Книга: Лекции о работе больших полушарий головного мозга
Назад: Лекция четырнадцатая
Дальше: Лекция шестнадцатая
Лекция пятнадцатая
Внутреннее торможение и сон – один и тот же процесс…… в своей физико-химической основе
Мм. гг.! В прошлой лекции мы пришли к очень важному положению, что корковая клетка под влиянием условных раздражений непременно рано или поздно, а при частых повторениях их и очень быстро, приходит в тормозное состояние. А это всего законнее надо было понимать так, что эта клетка, как, так сказать, сторожевой пункт организма, владеет высшей реактивностью, а следовательно, стремительной функциональной разрушаемостью, быстрой утомляемостью. Наступающее тогда торможение, не будучи само утомлением, является в роли охранителя клетки, предупреждающего дальнейшее чрезмерное, опасное разрушение этой исключительной клетки. За время тормозного периода, оставаясь свободной от работы, клетка восстановляет свой нормальный состав. Это касается всех клеток коры, и, следовательно, при условии множества работавших клеток коры, вся кора должна приходить в то же тормозное состояние, которое мы видели в отдельных клетках коры, когда на них действовали наши условные раздражители. А это и есть каждодневный факт, есть сон наш и всех животных. Вся наша двадцатипятилетняя работа над большими полушариями является сплошным и постоянным доказательством этого заключения. Сейчас в физиологии больших полушарий, как мы ее изучаем нашим методом, это одно из наиболее прочных положений. Сонливость и сон наших экспериментальных животных сопровождали нашу работу с самого ее начала, и до сих пор мы постоянно имеем с ними дело. Таким образом, нами собран огромный фактический материал, который в разные фазы нашего исследования, конечно, давал основание для многих предположений, несколько разнящихся друг от друга. Но уже несколько лет тому назад все эти предположения слились в окончательное, гармонирующее решительно со всеми нашими фактами положение, что сон и то, что мы называем внутренним торможением, есть один и тот же процесс.
Основное условие появления и развития как этого торможения, так и сна – совершенно одно и то же. Это более или менее продолжительное или много раз повторяющееся изолированное условное раздражение, т. е. раздражение корковой клетки. При всех случаях внутреннего торможения, с которыми мы знакомились в четвертой, пятой, шестой и седьмой лекциях, мы постоянно встречались с сонливостью и сном наших животных. Если мы производим угасание, то у иного животного и при первом случае с ним, помимо прекращения условного слюноотделения и соответствующей двигательной реакции, уже замечается бóльшая вялость животного сравнительно с состоянием до угасания. При повторении угасания в течение ряда дней, хотя бы при чередовании с подкрепляемыми условными раздражителями, почти всегда дело кончается явной сонливостью и сном на станке, хотя до этого с животным этого не случалось. То же самое еще резче наблюдается при выработке дифференцировок. Мы, например, имеем у животного ряд условных раздражителей и между другими определенный тон. Животное все время остается на станке бодрым. Затем мы переходим к дифференцировке одного из близких тонов. Вместе с начинающим развиваться дифференцировочным торможением появляется сонливость, все усиливающаяся и кончающаяся глубоким сном с полным расслаблением скелетной мускулатуры и храпом животного, так что при следующих положительных условных раздражителях и подаче еды приходится расталкивать животное или даже вкладывать ему пищу в рот для начала акта еды. С тем же самым встречаемся и при образовании сильно запаздывающего отставленного рефлекса (на 3 минуты), так что раньше, когда мы еще мало были знакомы с делом, из-за возникающего при этом сна у иных животных нельзя было добиться выработки нужного нам рефлекса. Наконец, то же имеет место и при выработке условного торможения. Надо заметить, что при разных случаях внутреннего торможения степень вмешательства сна несколько различна. Всего меньше он дает себя знать именно при условном торможении.
«Вся наша двадцатипятилетняя работа над большими полушариями является сплошным и постоянным доказательством заключения о том, что сон наш и всех животных есть тормозное состояние всей коры»
Это все случаи, где внутреннее торможение развивается быстро вследствие того, что условный раздражитель не сопровождается безусловным раздражителем, но то же самое происходит и при медленно нарастающем торможении подкрепляемых, но много раз повторенных условных рефлексов, торможении, о котором специально была речь в прошлой лекции. И тут через месяцы и годы дело кончается разными переходными состояниями между бодрым состоянием и сном (что составит содержание следующей лекции) и полным сном, смотря по животному. В этом отношении животные также разнятся между собой, как и в отношении скорости наступления торможения.
Случаями перехода торможения в сон так переполнена наша работа с условными рефлексами, что является излишним приводить отдельные примеры. Таким образом, действительно торможение тесно связано со сном, прямо переходит в сон, если не принимаются соответствующие меры.
Интересно, что как для внутреннего торможения почти нет раздражителя, который бы не подвергся ему при определенных условиях, так и для сна. Сильнейший электрический ток, приложенный к коже в качестве условного пищевого раздражителя, в опытах Ерофеевой обнаружил после многих месяцев нарастающее торможение при подкрепляемых рефлексах, а в опытах Петровой демонстративно вызывал сон. С другой стороны, различные внешние агенты в роли условных раздражителей совершенно одинаково располагаются в отношении скорости развития при них как внутреннего торможения, так и сна. В прошлой лекции было сообщено, что всего легче развивается торможение при температурных раздражениях и всего труднее при звуковых. Строго параллельно с этим сон быстро наступает при температурных условных рефлексах и позже и реже при звуковых. При температурных условных раздражениях вмешательство сна так настойчиво и так нарушает работу, что в первую пору наших исследований трудно было найти сотрудников, соглашавшихся работать с этими раздражителями.
Наконец, как раньше было указано, для торможения и для наступления сна определяющее значение имеет продолжительность условного раздражения. У иной собаки, пока условный раздражитель отставлен на 10–15 секунд, за длинный период работы сохраняется совершенно бодрое состояние. Лишь только рефлекс у нее отставляется на 30 секунд и больше, сейчас же появляются сонливость и сон. Эта форма опыта часто поистине поразительна. Кажется совершенно неожиданным этот быстрый переход от полной бодрости к настоящему сну при таком, по-видимому, малозначительном изменении условий опыта. Таких примеров при разной длине отставления и с разной степенью отчетливости рассеяно по нашей работе множество.
«Наступающее торможение, не будучи само утомлением, является в роли охранителя корковой клетки, предупреждающего дальнейшее чрезмерное, опасное разрушение этой исключительной клетки»
Все без исключения приемы, которые были приведены в прошлой лекции как задерживающие наступление нарастающего торможения давно повторяющихся, хотя и постоянно подкрепляемых условных рефлексов, так и устраняющие торможение, служат одинаково и для борьбы со сном.
После всего только что приведенного поднимается естественнейший вопрос. Если сон так совпадает в его появлении и в его исчезновении с внутренним торможением, то каким образом последнее может быть важнейшим фактором бодрого состояния, основанием тончайшего уравновешивания организма с окружающей средой? Сообщенные в ранних лекциях факты должны, по моему мнению, совершенно устранить кажущуюся, может быть, на первый взгляд трудность ответа. Внутреннее торможение в бодром состоянии есть раздробленный сон, сон отдельных групп клеток, как сон есть внутреннее торможение, иррадиированное, распространившееся сплошь на всю массу полушария и на лежащие ниже отделы головного мозга. Все дело, следовательно, состоит в пространственном ограничении торможения, в заключении его в определенные рамки. А делает это, конечно, противоположный нервный процесс, как это мы видели в лекциях о мозаичности коры и об анализаторной деятельности ее.
«Корковая клетка, как сторожевой пункт организма, владеет высшей реактивностью, а следовательно, стремительной функциональной разрушаемостью, быстрой утомляемостью»
При угасании сон не наступает, не допускается только тогда, когда после достигнутого угасания мы систематически применяем подкрепляемые условные раздражители и не повторяем угасание слишком часто. При дифференцировках развивающееся торможение, сначала сопровождаемое сном, остается, так оказать, деловым торможением, без осложнения неуместным сном, только тогда, когда вперемежку с неподкрепляемым агентом применяется, и обыкновенно чаще, наш условный раздражитель, положительно действующий агент, т. е. процесс раздражения постоянно противодействует распространению процесса торможения.
То же самое при условном торможении и при запаздывании. Во всех этих случаях, если опыт ведется целесообразно, сонливость и сон являются только как фазовое явление, когда еще не установилось точное разграничение между районами раздражительного и тормозного процессов. Как только условия опыта дают перевес торможению, наступает сон. Вот резкий пример. В лекции о мозаике я упоминал о собаке, у которой на определенный тон был выработан положительный условный рефлекс, а 20 соседних с ним тонов вверх и вниз были отдифференцированы, т. е. были отрицательными условными раздражителями. Собака вообще была очень не склонна ко сну, когда соблюдалось определенное отношение при применении положительных и отрицательных раздражителей, и всегда тогда давала полные эффекты на все положительные раздражители. Но как только несколько раз подряд пускались в действие отдифференцированные тоны, собака скоро впадала в такой глубокий сон, что ее не будили очень сильные посторонние раздражители. Наоборот, сон не наступает, несмотря и на частое применение отрицательных условных раздражений, если они чередуются с положительными. В этом отношении очень поучительны уже упоминаемые ранее опыты с мозаикой и особенно опыты с кожно-механической мозаикой. Несмотря на то что условное кожно-механическое раздражение очень располагает к сонливости и сну, тем не менее собака Купалова, на которой более двух лет практикуется исключительно кожно-механическая мозаика, никакой наклонности ко сну не обнаруживает, очевидно, потому, что тормозной процесс постоянно отмежевывается, заключается в узкие рамки раздражительным процессом. Обширно иррадиирующему торможению мы противодействуем еще и тем, что увеличиваем число положительных условных раздражителей, ограничивающих распространение торможения из его исходных пунктов. Приведу более сложный, отчасти сюда относящийся, пример из работы Петровой. У собаки было приступлено сразу к выработке сильно отставленного (на 3 минуты) пищевого рефлекса на удары метронома. Собака очень скоро стала впадать в сонливое состояние и кончила полным сном. Очевидно, торможение, которое должно было развиться на первом периоде действия метронома, как очень удаленном от времени присоединения безусловного раздражителя, взяло перевес, не встречая должного противодействия со стороны еще не проявившегося процесса раздражения на втором периоде действия метронома, как непосредственно предшествующем действию безусловного раздражителя. Тогда было введено еще 5 новых агентов для выработки из них условных раздражителей, причем еда присоединялась к ним спустя только 5 секунд после их начала. Сонливое состояние быстро исчезло, и все рефлексы легко образовались, затем началось постепенное, каждый день на 5 секунд, отодвигание акта еды от начала условных раздражителей. Соответственно удлинялся и латентный период рефлексов, и в конце, без малейшего вмешательства сна, получилось 6 рефлексов с латентным периодом в 1–1,5 минуты, т. е. с предварительным тормозным процессом. Таким образом, процесс раздражения, первоначально возникший в 6 пунктах коры и только постепенно давая место процессу торможения, ограничил его и во времени и в пространстве, не допустив сна.
«За время тормозного периода, оставаясь свободной от работы, клетка восстановляет свой нормальный состав. Это касается всех клеток коры»
Сюда же надо отнести и влияние на некоторых, правда немногих собак, особенно наклонных к торможению и сонливости, ограничения движений в нашем станке. Тогда иногда целесообразно вести опыты, по крайней мере временно, на полу, на свободе. Надо думать, что раздражения, идущие при движении из двигательного аппарата и с кожи, образуют переменно возникающие очаги возбуждения в коре полушарий, которые также до известной степени противодействуют распространению торможения, хотя тут есть и другой момент, вероятно, большего значения, о чем речь впереди.
То, что мы видели на корковой клетке относительно появления в ней торможения в случаях условного раздражения, повторяется на ней и при раздражении ее всяким агентом, не имеющим никакого условного специального физиологического значения. Как уже упомянуто раньше, между рефлексами имеется исследовательский рефлекс. Этот рефлекс имеет своими пунктами приложения клетки как больших полушарий, так и нижележащих отделов головного мозга. При наличии больших полушарий он, очевидно, происходит при участии клеток больших полушарий. Это несомненно доказывается его высочайшей тонкостью, когда он появляется при всяком малейшем колебании окружающей среды, что возможно только при высшей анализаторной функции больших полушарий и что совершенно недоступно для нижележащих отделов мозга. Исследовательский рефлекс, как все мы знаем, при повторении непременно слабеет и, наконец, совершенно исчезает, несмотря на продолжение существования агента, его вызвавшего. Специальные опыты в нашей лаборатории Н. А. Попова показали, что исчезание исследовательского рефлекса основано на развитии торможения и вполне аналогично в отношении подробностей угасанию условных рефлексов.
Если повторяемый через короткие промежутки времени в течение опыта данный агент исследовательского рефлекса перестает вызывать соответствующую двигательную реакцию, то при большем промежутке в том же опыте эта реакция восстанавливается, как это делается с угашенным условным рефлексом. Исчезнувший в опыте от повторения определенный исследовательский рефлекс на время появляется, если сейчас же после его исчезания применяется новый посторонний раздражитель, т. е. производится другой исследовательский рефлекс. Следовательно, он растормаживается, как и задержанный условный рефлекс. Если исследовательский рефлекс на определенный агент повторяется в течение ряда дней, то он хронически исчезает, как систематически неподкрепляемый условный рефлекс. Наконец, исчезнувший исследовательский рефлекс временно восстанавливается под влиянием возбуждающих веществ (кофеина), как это наблюдается, например, при условных дифференцированных агентах. Это торможение исследовательского рефлекса непременно, и даже легче, чем торможение условного рефлекса, ведет к сонливости и сну. Привожу опыты из работы С. И. Чечулина, в которой торможение и сон при исследовательском рефлексе комбинировались с применением условных раздражителей.
У собаки выработан пищевой условный рефлекс на свисток. В качестве возбудителей исследовательского рефлекса употребляются впервые применяемые агенты: шипение, бульканье, механическое раздражение кожи и другие.
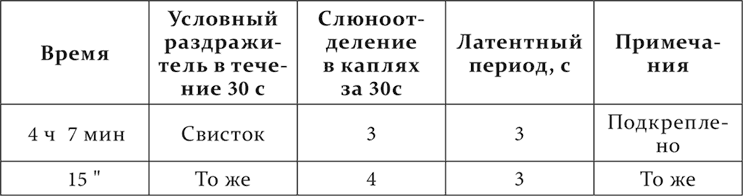
Затем, начиная с двадцать первой минуты, применяется в течение 30 секунд и повторяется через каждые 2 минуты звук бульканья. При первых трех повторениях наступают движения в сторону звука, постепенно ослабляющиеся. С четвертого обнаруживается сонливость. До восьмого повторения сон прерывается на разные моменты раздражения. При восьмом и девятом нет более никаких движений на раздражитель, сон сплошной. В 43 минуты сперва одно бульканье применяется 10 секунд, а затем на 30 секунд к нему присоединяется свисток. Нет никакой – ни двигательной, ни секреторной – реакции, продолжается сон. Подача еды пробуждает собаку, собака начинает есть, но и после еды остается сонливой. Опыт с условным раздражителем продолжается следующим образом.
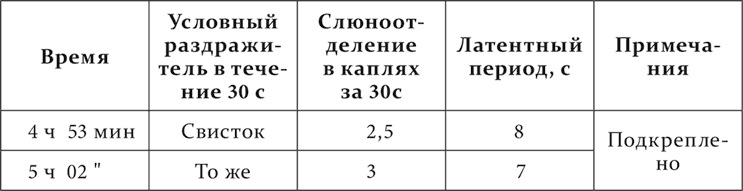
Собака до этого опыта в станке никогда не спала. В следующих опытах новые агенты, вызывающие исследовательский рефлекс, то повторялись до наступления сна, то только до тех пор, пока не исчезали на них движения, но еще до появления сонливости. 21 день спустя после вышеприведенного опыта ставится опыт с механическим раздражением кожи. Опыт идет следующим образом.
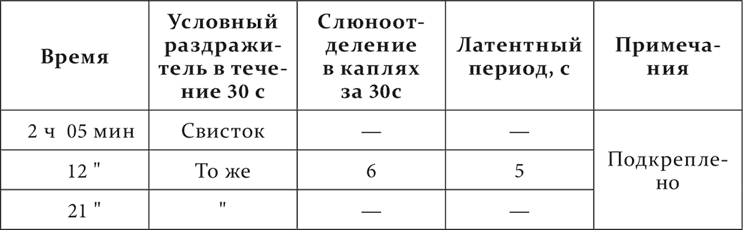
С двадцать пятой минуты начинается механическое раздражение кожи, продолжающееся 30 секунд каждый раз и повторяемое через минуту. При первых трех повторениях движения головой в сторону раздражаемого места. При четвертом и пятом повторениях движений больше нет. В 32½ минуты на 10 секунд применяется одно механическое раздражение кожи, а затем в продолжение 30 секунд к нему присоединяется свисток. На пятнадцатой секунде от начала свистка начинается слюноотделение, и за 30 секунд присоединения свистка выделяются 2 капли. Дальше опыт идет так.
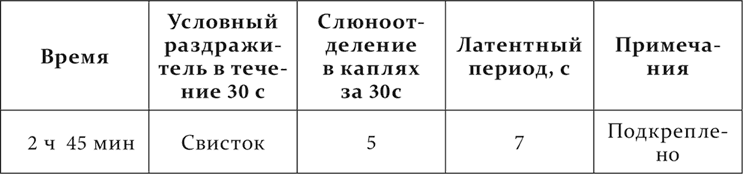
В начале опыта условный раздражитель подкреплялся то 5 секунд спустя, то 30, чтобы сохранить нормальную величину его эффекта до конца опыта.
Мы видим, что при повторениях агента, вызывающего исследовательский рефлекс, его двигательный эффект постепенно все уменьшается, а затем при продолжающихся повторениях то прямо наступают сонливость и сон, все углубляющиеся, то перед этим агент остается некоторое время как бы без всякого действия. Однако присоединяемый к нему условный раздражитель (во втором из только что приведенных опытов) свидетельствует, что он в этот период производит тормозящее действие. Что тормозящее действие на условный раздражитель не основано теперь на механизме так называемого нами внешнего торможения, доказывается тем, что (как это мы имели в шестой лекции при изучении действия исследовательского рефлекса на запаздывающий условный рефлекс) исследовательский рефлекс, когда он оказывается очень малым, растормаживает, а не тормозит условный рефлекс. Следовательно, здесь действует уже торможение, развивающееся при повторении исследовательского рефлекса и переходящее потом в сон, при котором условный рефлекс и совсем исчезает (в первом из только что приведенных опытов).
То же мы видим в особенности резко выраженным на щенках (опыт И. С. Розенталя). При однообразном повторении какого-нибудь раздражителя, при отсутствии вообще колебаний в окружающей среде, щенки часто поразительно точно и быстро впадают в сон. А с другой стороны, разве это не общеизвестная истина, хотя до сих пор остававшаяся без научного освещения, что все люди, а особенно не имеющие сильной внутренней жизни, при однообразных раздражениях, как бы это ни было неуместно и несвоевременно, неодолимо впадают в сонливость и сон. Значит, определенные корковые клетки, реагирующие на данный длительно продолжающийся внешний агент, истрачиваясь, переходят в тормозное состояние, а при отсутствии противодействия со стороны других деятельных пунктов коры тормозной процесс, распространяясь, обусловливает сон. Чрезвычайно быстрая истощаемость корковой клетки с переходом клетки в тормозное состояние резко контрастирует с выносливостью клеток низших отделов головного мозга при тех же условиях. Опыты Зеленого в нашей лаборатории показали, что в то время как у нормальной собаки на определенный звук исследовательский рефлекс быстро исчезает, тот же звук при той же обстановке у собаки без больших полушарий вызывает этот рефлекс стереотипно огромное число раз.
Возвращаюсь к условным рефлексам.
«Корковая клетка под влиянием условных раздражений непременно рано или поздно, а при частых повторениях их и очень быстро, приходит в тормозное состояние»
Что основание для развития торможения, а за ним и для наступления сна дает истощение корковой клетки и вообще ее ослабление, доказывается и следующими постоянно перед нами повторявшимися фактами. Когда мы хирургически, операцией на полушарии, нарушали какой-нибудь анализатор, то положительные условные раздражители, к нему относящиеся, или почти нельзя было применять изолированными даже очень короткое время, – они быстро превращались в тормозные, – или даже они совсем теряли их положительное действие, становясь только отрицательными, тормозящими. Это в особенности легко и постоянно наблюдается при нарушении кожного анализатора.
Когда у собаки удаляются guri coronarius и ectosylvius, то у нее на значительный срок (часто на многие недели) исчезают положительные условные кожно-механические рефлексы с конечностей, с плечевого и тазового поясов туловища, заменяясь отрицательными тормозными. Последнее доказывается тем, что положительные условные раздражители из других анализаторов хорошо действуют до применения кожно-механического раздражения, теряя свое действие после него. Вместе с этим эти кожно-механические раздражения чрезвычайно легко и скоро вызывают сон у таких собак, которые раньше при тех же раздражениях никогда не спали во время нашей работы с ними. Факт часто принимает следующую выразительную форму. Условный кожно-механический раздражитель на поврежденном мозговой операцией отделе кожной поверхности ведет к торможению и сонливости, а тот же раздражитель на неповрежденных местах дает положительный эффект и оставляет животное совершенно бодрым (опыты Н. И. Красногорского, И. П. Разенкова и В. М. Архангельского).
«Торможение тесно связано со сном, прямо переходит в сон, если не принимаются соответствующие меры»
Сюда же справедливо отнести и факт, наблюдавшийся в лаборатории в наш голодный период, несколько лег тому назад. На истощенных животных нельзя было вести исследование с условными рефлексами, так как все положительные условные раздражители чрезвычайно быстро становились у них отрицательными, и вместе с тем собаки начинали спать именно в связи с применением условных раздражителей. Очевидно, общее истощение животного особенно давало себя знать на корковых клетках (опыт Фролова, Розенталя и др.).
В перечисленных до сих пор многочисленных случаях мы постоянно видели, что торможение переходит в сон, но можно наблюдать и обратное – переход сна в торможение. Мы выработали отставленный на 3 минуты условный рефлекс. Отдельный опыт иногда идет следующим образом. Мы ставим животное в станок. Оно бодро. Но как только применяется условный раздражитель, животное сейчас же становится сонным, слюноотделительный эффект отсутствует все 3 минуты, и при подаче еды после этого животное не сразу и вяло ест подаваемую пищу. Мы повторяем в опыте через обыкновенные промежутки условный раздражитель несколько раз. С каждым разом наша собака при раздражении держится все бодрее, в конце 3 минут раздражения появляется слюноотделение. С повторением это слюноотделение все растет. Наконец трехминутный период раздражения делится приблизительно на две половины: в первую нет слюноотделения, хотя животное остается совершенно бодрым, во вторую обильное слюноотделение, и собака стремительно и с жадностью ест подаваемую ей еду. Здесь иррадиированное торможение – сон, – наступившее в силу преобладания сначала тормозного процесса при первой половине действия условного раздражителя, постепенно переходит в ограниченное, концентрированное торможение под давлением все усиливающегося процесса раздражения, связанного со второй половиной действия того же условного раздражителя. В подобных случаях приходится видеть и чистую замену торможения сном. Как при продолжительном отставлении на 3 минуты, так и при более коротком, на 30 секунд, иногда наблюдается следующее отношение. Животное, постоянно остающееся на станке во время опыта бодрым, точно каждый раз с началом действия повторяющегося условного раздражителя засыпает: закрываются глаза, опускается голова, все туловище расслаблено виснет на лямках, временами даже слышится храп. Но проходит определенный срок, при длинном отставлении 1,5–2 минуты, при коротком 25 секунд, животное быстро само собой просыпается, появляется слюноотделение и обнаруживается резкая пищевая двигательная реакция. Ясно, что в данном случае концентрированное торможение хронически заменяется разлитым сном. Наконец, можно иногда видеть, как суммирование двух торможений дает сонливость.
«При разных случаях внутреннего торможения степень вмешательства сна несколько различна. Всего меньше он дает себя знать именно при условном торможении»
У одной собаки (опыт Фурсикова) имелся хорошо выработанный условный рефлекс, отставленный на 3 минуты. Первые 2 минуты не было слюноотделения, затем оно начиналось, достигая максимума к концу третьей минуты. В данном опыте вместе с условным раздражителем применяется посторонний агент – слабое шипение. Оно растормаживает первую половину рефлекса, причем на шипение была заметна небольшая двигательная реакция, исследовательский рефлекс. Условный рефлекс был подкреплен. При повторении той же комбинации ориентировочной реакции на шипение больше нет, а условный рефлекс совершенно исчезает и у животного замечается резкая сонливость. Рефлекс опять подкрепляется. Результат, очевидно, надо понимать так. Исследовательский рефлекс на шипение со второго раза уже гаснет, т. е. теперь на шипение развивается торможение, как об этом сказано раньше в настоящей лекции. Это торможение, складываясь с торможением первой фазы запаздывающего рефлекса, усиливает торможение настолько, что деятельная фаза рефлекса отпадает, уступая место сонливости животного. Что это толкование хода опыта верно, свидетельствует продолжение опыта. При следующем повторении условного раздражения без присоединения шипения получается правильный запаздывающий рефлекс с двумя фазами. Повторенная еще раз комбинация метронома с шипением опять производит сонливость при исчезании условного рефлекса.
Вот подлинные цифры этого опыта.
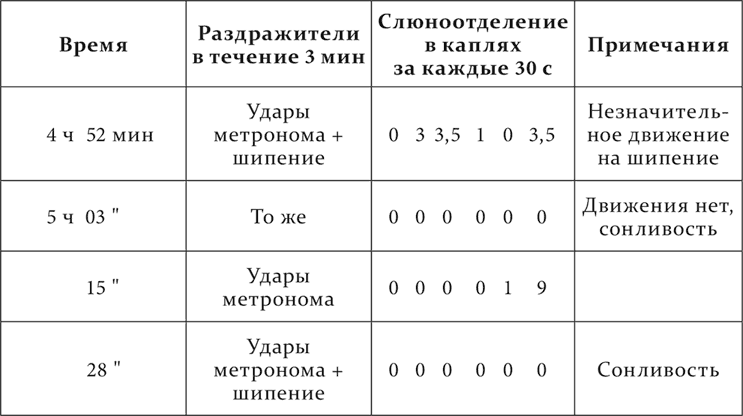
При этом случае считаю небесполезным ради ясности нарочито обратить ваше внимание на следующее. Очевидно, этот опыт вместе с вышеприведенными опытами Чечулина прибавляет лишнюю новую фазу в действии посторонних агентов на условный рефлекс. Если взять сильный внешний агент, то он, как вы помните из шестой лекции о запаздывающем рефлексе, вызываемым им исследовательским рефлексом сперва тормозит весь запаздывающий рефлекс, затем, когда исследовательский рефлекс при повторении значительно слабеет, он только растормаживает первую фазу запаздывающего рефлекса и наконец, как вы узнаете теперь, он еще раз тормозит этот рефлекс, но уже на другом основании, сам становясь первичным возбудителем тормозного процесса в коре полушарий. Слабый же внешний агент, как это оказалось в опыте Фурсикова, обусловливая с самого начала слабый и коротко продолжающийся исследовательский рефлекс, начинает при первом применении прямо с растормаживающего действия на запаздывающий рефлекс и кончает тем же вторым торможением.
«Внутреннее торможение в бодром состоянии есть раздробленный сон, сон отдельных групп клеток, как сон есть внутреннее торможение, иррадиированное, распространившееся сплошь на всю массу полушария и на лежащие ниже отделы головного мозга»
За тождество процессов торможения и сна говорят также и их общие свойства. В прошлых лекциях мы даже слишком много видели фактов, бесспорно доказывающих движение тормозного процесса по массе больших полушарий, причем это движение оказывалось очень медленным, измерялось минутами, и даже многими, и, кроме того, очень варьировало в отношении скорости у разных животных и при разных условиях. Очевидно, что и сон – тоже движущийся процесс. Все мы знаем, как постепенно овладевают нами сонливость и сон и как часто они трудно и медленно отступают, рассеиваются. Имеются и научные сведения о постепенном исчезании при засыпании работы разных органов чувств и других более сложных умственных деятельностей. С другой стороны, общеизвестны очень большие разницы между разными людьми в отношении скорости засыпания и пробуждения, а также разницы этой скорости и при разных условиях. То же самое наблюдалось нами у собак.
Далее в нашей работе мы постоянно видели, как торможение, если оно сначала развивается трудно, от практики, в связи с повторением и применением разных случаев торможения делается все легче и легче воспроизводящимся процессом. Точно так же условные раздражители, когда они при соответствующих условиях обусловливают сон, а также индифферентные раздражители вызывают сон быстрее и быстрее, чем чаще они повторяются.
Интересно особенно следующее. Как показано ранее, торможение индуцирует возбуждение. У некоторых собак, у которых, как сообщено это несколько раньше, при запаздывающем рефлексе условный раздражитель вместо торможения в начальной фазе раздражения заменяется сном, наступлению сна иногда предшествует на очень короткое время небольшое общее возбуждение животного. А еще рельефнее и постояннее это явление выступает при засыпании животного под влиянием продолжительного или повторяющегося индифферентного раздражителя. Это часто наблюдалось в вышеупомянутых опытах Розенталя. Когда индифферентный раздражитель явно начинает делать щенка сонливым, то, перед тем как ему окончательно заснуть, он приходит на некоторое время в возбужденное состояние: начинает беспокойно двигаться, чесаться, лаять беспричинно в воздух. То же мне приходилось замечать и на засыпающих детях. Совершенно своеобразная и неожиданная картина. Можно с правом видеть в ней явление индукции. Так же могло быть понимаемо и известное возбуждение при начале наркотизации.
«Всего легче развивается торможение при температурных раздражениях и всего труднее при звуковых. Строго параллельно с этим сон быстро наступает при температурных условных рефлексах и позже и реже при звуковых»
Мне кажется, что совокупность приведенных фактов должна считаться достаточной для полного доказательства истинности нашего положения, что сон и внутреннее торможение – один и тот же процесс. Я лично сейчас не знаю, не вижу ни одного факта в нашей работе, который бы противоречил серьезно нашему заключению. Надо сожалеть, что у нас до сих пор нет хорошего, графического способа для изображения сна. Только изредка мы пользовались для этой цели записыванием положения головы животного. Конечно, сообщение наших опытов, относящихся ко сну, сопровождаемое тем или другим изображением сна, очень бы прибавило в ваших глазах к убедительности наших доводов.
Очевидно, с нашим заключением вполне гармонируют и все подробности нашего нормального обычного сна. Наша дневная работа, у одних очень однообразная, у других, наоборот, очень разнообразная, одинаково в конце должна обусловливать наступление сна. Продолжительное раздражение одних и тех же пунктов коры ведет к очень глубокому торможению их, которое, естественно, сильно иррадизируя, захватывает полушария и опускается на нижележащие отделы головного мозга. С другой стороны, при разнообразной деятельности, хотя отдельные пункты коры не доходят до значительной степени тормозного состояния, зато их большое количество создает и без большой иррадиации распространенное тормозное состояние, также спускающееся ниже. Конечно, очень большое количество быстро сменяющихся раздражений может часто долго сопротивляться общему захватыванию полушарий тормозным процессом и отсрочивать наступление сна. И обратно, строго заведенный порядок в смене бодрствования и сна, установленный ритм может увеличивать настойчивость сна и без достаточного утомления клеток коры. Для того и другого мы имели в наших опытах достаточно примеров аналогичных отношений раздражительного и тормозного процессов.
«В высшей степени важно иметь полный список и надлежащую систематизацию рефлексов, потому что вся остальная нервная деятельность организма надстраивается на фундаменте этих рефлексов»
«С фактом условного рефлекса отдается в руки физиолога огромная часть высшей нервной деятельности, а может быть и вся»
«Большие полушария являются главнейшим органом условных рефлексов, они обладают синтетической функцией в таком объеме и совершенстве, как никакая другая часть центральной нервной системы»
Назад: Лекция четырнадцатая
Дальше: Лекция шестнадцатая

