарий – сперва частичное, только в районе двигательной области, а потом и полное с переходом на нижележащий отдел мозга. Этот случай надо считать совершенно тождественным с обычной формой опыта с так называемым гипнозом животных, когда стремительное обездвиживание животного и укладывание его на спину также ведет к торможению в различной степени его распространенности, то лишь к каталепсии, и даже частичной (неподвижность туловища при движении глаз и всей головы), то к полному сну. И у нас в лаборатории раз, когда попалось очень непокорное животное, сильно сопротивлявшееся его снаряжению к опыту, стремительное ограничение движений сильными руками, при причинении ему значительного механического раздражения, сейчас же повело к полному сну животного в станке.
«Так как распространение торможения происходит постепенно, имеется различная экстенсивность сна, постепенное захватывание им то бóльших, то меньших районов»
Таким образом, как частичный сон, так и полный производится как слабыми, долго продолжающимися общими раздражениями, так и короткими, но сильными, – как тормозными условными раздражителями, так и положительными. К некоторым подробностям, сюда относящимся, я еще не раз возвращусь в следующих лекциях.
Но рядом с различной экстенсивностью разлитого торможения наш фактический материал познакомил нас с различными вариациями, различными этапами самого процесса торможения, с различной, может быть, позволительно сказать, интенсивностью разлитого торможения, сна.
Но прежде чем обратиться к этому предмету, я должен предварительно коснуться пункта, имеющего существенное значение для многих наших опытов относительно вариации разлитого тормозного процесса. В восьмой лекции я ставил вопрос: на чем основано в одновременном комплексном раздражителе замаскирование раздражителя из одного анализатора раздражителем из другого анализатора, и высказал предположение, что, может быть, оно связано с различной силой употребляемых нами агентов, принадлежащих к разным анализаторам. Там проектированные опыты были исполнены в течение этих наших лекций и вполне подтвердили то предположение. Когда мы нарочито чрезвычайно изменили относительную интенсивность наших обычных раздражителей, звуковые очень ослабили, а другие или оставили, как они обыкновенно употреблялись, или усилили, то в эффекте комплексных раздражителей теперь, наоборот, оказались менее участвующими звуковые агенты, чем другие, т. е. при отдельных пробах условное действие принадлежало гораздо более вторым, чем первым.
Вот наши опыты. У одной собаки одновременное комплексное раздражение состояло из нашего обыкновенного кожно-механического раздражения и из звукового, чрезвычайно ослабленного (опыты В. В. Рикмана). Прочно выработанный раздражитель давал 4–4,5 капли слюны за 30 секунд изолированного действия. Отдельно испытанный звуковой компонент давал от 0,5 до 1,5 капли, а кожно-механический – от 2,5 до 5 капель. У другой собаки комплексное, также одновременное, раздражение составляли: ритмическое вспыхивание лампочки в 400 свечей и звучание чрезвычайно заглушенного тона. Выработанный комплексный раздражитель давал 7–8 капель слюны за 30 секунд. Отдельно примененный световой раздражитель давал 5 капель, звуковой – 2,5. Точно так же комбинированное раздражение температурой 0 °C, приложенной к коже, и очень слабым звуком при разложении на компоненты показало гораздо большее действие на стороне холода сравнительно с звуком (опыты W. Horsley Gannt и Купалова).
Таким образом, различие в величине эффекта наших обычных условных раздражителей, принадлежащих к различным анализаторам, обусловлено различной силой этих раздражителей, а не связано с качеством клеток разных анализаторов.
С этим положением мы можем приступить к нашей очередной теме об этапах разлитого тормозного процесса. Поводом к этому исследованию послужил случай патологического состояния нервной системы собаки под влиянием одного нашего приема не оперативного, а функционального характера. Экспериментально нами производимые патологические изменения нервной системы сами по себе займут наше внимание в следующей лекции. Здесь я опишу только исходный патологический опыт, из которого вытекало наше дальнейшее исследование на здоровых животных.
У собаки (опыты Разенкова) были выработаны положительные условные пищевые рефлексы на свисток, удары метронома, ритмическое механическое раздражение кожи (24 прикосновения в минуту), вспыхивание электрической лампочки, а также несколько дифференцировок, между ними дифференцировка на другую частоту прикосновений (12 в минуту) при механическом раздражении кожи на одном и том же месте.
Вот нормальное действие положительных условных раздражителей.
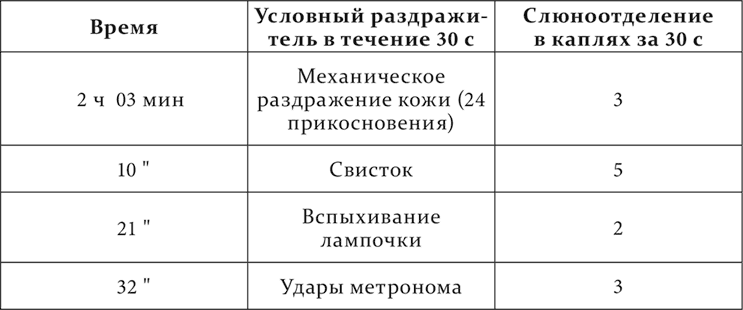
Следовательно, на основании предыдущего, раздражители по силе располагаются, идя от сильного к слабым, так: свисток, удары метронома, механическое раздражение кожи и вспыхивание лампочки.
Затем ставится такой опыт, что среди других раздражений применяется дифференцированное механическое раздражение (12 прикосновений в минуту) в течение 30 секунд, и оно непосредственно сменяется на положительное механическое раздражение (24 прикосновения в минуту) тоже в течение 30 секунд.
На другой день после этого опыта и в следующие 9 дней все условные рефлексы исчезают, только очень изредка проявляясь и в минимальном размере. За этим периодом наступает совершенно особенный. Вот он.
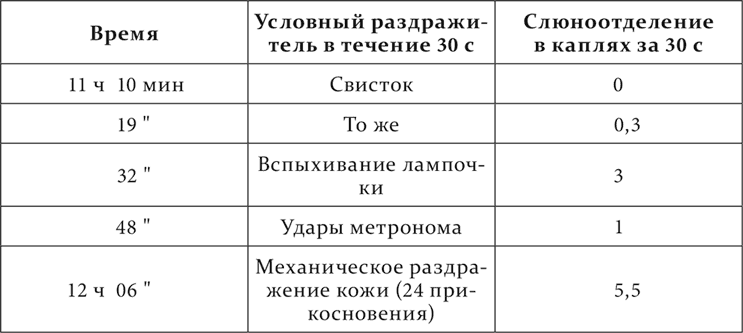
Получилось, как видите, совершенно обратное тому, что имелось раньше в норме: сильные раздражители совсем не действуют или едва, а слабые производят эффект даже больший сравнительно с нормой. Это состояние полушарий, по примеру Н. Е. Введенского, мы назвали парадоксальной фазой. Парадоксальная фаза держалась 14 дней и перешла в следующую такого вида:

Эту фазу мы назвали уравнительной фазой, так как при ней все раздражители оказывались равными по эффекту. Уравнительная фаза продолжалась 7 дней и сменилась еще на новую фазу, где очень усилились средние раздражители, сильный несколько упал, а слабый совсем остался без эффекта. 7 дней спустя вернулась норма. В этой работе, как и в наших дальнейших опытах относительно этого предмета, применялись ради большей бесспорности одни и те же раздражители, но разной интенсивности, так что было очевидно, что все дело в отношении клеток именно к разной силе раздражения.
Таким образом, мы в наших опытах впервые констатировали, что клетки полушарий переживают ряд переходных особенных состояний между нормально возбудимым и полным тормозным, обнаруживающихся в необычных отношениях этих клеток к различной силе раздражений.
Ознакомившись с этими переходными состояниями в явно патологическом случае, мы поставили себе дальше вопрос: не существуют ли эти состояния и в норме при переходе от бодрости ко сну и обратно? Представлялось вероятным, что в описанном случае патологическое состояние заключалось только в фиксировании надолго этих состояний, когда в норме они могли быть скоро проходящими, не бросающимися в глаза, как это оказалось с каталепсией. Наше исследование направилось по этой дороге и привело к утвердительному ответу. Вот ряд наших опытов.
Вот собака, уже упоминавшаяся в этой лекции раньше, у которой кроме одного положительно действующего тона было 20 отдифференцированных соседних тонов. Среди многих других положительных условных рефлексов у нее были выработаны дополнительно рефлексы на тихий и громкий треск, резко отличавшиеся по величине эффекта. Вот их нормальные отношения.

Опыт ведется дальше следующим образом. Повторным применением отдифференцированных тонов мы доводим собаку до явного засыпания и пускаем в действие слабый треск. Секреторного эффекта нет. Подачей еды мы пробуждаем собаку, она начинает есть. Через некоторое время слабый треск повторяется. Теперь он уже действует, но еще мало. Собака опять подкармливается. В третий раз слабый треск вызывает нормальный эффект и даже иногда несколько больший. Происходит опять подкрепление. В следующий раз применяется сильный треск. Его эффект более или менее уступает по величине эффекту перед этим действовавшего слабого треска. И только несколько позже при полном восстановлении бодрого состояния сильный треск достигает его полного нормального эффекта, и с этим вообще появляются обычные количественные отношения между этими двумя раздражителями. Вот подлинные цифры одного из таких опытов.

Затем собака усыпляется применением отдифференцированных тонов.

Иногда при таких опытах наблюдается при повторении этих раздражений, вместо первоначального преобладания в отношении эффекта слабого треска перед сильным, их равенство. Очевидно, при постепенном рассеивании сонного состояния под влиянием повторяющегося короткого акта еды корковая клетка, возвращаясь к бодрому состоянию, проходит и теперь как парадоксальную, так и уравнительную фазы. Следовательно, то же самое, что в патологическом случае; только то, что там держалось днями, здесь меняется минутами (опыт Бирмана).
«В одновременном комплексном раздражителе замаскирование раздражителя из одного анализатора раздражителем из другого анализатора связано с различной силой употребляемых агентов, принадлежащих к разным анализаторам»
У другой нашей собаки, вследствие продолжительной работы с ней, развилось легкое дремотное состояние на станке, отразившееся на условных рефлексах в том, что ранее отчетливо отличавшиеся различные раздражители в отношении величины их условного эффекта теперь все сравнялись. Благодаря впрыскиваниям соответствующей дозы кофеина собака приходила в первоначальное бодрое состояние, а с ним восстановлялись и нормальные отношения между условными раздражителями (опыты Н. В. 3Имкина).

На другой день за 18 минут до опыта собаке впрыскивается под кожу 8 см³ 2 % раствора coffeini puri: собака совершенно бодра.

У собаки, которая уже упоминалась в этой лекции как представлявшая стадию разъединения секреторной реакции от двигательной, часто в этой стадии наблюдалось, что из всех раздражителей только самый слабый (вспыхивание лампочки) действовал иногда всего более положительно, вызывая полный нормальный рефлекс: текла слюна, и собака ела подаваемую ей еду. Таким образом, в таких случаях с определенной степенью экстенсивности разлитого торможения существовала и парадоксальная фаза (опыты Розенталя).
При явном сонливом состоянии, но еще до наступления полного сна, замечалось еще следующее совершенно особенное явление. Когда условный положительный раздражитель терял совсем, или почти, свое действие, хорошо выработанный отрицательный раздражитель, наоборот, получал отчетливое положительное действие. Вот пример (опыты Шишло).
«Основная и самая общая деятельность больших полушарий есть сигнальная с бесчисленным количеством сигналов и с переменной сигнализацией»
У собаки были выработаны пищевые положительные условные рефлексы на механическое раздражение кожи плеча и бедра и на приложение к коже тепла 45 °C, а также постоянный отрицательный условный рефлекс на механическое раздражение кожи на спине. Эффект положительного механического раздражения кожи достигал в норме 15–18 капель в минуту. Температурный условный раздражитель довольно скоро начал вызывать сонливость и сон. В данном опыте сперва применяется температурный раздражитель. Развивается сонливость. Дальше опыт идет следующим образом.
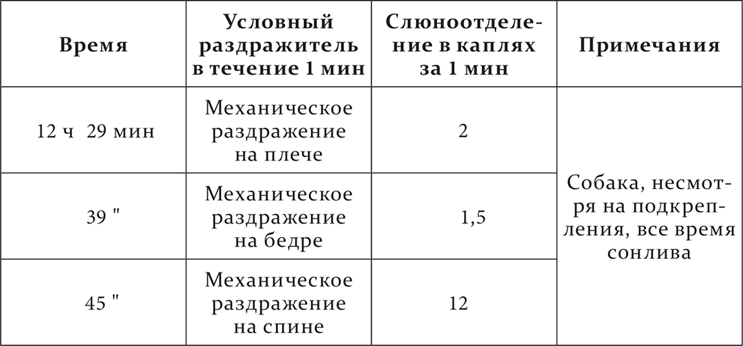
То же мы не один раз наблюдали и при некоторых патологических состояниях полушарий. Это состояние мы назвали ультрапарадоксальной фазой.
Наблюдая столько различных состояний клеток полушарий при переходе животного от бодрого состояния к полному сну и зная, что сон есть сплошное и разлитое внутреннее торможение, мы должны были ожидать, что некоторые из этих состояний мы встретим и в так называемом нами последовательном торможении, с которым мы основательно познакомились в ранних лекциях о внутреннем торможении. Как кажется, это и было, пока в единственном случае, который мы успели исследовать, именно при условном торможении (опыты Быкова).
У собаки имелись пять положительных условных раздражителей: удары метронома, громкий тон, он же сильно заглушенный, появление перед глазами собаки круга из картона и механическое раздражение кожи. Условное торможение было выработано на комбинацию механического раздражения кожи со звуком бульканья. Условные раздражители по величине слюноотделительного эффекта, средней из многих опытов, располагались в том порядке, как они перечислены: 22 капли, 18,5, 16,5, 13,5 и 10 капель за 30 секунд. Когда условное торможение только что было окончательно выработано, испытывались все условные раздражители. 10 минут спустя после того, как было применено условное торможение, удары метронома давали 16,5 капли, сильный тон 16, заглушенный тон 20 капель и круг 18 капель. Принимая во внимание возможное участие в факте как движения торможения, так и индукции, единственное, что можно было отнести на счет интересующего нас пункта, это то, что заглушенный тон действовал больше нормы, когда эффект сильного был менее нормы. Так как это происходило в одном и том же пункте коры, то в этом можно было видеть обнаружение парадоксальной фазы. Сейчас мы продолжаем это исследование дальше на других видах внутреннего торможения.
Затем мы остановились на следующем пункте. В лекции о взаимной индукции мы сделали предположение: не есть ли так называемое нами внешнее торможение явление отрицательной индукции, т. е. торможение, индуцированное на периферии района, находящегося в состоянии раздражения, иначе говоря, не есть ли внешнее и внутреннее торможение в своей физико-химической основе один и тот же процесс? Мы рассчитывали получить, может быть, некоторое подтверждение этого предположения при исследовании вопроса: не обусловливает ли внешнее торможение те же состояния корковой клетки, с которыми мы только что познакомились в случаях внутреннего торможения? Так как для этого исследования нужен был более или менее продолжительный период внешнего торможения, то мы производили его введением в рот животного различных отвергаемых веществ, при которых внешнее торможение дает себя знать очень долго.
Опыты произведены на двух собаках с пищевыми рефлексами.
У одной собаки (опыты Пророкова) после вливания ей в рот раствора соды, когда вызванное этим слюноотделение прекратилось, испытанные сейчас же как сильные, так и слабые условные раздражители были одинаково сильно заторможены, но позднее в ближайшие 15–20 минут слабые уже действовали как в норме, так и даже больше и были или равны по действию с сильными, или даже в значительной степени превосходили их, если они были теперь еще очень ослаблены. Вот пример. В 9 часов 41 минуту влит раствор соды.

Обыкновенный эффект электрического звонка около 8 капель за 30 секунд, эффект механического раздражения кожи около 4 капель.
У другой собаки (опыты П. К. Анохина) результат опытов частью согласовался с результатом, полученным на первой собаке, частью представлялся особенным. После введения отвергаемых веществ в рот собаки и прекращения вызванного ими слюноотделения сейчас же и дальше до окончания опыта все раздражители, ранее постоянно различавшиеся между собой по величине эффекта (сильный звонок был первым раздражителем по эффекту, свет лампы – последним), сравнивались в этом отношении. Но рядом с этим происходило ступенеобразное падение рефлексов в течение опыта. Вот как шел опыт.
Влит раствор соды. Слюноотделение продолжалось 10 минут.

Хотя полученный на обеих собаках результат в общем и благоприятен предположению, что внутреннее и внешнее торможение в своей основе является одним и тем же процессом, но сложность явлений требует повторения и варьирования этих опытов при более строгом внимании к другим возможным толкованиям фактов.
Наконец, нам представлялось интересным исследовать: как относятся наши условные рефлексы при действии снотворных средств от начала этого действия до полного усыпления животного и обратно до возврата к бодрому состоянию. Мы употребляли с этой целью уретан и хлоралгидрат. Здесь почти исключительно преобладал другой ход явлений, именно постепенное падение всех рефлексов, так что слабые раздражители естественно ранее делались недействительными, чем сильные. Мы назвали такое состояние клеток наркотической фазой. Привожу один из этих опытов (С. И. Лебединской).
У собаки имелись следующие положительные условные раздражители: сильный звонок, удары метронома, тихий звонок, механическое раздражение кожи и прерывистое вспыхивание электрической лампочки перед глазами собаки. По эффекту раздражители располагались, как они перечислены. В 10 часов 9 минут введено в rectum 2,0 г хлоралгидрата в 150 см³ воды. Собака стоит в станке. Опыт идет дальше так.
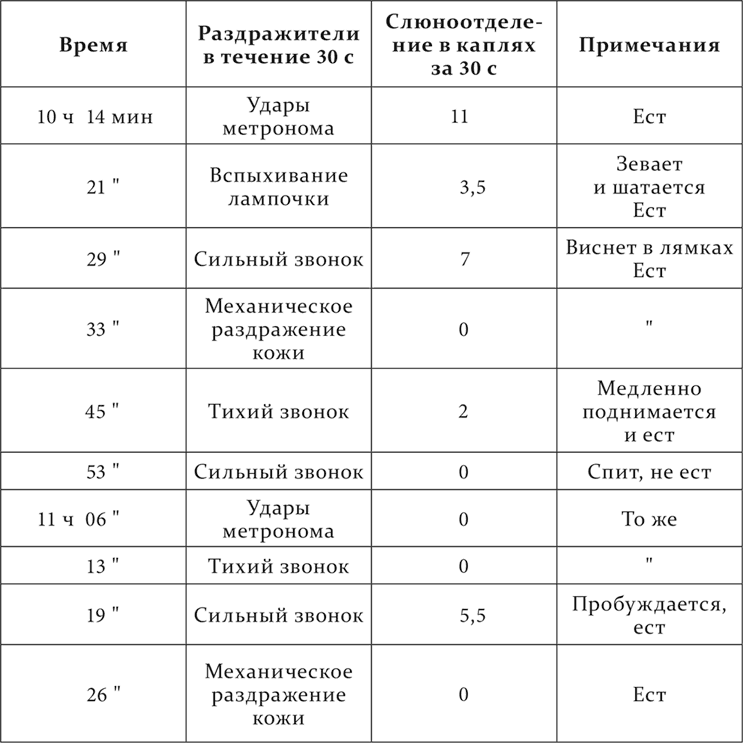
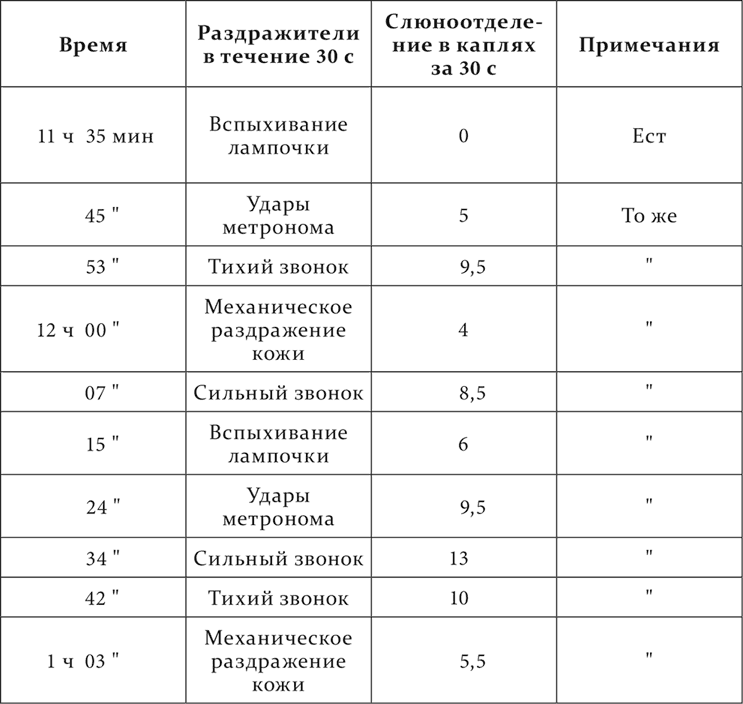
Мы видим, что при развитии снотворного действия все раздражители постепенно теряли в их эффекте, а при возврате к бодрому состоянию также все и постепенно достигали нормальной величины их эффекта. Единственное резкое исключение из 20 раздражений представил тихий звонок, произведший в 11 часов 53 минуты несоответственно большое действие.
«Парадоксальная фаза – состояние полушарий, когда сильные раздражители совсем не действуют или едва, а слабые производят эффект даже больший сравнительно с нормой. Уравнительная фаза: все раздражители равны по эффекту. Ультрапарадоксальная фаза: постоянная сонливость. Наркотическая фаза: постепенное падение всех рефлексов, так что слабые раздражители естественно ранее делаются недействительными, чем сильные»
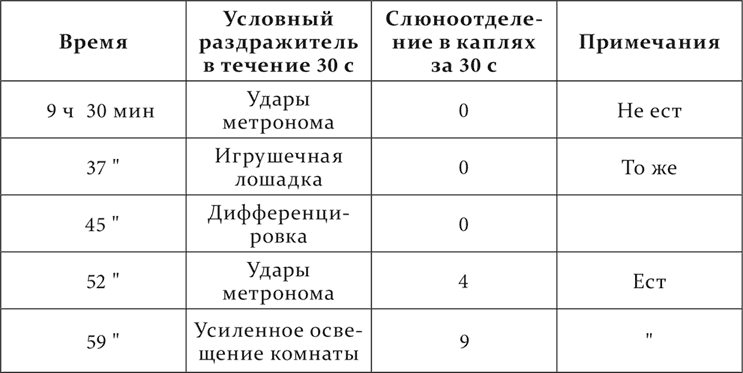
Итак, на разных здоровых животных и при разных условиях мы получили много разных состояний полушарий в отношении реакции их на условные раздражители. Поднимался вопрос: насколько все эти состояния, включая сюда и наркотическую фазу, свойственны каждому животному и при обычных условиях жизни? При решении этого вопроса мы оказались в счастливом положении. Среди наших собак имелась одна (на ней были поставлены опыты, приведенные в конце четырнадцатой лекции), принадлежащая к резкому нервному типу, о котором будет сообщено в следующей лекции. Эта собака отличалась часто прямо изумительной при определенных условиях стереотипностью ее высшей нервной деятельности, в виде условных рефлексов. Заслуженно собака много раз получала от нас название живого инструмента. У нее, как было сообщено раньше, было 10 условных рефлексов: 6 положительных – на звонок, удары метронома, свисток, усиление общего освещения комнаты, появление перед ее глазами формы круга и игрушечной лошадки, и 4 отрицательных – на другую частоту ударов метронома, уменьшение общего освещения комнаты, форму квадрата и игрушечного зайчика приблизительно того же размера и цвета, как лошадка. В последнее время по некоторым основаниям не употреблялся звонок, а из отрицательных мы пользовались почти только дифференцировкой ударов метронома. Звуковые раздражители обычно, а в ранних опытах постоянно, производили гораздо больший (на 30–50 %) слюноотделительный эффект, чем световые. После долгой лабораторной службы собаки (больше 2 лет) положительные условные раздражители стали обнаруживать тенденцию к уменьшению размера их действия и к изменению количественных отношений между ними, как это часто происходит с нашими собаками при долговременном пользовании одними и теми же условными раздражителями. И теперь-то мы могли видеть на нашей собаке в отчетливой форме все те состояния полушарий, которые описаны раньше в этой лекции как вариации наступающего разлитого тормозного процесса. Каждое такое состояние полушарий или резко выступало в продолжение целого опыта, или же то само собой, то под влиянием наших мер переходило в другие (опыты Сперанского). Единственно, что у нее не наблюдалось, – это ультрапарадоксальная фаза. Но не представлялось и случая для этого, так как собака никогда не становилась отчетливо сонливой. Вот опыты из разных дней и разных периодов.
Нормальный опыт
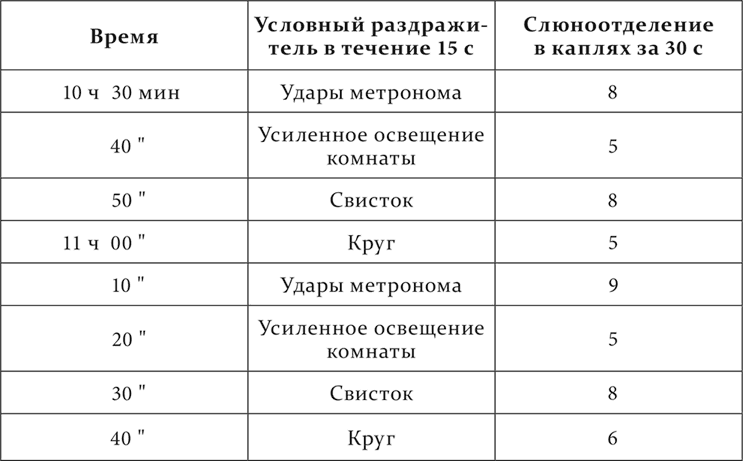
Опыт с уравнительной фазой
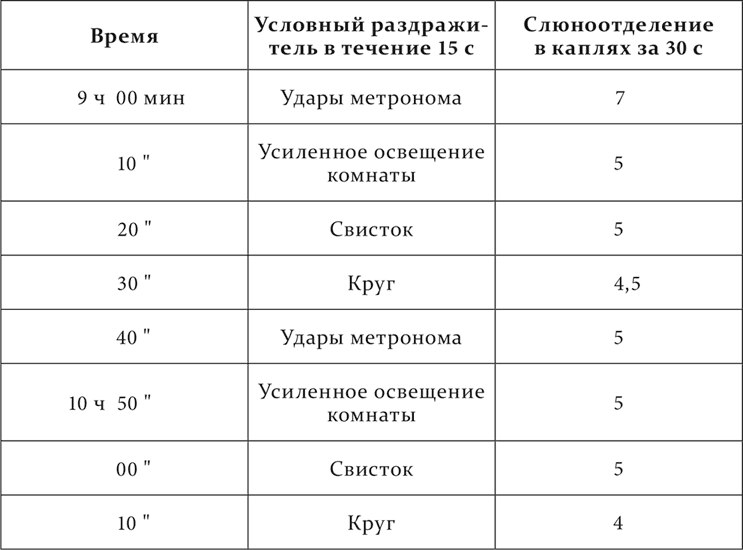
Опыт с парадоксальной фазой, переходящей в нормальную
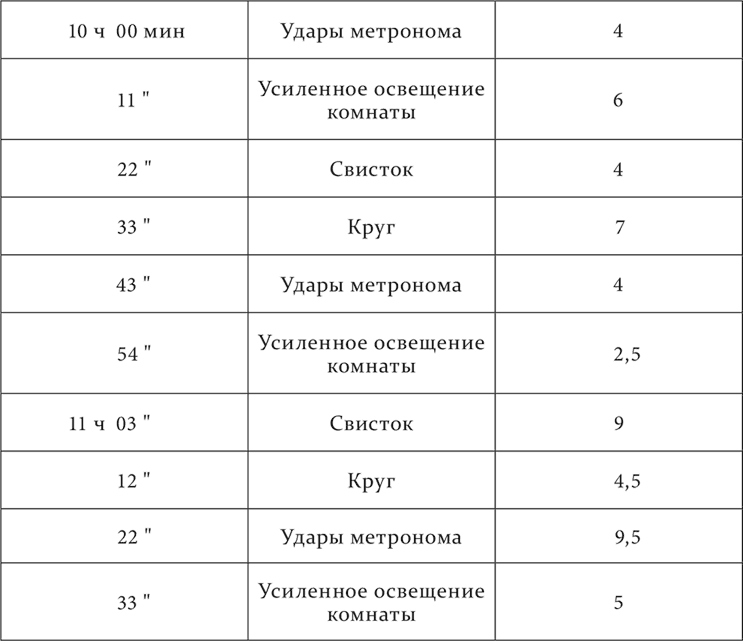
Опыт с полным торможением, переходящим в наркотическую фазу
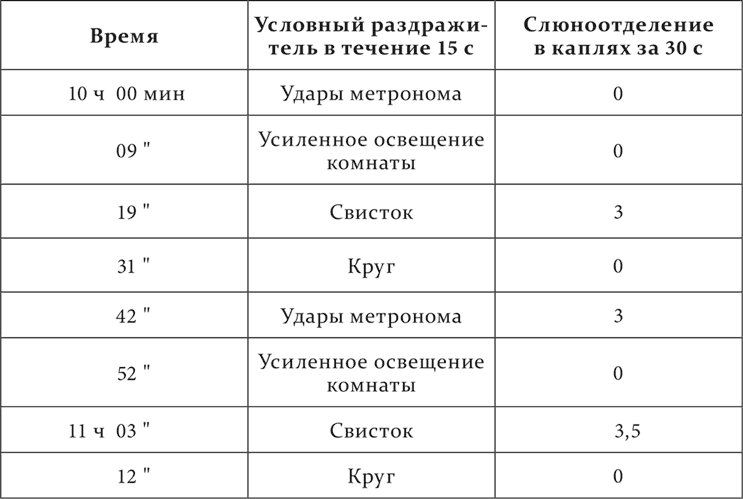
Когда рефлексы очень падали и искажались, мы поднимали и исправляли их, как указано было в четырнадцатой лекции, тем, что действовали условными раздражителями до присоединения безусловного более короткое время. Отсюда в приведенных там опытах различная продолжительность изолированного условного раздражения. Переход из одной фазы в другую в двух последних опытах, надо думать, происходил под влиянием повторяющегося акта еды. Но мы сами располагали двумя специальными приемами, которыми сейчас же могли сменять фазы. Это – применение всегда полной дифференцировки (на другую частоту ударов метронома), как агента, концентрирующего торможение или индуцирующего процесс раздражения, а также применение социального раздражителя, которым являлось присутствие в экспериментальной комнате хозяина собаки – экспериментатора. Вот примеры.
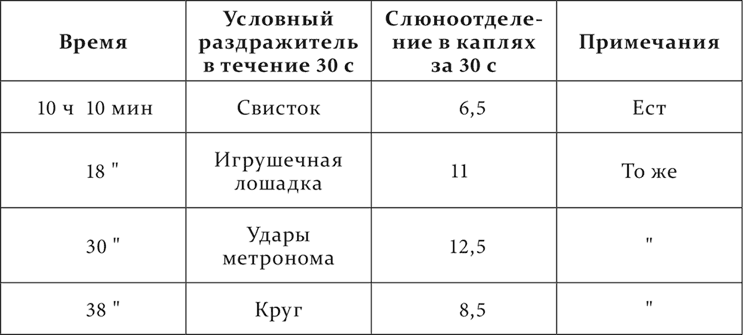
Полная тормозная фаза (нет ни секреторной, ни двигательной реакции) после применения дифференцировки переходит сперва в парадоксальную, а потом и в нормальную фазу.
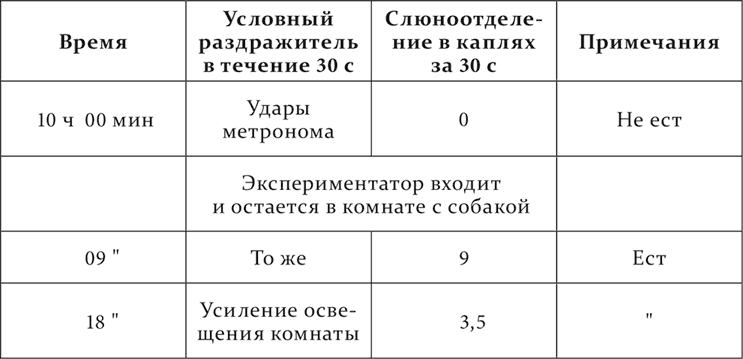
Присутствие экспериментатора в комнате с собакой сейчас же превращает полную тормозную фазу в нормальную.
Вопрос: можно ли перечисленные переходные фазы в состоянии полушарий расположить в один ряд, и если можно, то в какой именно, остается пока для нас полностью нерешенным. Если взять все наши случаи, преемственность фаз оказывается довольно разнообразной. Таким образом, остается неясным: есть ли эти состояния строго последовательные или параллельные. Мы также не можем точно указать, почему данная фаза прямо переходит то в ту, то в другую. Следовательно, требуется дальнейшее исследование.
Едва ли можно сомневаться в том, что описанные в этой лекции состояния полушарий есть то, что называется гипнозом в его разнообразных стадиях и чертах. Подробно о явлениях человеческого гипноза в связи с полученными нами фактами будет речь в последней лекции.
Назад: Лекция пятнадцатая
Дальше: Лекция семнадцатая

