Книга: Лекции о работе больших полушарий головного мозга
Назад: Лекция двенадцатая
Дальше: Лекция четырнадцатая
Лекция тринадцатая
Кора как мозаика: а) примеры мозаичности коры и наиболее простые способы ее происхождения; б) изменяемость физиологической роли отдельных пунктов; в) стойкость некоторых пунктов. – Кора как объединенная сложная динамическая система
Мм. гг.! Как очевидно из всего сообщенного доселе, бесконечная масса явлений природы постоянно обусловливает посредством аппарата больших полушарий образование то положительных, то отрицательных условных рефлексов и тем подробно определяет всю деятельность животного, его ежедневное поведение. Для каждого из этих рефлексов в коре полушарий должна иметься своя точка приложения, т. е. своя клетка или группа клеток. Одна такая единица коры связана с одной деятельностью организма, другая – с другой, одна вызывает деятельность, другая ее не допускает, подавляет. Таким образом, кора полушарий должна представлять собой грандиозную мозаику, грандиозную сигнализационную доску. При этом на этой доске постоянно остаются огромные запасы пунктов для образования новых сигнальных условных раздражителей, а кроме того, занятые ранее пункты более или менее часто подвергаются изменениям в их связях с разными деятельностями организма, в их физиологическом значении.
Механизм такой функциональной мозаичности коры полушарий как с анатомической стороны, так и с физиологической отчасти намечается и сейчас. Как состав рецепторов из множества элементов, так и обилие клеток коры с их огромным разнообразием в величине, форме и расположении в значительной степени отвечают требованию этой мозаичности. С этим достаточно согласуется, хотя и более грубая, локализация, устанавливаемая современной физиологией коры. Но в целом вопрос о функциональных разграничениях коры, конечно, вопрос чрезвычайной, исключительной сложности. В настоящее время могут быть делаемы только самые первые шаги экспериментального подхода к изучению его наипростейших сторон. Однако и тут предмет все еще сложен и представляет большие трудности для исследования. Пусть функциональное размежевание коры по крайней мере в некоторых случаях допускалось бы на чисто анатомических основаниях, т. е. благодаря существованию отдельных клеток, отвечающих отдельным рецепторным элементам. Но мы знаем из факта первоначальной генерализации условных раздражителей, что раздражение сперва непременно иррадиирует из исходного пункта, захватывает клетки и других рецепторных элементов. Каким же образом оно опять сосредоточивается в исходном пункте? Мы должны различать тут два случая. Как размежевываются друг от друга два положительных пункта, когда они связываются с двумя разными безусловными рефлексами, и как разграничиваются положительный и отрицательный (т. е. тормозно действующий) пункты, когда они относятся к одному и тому же безусловному рефлексу? Наш наличный материал почти исключительно до последнего времени имел дело только со вторым случаем, как с более простым. За решение первого вопроса мы взялись только теперь, и потому мое дальнейшее изложение будет касаться только второго вопроса. Здесь я соединю наши данные, относящиеся к теме о мозаике и не вошедшие в предшествующие лекции. Начнем с наиболее очевидного случая.
«Бесконечная масса явлений природы постоянно обусловливает посредством аппарата больших полушарий образование то положительных, то отрицательных условных рефлексов и тем подробно определяет всю деятельность животного, его ежедневное поведение»
Вот собака, у которой кроме других условных пищевых рефлексов имеются кожно-механические: положительный на правом плече и отрицательный на правом бедре. Когда эти рефлексы были хорошо выработаны, впервые испытывались на их эффект новые места на передней лапе на 17 см вниз от места на плече, на боку на 12 см кзади от места на плече, тоже на боку на 15 см кпереди от места на бедре и, наконец, на задней лапе на 18 см вниз от места на бедре. Слюноотделение отмечалось за 30 секунд изолированного действия условного раздражения.
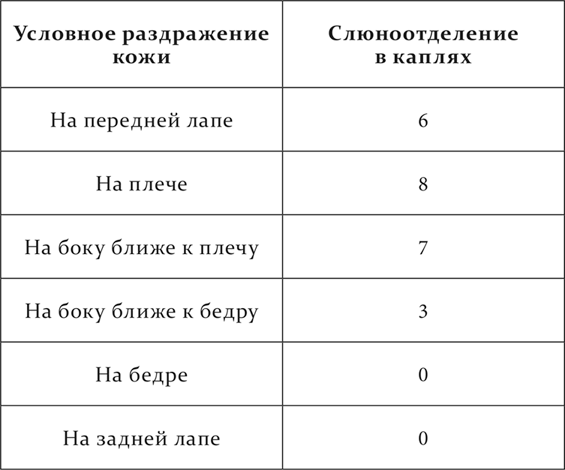
То же получилось и еще на двух собаках, служивших для этих опытов (опыты Фурсикова).
Мы видим, что под влиянием внешних раздражений, действовавших при разных условиях (одно сопровождалось безусловным рефлексом, другое – нет), в каждом анализаторе возникли два разных пространственно разделенных пункта, пункт раздражения и пункт торможения, причем около каждого из них образовался соответственный район. Эти районы, распространяясь навстречу друг другу, пространственно разграничивались. Резкое уменьшение положительного эффекта на боку ближе к бедру и нуль эффекта на задней ноге надо рассматривать как преобладание тормозного процесса, потому что генерализация условного положительного кожно-механического раздражения, как это мы видели в десятой лекции, обыкновенно захватывает весь анализатор, и уменьшение эффекта по мере удаления от места выработанного рефлекса происходит очень постепенно, а не обрывисто.
«Кора полушарий должна представлять собой грандиозную мозаику, грандиозную сигнализационную доску. При этом на этой доске постоянно остаются огромные запасы пунктов для образования новых сигнальных условных раздражителей, а кроме того, занятые ранее пункты более или менее часто подвергаются изменениям в их связях с разными деятельностями организма, в их физиологическом значении»
Таким образом, первое основание для функциональной мозаики коры дают внешние раздражения, производящие противоположные процессы, как это мы уже видели в лекции об анализаторной деятельности коры.
Затем на следующих трех собаках мы поставили более сложные опыты. У двух собак на разных тонах (опыты В. В. Сирятского), а у одной на разных местах кожи (опыты Купалова) мы правильно чередовали несколько положительных и отрицательных пунктов. Это предпринималось с несколькими целями, чтобы видеть: как вырабатывается мозаика, как далеко она может простираться, какой достигает точности, как взаимно действуют ее разные пункты, как относятся промежуточные пункты и как эта мозаика отзывается на общем состоянии животного?
Некоторые из результатов этих опытов я приведу сейчас, другие будут использованы в других отделах моего изложения. Вообще же исследование этих животных еще не закончено.
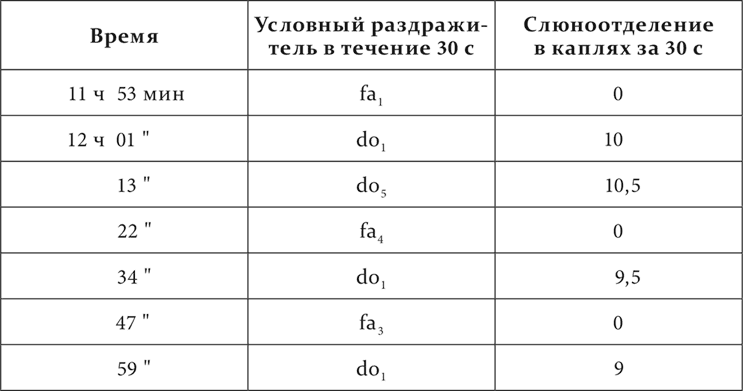
У одной собаки из пяти смежных октав фисгармонии от большой октавы до третьей включительно (от 64 колебаний в секунду до 1440) все тоны do были сделаны положительными условными возбудителями пищевой реакции, все тоны fa – отрицательными. В протоколах тоны do и fa отдельно обозначены номерами по порядку, начиная с низких.
Вот окончательное положение дела у этой собаки.

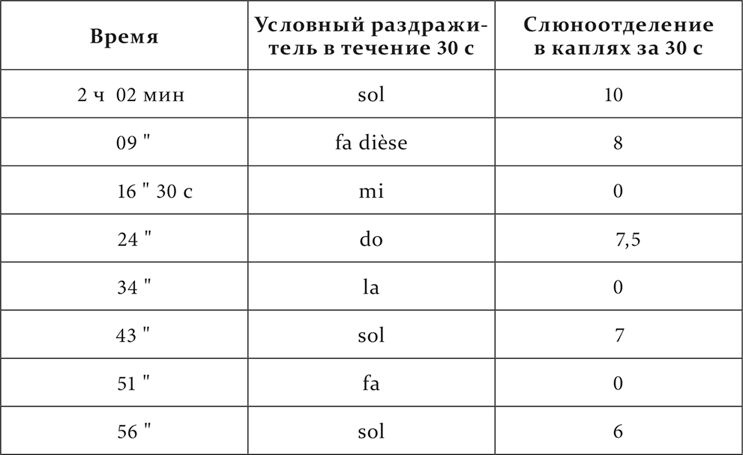
У другой собаки в пределах одной октавы от fa малой октавы до fa dièse первой октавы тоны тоже сделаны чередующимися положительными и отрицательными условными раздражителями, но без правильного ритма, что касается промежутков между действующими тонами и на более коротких расстояниях между ними, именно: fa dièse – положительный, mi – отрицательный, do – положительный, la – отрицательный, sol – положительный и fa – отрицательный.
Вот окончательное положение дела у этой собаки.
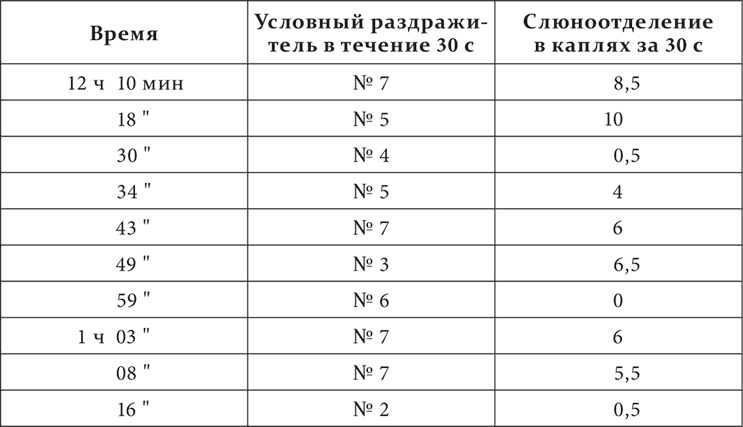
У третьей собаки были выработаны условные кожно-механические пищевые рефлексы. Приборы для раздражения были расположены на строго определенных местах по линии от левой голени через все туловище до левого предплечья, на одинаковых расстояниях один от другого, именно на расстоянии 12 см, считая от центра приборов. Счет приборов велся от голени к предплечью. Всех приборов было 9. Раздражения нечетными приборами были сделаны положительными, раздражения четными – отрицательными.
Вот один из более резких опытов.
Представленная функциональная мозаичность коры давалась ей трудно, но только на первых порах выработки. Но чем дело шло дальше, тем выработка становилась все более легкой. Особенный интерес заключается в том, что в конце выработки у первой и третьей собак некоторые новые рефлексы были готовы сами собой сразу. Нужно иметь в виду, что новые условные раздражения в этих случаях возникали в районах противоположного нервного процесса.
«В целом вопрос о функциональных разграничениях коры, конечно, вопрос чрезвычайной, исключительной сложности»
У первой собаки был, и уже давно, выработан положительный условный рефлекс на do4. В данном опыте это do4 имело эффект в 8 капель за 30 секунд. Когда впервые было применено раздражение fa4, то оно осталось без положительного эффекта, т. е., надо было думать, было прямо тормозным. Это доказал опыт следующего дня. Этот день был начат с пробы fa4, которое и теперь оказалось без положительного эффекта, а последовавшее за ним положительное раздражение do4 являлось в значительной степени заторможенным. Следовательно, fa4 действительно сделалось тормозным само собой без выработки, находясь в районе положительного do4. То же самое повторилось с do5, которое, испробованное в первый раз, дало положительный эффект, находясь в районе тормозного fa4.
У третьей собаки с кожно-механическими условными раздражениями оба крайние положительные раздражения № 1 и № 9 также были налицо прямо без выработки, притом с полным эффектом, находясь оба в районах тормозных пунктов № 2 и № 8.
Естественное объяснение последних фактов надо искать в явлениях взаимной индукции, исходящей из пунктов фиксированного раздражения и торможения, пунктов, расположенных притом ранее в правильном чередовании. Отсюда становится также понятным, почему ритмическая деятельность вообще особенно легка и выгодна. На всех только что описанных собаках постоянно повторялся факт, что при правильном чередовании положительных и отрицательных раздражений рефлексы отличались особенной точностью. Вот пример, как это чередование постепенно даже в течение одного опыта все резче отграничивает положительные и отрицательные пункты, если их эффекты вначале в значительной степени сливались.
Третья собака с кожно-механическими раздражениями.
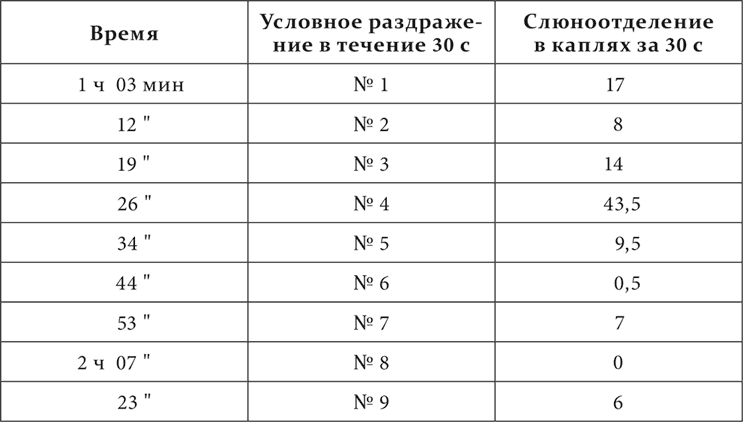
Вот еще примеры, взятые из работы над первой собакой, когда рефлексы вообще еще не были регулярны.
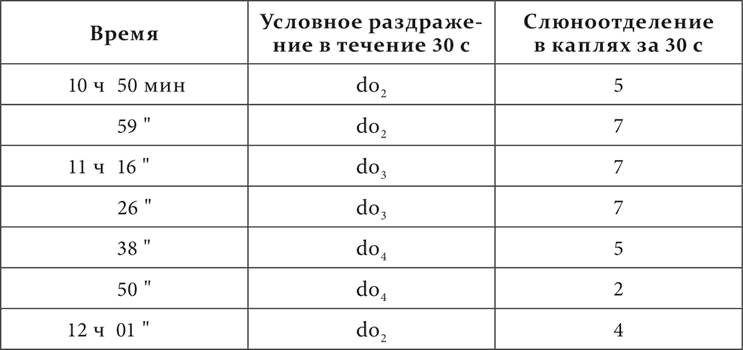
Та же собака спустя 3 дня.


Таким образом, дальнейшее основание для функциональной мозаичности коры надо видеть во взаимной индукции нервных процессов. Затем в этих же опытах мы занимались определением районов (полей) как раздражительного, так и тормозного процессов, а также возможных индифферентных, промежуточных между деятельными районами.
«Мы знаем из факта первоначальной генерализации условных раздражителей, что раздражение сперва непременно иррадиирует из исходного пункта, захватывает клетки и других рецепторных элементов»
На собаках с тоновой мозаикой мы пробовали другие тоны, находящиеся между нашими положительными и отрицательными раздражителями, и наблюдали их эффекты. Положительные эффекты в виде наступающего слюноотделения, конечно, были очевидны прямо. Что касается до нулевых, то надо было разобраться особыми вариациями опыта в их индифферентном или тормозном значении. Мы испытали их ради этого или на их последовательное торможение, или на их положительную индукцию.
Опыт 9.X.1925

Опыт 14.X.1925
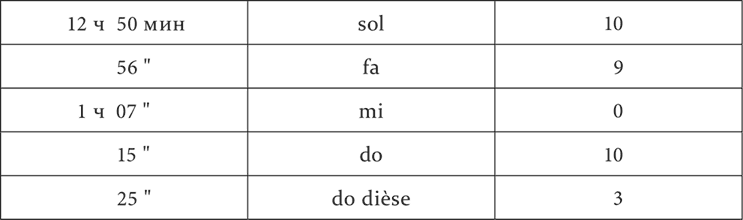
Опыт 20.X.1925
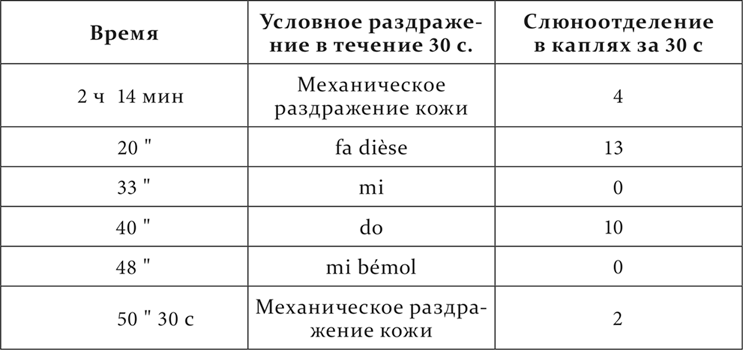
Мы видим, что из расположенных между положительным do и отрицательным mi промежуточные тон re и полутоны do dièse и mi bémol оказались разного значения. Полутон do dièse имел, хотя и уменьшенный, но положительный эффект, т. е. принадлежал к району положительного do. Тон re и полутон mi bémol, по-видимому, были одинаковы по действию, дав нуль. Но испытание их на последовательное торможение обнаружило их разницу. 2 минуты спустя после применения полутона mi bémol условное механическое раздражение потеряло в своем обыкновенном эффекте 50 %, т. е. было заторможено, после же тона re в тех же условиях времени осталось без изменения. Следовательно, полутон mi bémol бесспорно находился в районе отрицательного mi, тон же re или был совершенно индифферентным, или заторможен в гораздо меньшей степени. Последнее надо было решить более тонкими опытами, и потому вопрос об истинно индифферентных пунктах подвергается дальнейшему исследованию.
В этих же опытах исследовалось также взаимодействие пунктов при непосредственном переходе с раздражением от одних положительных пунктов к другим положительным, от одних отрицательных к другим отрицательным, от положительных к отрицательным, и наоборот. Но результаты большей частью получались такой сложности, что мы были вынуждены решать ту же задачу в более упрощенной обстановке.
Как указано было еще в первой лекции и упомянуто опять в начале этой, функциональная мозаика коры не только постоянно может пополняться, но и подлежит частой переделке, т. е. одни и те же пункты коры связываются то с одной, то с другой физиологической деятельностью организма. К последнему пункту относится у нас только одно исследование (опыты С. С. Фридемана).
Одни и те же агенты делались сначала условными пищевыми раздражителями, а затем переделывались в кислотные, и наоборот – прежние кислотные раздражители превращались в пищевые. У двух собак первоначальный условный пищевой раздражитель был превращен в условный кислотный, а у третьей – кислотный в пищевой. Производилось это, конечно, так, что условный раздражитель сопровождался теперь другим безусловным раздражителем. Общий ход дела представляется в следующем виде. Условный раздражитель быстро, даже в течение одного опыта, теряет свой прежний слюноотделительный эффект и становится нулевым на более или менее значительный срок и затем снова приобретает слюногонное действие – и это уже был другой рефлекс, как показывало резкое изменение в составе слюны подчелюстной железы. Вся эта процедура требует около 30 сочетаний условного раздражителя с новым безусловным раздражителем. Когда затем у первых двух собак после довольно длинного периода с условными кислотными рефлексами условные раздражители еще раз подвергались превращению опять в пищевые, это превращение произошло очень быстро, через небольшое число раз. Значит, первая связь до известной степени уцелела, несмотря на существование второй связи.
«Первое основание для функциональной мозаики коры дают внешние раздражения, производящие противоположные процессы»
В настоящем исследовании главный интерес для нас привязывался, однако, к другому пункту. Мы желали знать, сохранится ли дифференцировка, выработанная для условного раздражителя на одном рефлексе, при превращении его в условный раздражитель другого рефлекса. У всех трех собак результат получился тождественный. Я опишу его более подробно для одной из собак. Условным пищевым раздражителем был тон mi (2600 колебаний в секунду). От него была выработана и хорошо концентрирована дифференцировка на соседний низкий тон (2324 колебания в секунду). Затем условный пищевой раздражитель был превращен в условный кислотный. Та же дифференцировка, испробованная в первый раз на кислотном рефлексе, оказалась полной и в такой же степени концентрированной. Теперь была выработана новая более тонкая дифференцировка на полутон выше (2760 колебаний в секунду). Когда тон mi снова был сделан пищевым условным раздражителем, то и эта новая дифференцировка также вполне сохранилась.
На протяжении предыдущих лекций мы достаточно познакомились с фактом, что одни и те же пункты коры, смотря по условиям, являются местом то раздражительного процесса, то тормозного, вызывают то одну работу организма, то другую. И это изменение их физиологической роли происходит довольно легко, т. е. более или менее скоро они меняют свое состояние (прошу припомнить опыты с отрицательной индукцией) и свои связи (как показывают только что сообщенные опыты).
Но мы имеем также и факты, когда определенная роль известных пунктов и отделов коры или даже временные отношения процессов в них делаются чрезвычайно упорными, трудно или совсем не поддающимися переменам. С этим мы встретились в работе о следовых положительных и отрицательных условных рефлексах (опыты Фролова). Тут некоторые явления носили такой исключительный характер, что я считаю необходимым описать их подробно.
У одной собаки был выработан условный следовой кислотный рефлекс на тон органной трубы в 1740 колебаний в секунду, причем тон продолжался 15 секунд, и по прекращении его, спустя 30 секунд, производилось вливание собаке в рот кислоты. Этот следовой рефлекс повторился 994 раза в течение 1 года и 9 месяцев. Латентный период, считая от начала тона, составлял около 25 секунд, или, считая иначе, 10 секунд от начала следов тона. Когда одни опыты с этим рефлексом были покончены, было решено тот же тон переделать для других опытов из следового условного раздражителя в наличный условный, т. е. получать от него эффект не по прекращении, а еще во время его действия.
Для этого тон на пятнадцатой секунде его продолжения сопровождался вливанием кислоты. После двадцатикратного повторения такой процедуры не только не появилось условного рефлекса в течение 15 секунд, но после вливания кислоты слюноотделение начиналось лишь спустя 10 секунд, т. е. спустя те же 25 секунд от начала тона. Примененные вместо тона другие звуковые раздражители ничего не изменили в положении дела. Когда же тон и эти раздражители изолированно без вливания кислоты применялись 45 секунд, то спустя около 25 секунд от начала их слюноотделение начиналось и быстро достигало значительной величины (10 капель за остальные 20 секунд). После этого вливание кислоты начали производить спустя только 2 секунды от начала звучания тона. И, однако, после очень большого количества (многих десятков раз) таких сочетаний слюноотделение как во время вливания кислоты, так и при одном звучании тона без кислоты начиналось только около двадцатой – двадцать пятой секунды от начала тона.
Вот последний относящийся сюда опыт.
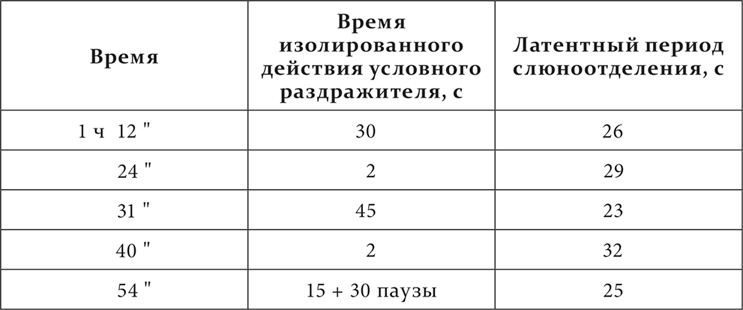
В опыте условное раздражение всякий раз подкреплялось кислотой.
Наконец на тон fis тонвариатора был выработан пищевой рефлекс, но и на нем латентный период упорно держался на 24–28 секундах.
И лишь когда условным кислотным раздражением было взято кожно-механическое раздражение, подкрепляемое через 2 секунды от его начала, на двадцать четвертой пробе получилось условное слюноотделение за 30 секунд в 12 капель с латентным периодом в 2 секунды.
То же упорство выработанного тормозного процесса наблюдалось и у другой собаки. У нее из следов тона органной трубы в 1740 колебаний в секунду был сделан условный тормоз к ударам метронома (104 удара в минуту) как положительному условному раздражителю. Даже следы тона спустя 60 секунд по прекращении его были действительным условным тормозом. Такое торможение, конечно, должно было вырабатываться очень постепенно, начиная с более ранних следов, и очень медленно. А вся выработка и работа с таким условным торможением заняла более двух лет. Нужно еще прибавить, что к этому условному тормозу были присоединены и его дифференцировки. Следовательно, торможение было, так оказать, очень напряженное. Затем собака должна была служить для другой темы, ради которой было приступлено к выработке положительного условного раздражителя из звука телефонной пластинки, приводимой в колебание переменным током частоты, почти совпадающей с числом колебаний тона. Положительный условный рефлекс необычно долго не проявлялся. Тогда новый агент был применен перед ударами метронома – и тогда обнаружилось его резкое условное тормозящее действие. Имея пример стойкости торможения на первой собаке, здесь мы дальнейшую процедуру варьировали таким образом, что стали сопровождать безусловным раздражителем тормозную комбинацию звука телефонной пластинки с ударами метронома, т. е. действовали точно обратно тому, как ранее поступали при образовании тормозной комбинации из следов тона с ударами метронома. И, действительно, теперь довольно скоро тормозная комбинация стала положительным раздражителем, а еще несколько времени спустя и звук мембраны также получил положительное условное действие. Однако сам тон органной трубы не сделался от этого положительным раздражителем, и его пришлось лишать тормозного действия особо и так же, как было поступлено с звуком телефонной пластинки. Но на этот раз цель была достигнута не так скоро.
«Функциональная мозаика коры не только постоянно может пополняться, но и подлежит частой переделке, т. е. одни и те же пункты коры связываются то с одной, то с другой физиологической деятельностью организма»
Описанные случаи стойкости тормозного процесса должны считаться чрезвычайными. Вероятно, помимо напряженности процесса здесь существовали и еще особые основания для нее в условиях данных опытов. Может быть, к объяснению их послужат и наши опыты, относящиеся к гипнотизму и сну, о которых речь впереди.
Рядом с этими случаями стойкости тормозного процесса в отдельных пунктах коры можно поставить случай стойкости в отдельных пунктах и процесса раздражения. У собаки (опыты Б. Н. Бирмана) был выработан условный пищевой рефлекс на тон фисгармонии в 256 колебаний в секунду. От него были отдифференцированы 22 тона вверх до тона 768 колебаний в секунду и вниз до тона 85 колебаний в секунду, т. е., как обыкновенно, тон 256 колебаний в секунду сопровождался едой, а все остальные нет. Положительный тон применялся в каждом опыте по нескольку раз, а из тормозных то те, то другие. Таким образом, положительный пункт был как бы окружен тормозным, но это повело не к уменьшению его эффекта, а наоборот – к резкому увеличению. Затем у собаки был произведен глубокий физиологический сон. Среди этого сна, когда резкий свисток или сильный стук кулаком в дверь экспериментальной комнаты, где стояла собака, не будили собаку, положительный тон сейчас же делал собаку бодрой, причем получался полный условный слюноотделительный эффект. Так как в последующих лекциях будет доказано нашими опытами, что сон есть торможение, то в описанном опыте мы имеем пример стойкости в отдельных пунктах коры процесса раздражения при сильном натиске торможения.
Если, с одной точки зрения, кору больших полушарий можно рассматривать как мозаику, состоящую из бесчисленной массы отдельных пунктов с определенной физиологической ролью в данный момент, то с другой – мы имеем в ней сложнейшую динамическую систему, постоянно стремящуюся к объединению (интеграции) и к стереотипности объединенной деятельности. Всякое новое местное воздействие на эту систему дает себя знать более или менее во всей системе. Мы выработали известное количество условных рефлексов. Прибавление новых положительных и особенно тормозных большей частью сейчас же отзывается на состоянии прежних (опыты Анрепа и других). Мало этого. Если рефлексы остаются те же и мы меняем только раз установленный и долго поддерживаемый порядок их, размер эффекта условных рефлексов резко меняется в сторону уменьшения его, т. е. в сторону преобладания тормозного процесса.
Вот пример этого, взятый из работы Д. И. Соловейчика. У собаки через каждые 10 минут применяются следующие условные пищевые раздражители: удары метронома, вспыхивание электрической лампочки, свисток и механическое раздражение кожи в приведенном порядке. В каждом опыте этот ряд повторяется.
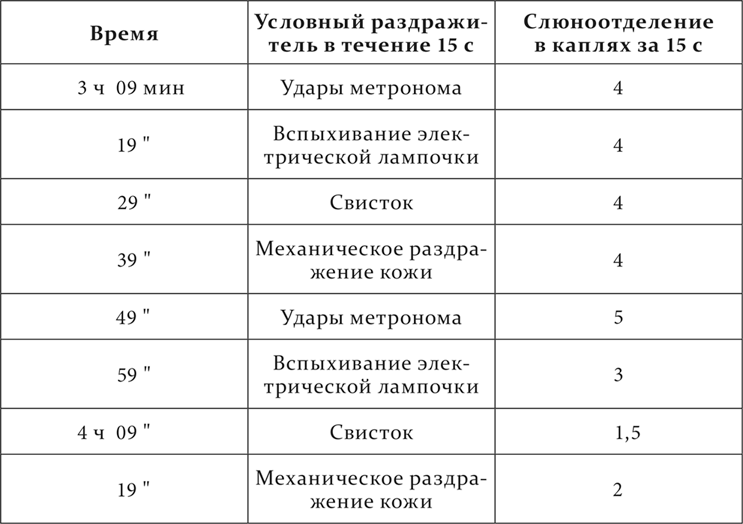
В опыте на другой день порядок тех же раздражителей несколько меняется.

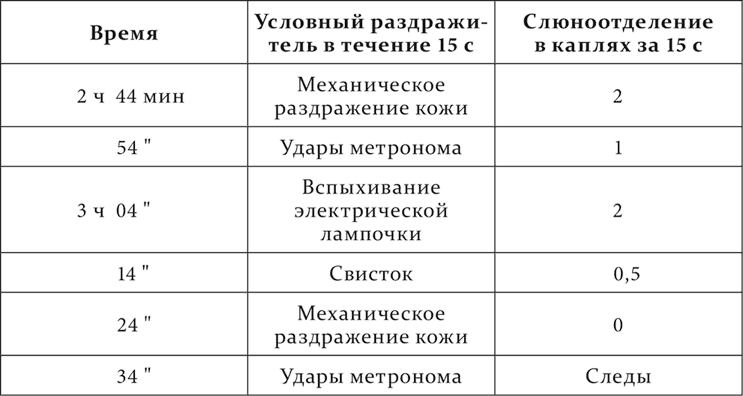
Это уменьшение величины условных рефлексов при изменении их порядка иногда, как в приведенном опыте, наступает сейчас же, иногда же оно резче обнаруживается на другой день уже при восстановлении старого порядка и продолжается то много дней, то быстро сменяется нормальной величиной.
Уменьшение рефлексов при повторном ряде в первом опыте – довольно частое явление, о котором речь будет в следующей лекции.
Назад: Лекция двенадцатая
Дальше: Лекция четырнадцатая

