Книга: Лекции о работе больших полушарий головного мозга
Назад: Лекция одиннадцатая
Дальше: Лекция тринадцатая
Лекция двенадцатая
Комбинирование явлений движения нервных процессов… с… явлениями их взаимной индукции
Мм. гг.! В последних трех лекциях мы познакомились сперва с явлениями иррадиирования и концентрирования нервных процессов в коре больших полушарий, а потом с явлениями взаимной индукции этих процессов. Эти явления были описаны нами отдельно, как бы вне зависимости друг от друга. В действительности же, как это ясно и по смыслу дела, они должны встречаться вместе, комбинироваться, взаимодействовать. Воспроизведение этих явлений в нашем прежнем изложении разрозненными вернее отнести к более редким случаям в работе больших полушарий, приуроченным или к определенным фазам развития и состояния нервных процессов, или к особенным типам нервной системы у наших экспериментальных животных. Иногда же, вероятно, мы сами искусственно упрощали явления иррадиирования и концентрирования процессов, не имея сначала идеи об их взаимной индукции, как это уже упомянуто в начале предыдущей лекции. В начале нашей работы, когда перед нами стояла подавляющая хаотическая сложность явлений, которые мы, однако, решились изучить, приходилось умышленно закрывать глаза на многое, даже прямо устраняться от особенно трудных случаев, заменяя, например, одних собак другими. Теперь уже не то. Многолетний опыт принес свои дары, обнаружил свою силу. Теперь всякая особенность, неожиданность, представляемая тем или другим животным, привлекает наше специальное внимание, делается для нас интересным вопросом, новой задачей.
Комбинирование явлений иррадиирования и концентрирования нервных процессов с явлениями их взаимной индукции представляет собой очень сложный процесс, полного знания которого придется еще долго ждать. Сейчас на эту тему мы только накопляем большей частью отрывочный, почти совершенно не систематизированный материал, к передаче которого я и подхожу теперь. Начну с наиболее простого случая (опыты Е. М. Крепса).
У собаки условным раздражителем служит механическое раздражение кожи. Раздражения мест кожи – двух на бедре (первый и второй пункты), по одному на животе (третий пункт), на груди (четвертый) и на плече (пятый) – были положительными условными (безусловный рефлекс – пищевой); раздражение на голени – тормозное, дифференцировка. Дифференцировка полная, положительные раздражения выравнены в отношении величины эффекта. В начале каждого опыта устанавливается размер слюноотделительной реакции для положительного раздражения. Затем применяется тормозное раздражение. По прекращении его или непосредственно, или через разные промежутки времени испытываются разные положительные раздражения. Такие опыты продолжались в течение 5 месяцев. Результаты их представлены в виде двух таблиц, на которых цифры обозначают величину эффекта положительных раздражений в процентах по отношению к величине эффекта этих раздражений до применения в опыте дифференцировки.
Таблица 1
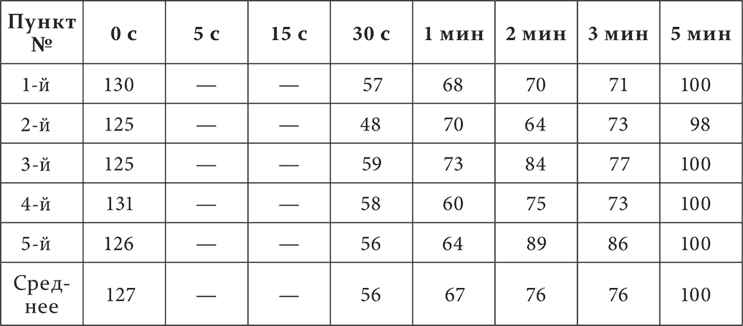
Таблица 2
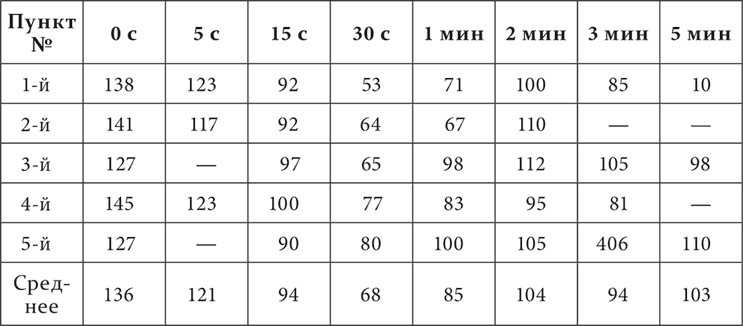
Первый вертикальный столбец указывает номера мест положительных раздражений, верхний горизонтальный – промежутки времени между концом тормозного и началом положительных раздражений. Первая таблица представляет средние цифры из всех опытов за все время работы, вторая – средние цифры только за последний месяц работы.
Отсутствие в таблице 1 цифр за пяти– и пятнадцатисекундный промежутки имеет свое основание в том, что эти промежутки применялись в опытах только за последний месяц работы.
Основной результат опытов заключается, как вы видите, в том, что непосредственно по прекращении тормозного раздражения эффект положительных раздражений оказывается увеличенным, к 15 секундам он уже опускается ниже нормы, а к 30 секундам достигает максимума падения и, наконец, только к пятой минуте снова представляет нормальную величину. Таким образом, картина последействия тормозного раздражения начинается с положительной индукции, а затем только наступает иррадиирование торможения, кончающееся восстановлением нормы эффекта положительного раздражения. Ту же смену положительной индукции последовательным торможением вы могли заметить и в протоколах опытов, приведенных в предшествующей лекции и констатировавших факт положительной индукции. Далее в изучаемых сейчас опытах обращают на себя внимание еще следующие подробности. Размер положительной индукции несколько возрос к позднему сроку работы. Наоборот, последовательное торможение уменьшалось с течением опытов и во времени, заканчиваясь к 2 минутам, и в пространстве, ограничиваясь двумя положительными, ближайшими к тормозному, пунктами. Последнее есть повторение факта, так часто встречавшегося нам в ранних лекциях при изучении разных случаев внутреннего торможения.
«Всякая особенность, неожиданность, представляемая тем или другим животным, привлекает наше специальное внимание, делается для нас интересным вопросом, новой задачей»
Другой также довольно простой случай представился нам на угасательном торможении (опыты Подкопаева).
В лекции об иррадиировании торможения было упомянуто наблюдение Когана, что у одной из его собак непосредственно после полного угашения кожно-механического условного раздражения на одном пункте, на самом дальнем от этого пункта постоянно замечалось повышение возбудимости – положительная индукция. Теперь этот факт в более подробном виде повторился на собаке, у которой был прослежен результат однократного угашения (неподкрепления) условного кожно-механического пищевого рефлекса. У нее были выработаны условные пищевые рефлексы на механическое раздражение нескольких мест (8) кожи одной половины тела, расположенных по линии от нижней части передней конечности через все туловище до нижней части задней конечности. Все места в отношении величины эффекта раздражения были выравнены. Четыре места из них находились между собой на расстояниях 1, 43 и 89 см. На каждом из них и производилось угашение условного рефлекса. Опыт начинался с пробы того или другого из этих мест ради установки нормы рефлекса. Затем на каком-либо из них угашали рефлекс, и теперь через промежутки времени в 45 секунд, 1, 3, 4 и 8 минут снова испытывали место, с раздражения которого начинался опыт. Опыт кончался раздражением безразлично какого-либо из всех 8 мест для проверки еще раз нормы положительного эффекта данного дня. Опыты с угашением ставились через 4–5 дней в расчете иметь более или менее стационарное положение торможения. Но этот расчет не оправдался. Торможение тем не менее все более и более концентрировалось в течение опытов, и таким образом общий результат надо было представить в трех последовательных сериях, на трех таблицах. Собака, на которой делались эти опыты, отличалась таким постоянством величины положительного условного эффекта, что можно было пользоваться каждым отдельным опытом, а не средним из нескольких опытов. В таблицах приведены величины эффекта в процентах по отношению к норме каждого дня. В таблице 5 прибавлены промежутки в 45 секунд, 1 и 3 минуты, потому что только с повторением опытов слюноотделение настолько быстро стало прекращаться, что получилась возможность все раньше испытывать последействие угашения.
Таблица 3
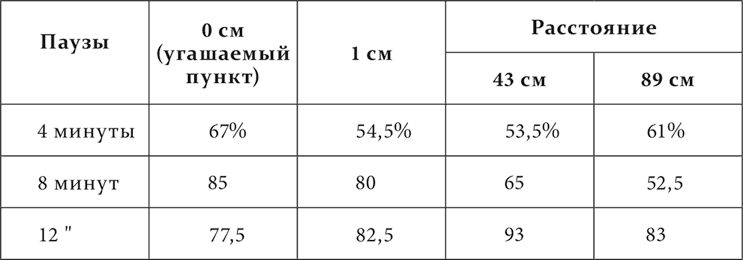
Таблица 4
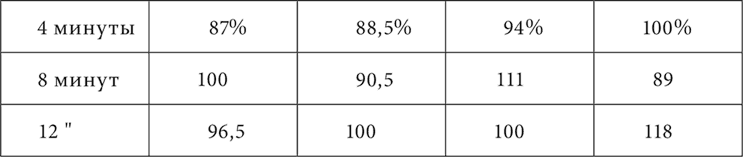
Таблица 5
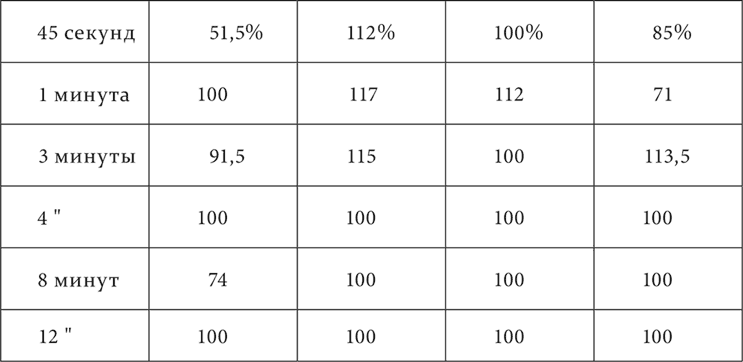
В этих таблицах мы видим, что в начале опытов (таблица 3) торможение простиралось до самого дальнего пункта и еще было отчетливо спустя 12 минут после угашения. Затем (таблица 4) торможение ограничивалось уже расстоянием в 43 см и было через 4 минуты меньше того, что было в предшествующей фазе опытов через 12 минут. Теперь на дальнем пункте и в более позднее время начинает встречаться, хотя и редко, вместо уменьшения увеличение эффекта сравнительно с нормальной величиной. Наконец, в последней фазе (таблица 5) торможение с колебаниями во времени держится только в первично угашаемом месте. На остальных пунктах, за исключением дальнего, в течение минуты даже на 1 см расстояния и к концу минуты после угашения или преобладает увеличение эффекта против норм, или устанавливается норма.
Комбинирование явлений иррадиирования и концентрирования нервных процессов с явлениями их взаимной индукции представляет собой очень сложный процесс, полного знания которого придется еще долго ждать»
Перед нами – положительная индукция, сперва возникающая поодаль от исходного пункта торможения и обнаруживающаяся более поздно, а затем постепенно подвигающаяся по направлению к исходному пункту и все быстрее появляющаяся после начала торможения, т. е. как бы постепенно преодолевающая и вытесняющая и во времени и в пространстве тормозной процесс.
Только что приведенные опыты интересны и своими подробностями. Во-первых, обращает на себя внимание высочайшая чувствительность элементов коры, когда однократное неподкрепление сказывается на большой территории коры и держится значительное время (более 12 минут). Во-вторых, перед нами лишнее свидетельство текучести, изменяемости явлений в работе больших полушарий, когда незначительное влияние, повторяемое через длинные сроки в 4–5 дней, резко меняет общее положение дела (быстрая смена фаз). Наконец, нельзя не заметить, что состояние разных пунктов полушарий представляет ясную волнообразность как во времени, так и в пространстве. Например, в таблице 5 первично угашаемый пункт в течение 12 минут оказывался то заторможенным, то свободным от торможения, – то же можно видеть и в пространстве на разных пунктах мозга в одно и то же время. Это капитальный факт, с которым мы будем в дальнейшем встречаться все чаще и чаще. Он является совершенно естественным результатом встречи, уравновешивания двух противоположных нервных процессов, раздражения и торможения, результатом, совершенно аналогичным, например, волнам третьего рода кривой кровяного давления, как результату взаимодействия прессорных и депрессорных иннерваций.
Та же волнообразность выступает на той же собаке (также в опытах Подкопаева) и при гашении до нуля и именно на пункте, ближайшем к угашаемому. На дальнем пункте она не была заметна. Может быть, за ее слабое выражение надо считать в соответствующем периоде времени остающийся в течение 7 минут без изменения уровень торможения.
Вот этот опыт.
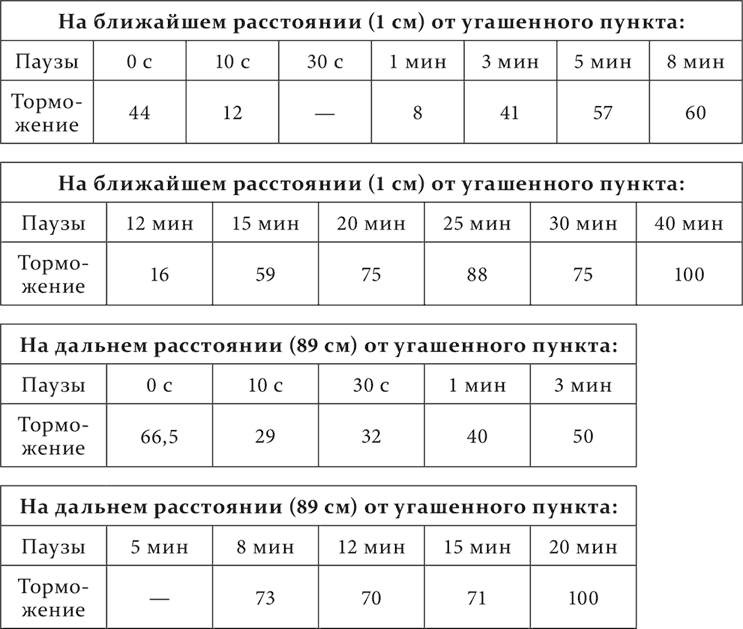
Торможение вычислено в процентах остающегося нормального положительного эффекта.
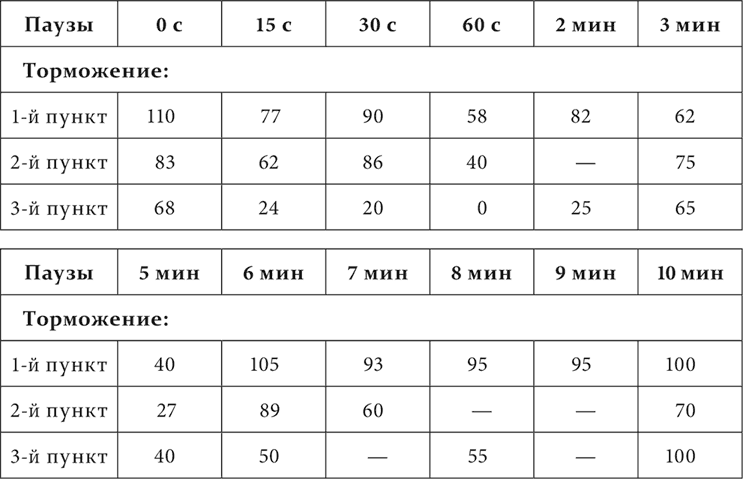
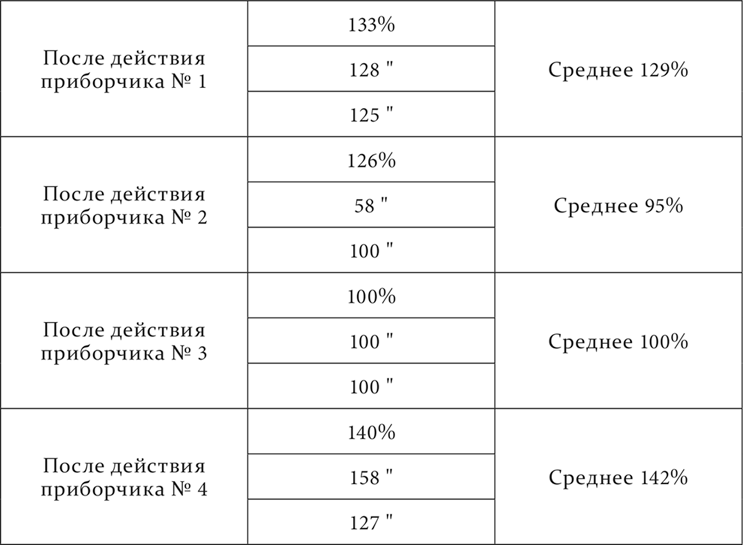
Волнообразность и вообще еще более сложные отношения между раздражением и торможением на разных пунктах кожного анализатора и при разных паузах после применения тормозного раздражения обнаружились на следующем нашем животном (опыты Андреева). По длине задней конечности с верхней трети бедра до нижней части голени прикреплялись приборчики для механического раздражения кожи, числом четыре, №№ 0, 1, 2 и 3, считая сверху вниз. Расстояние между ними приблизительно одинаковое, около 15 см. Раздражения тремя нижними приборчиками в отдельности были положительными условными пищевыми раздражениями, а раздражение верхним – тормозным, дифференцировкой. Дифференцировка не была особенно прочной. Общий ход опыта был таков. Паузы между раздражениями были постоянно семиминутные. Опыт начинался с условного раздражения звонком или ударами метронома. Затем применялось два положительных раздражения приборчиками, безразлично какими, за ними следовало тоже положительное кожно-механическое раздражение, специально испытуемое в данном экспериментальном сеансе. Теперь приводился в действие верхний приборчик, производивший торможение, а после него, на этот раз через разные паузы, от 0 до 12–16 минут, снова применялось кожно-механическое раздражение, последнее перед дифференцировкой, и на нем определялось влияние дифференцировки. Это влияние вычислялось в процентах по отношению к нормальной величине эффекта положительного кожно-механического раздражения, принимаемой за 100. За нормальную величину принималась средняя величина эффекта трех кожно-механических раздражений, предшествующих дифференцировке. Вот числовой результат опытов:
Для более легкого и быстрого представления об оказавшейся в этих опытах сложности отношений тот же результат воспроизвожу в виде кривых.
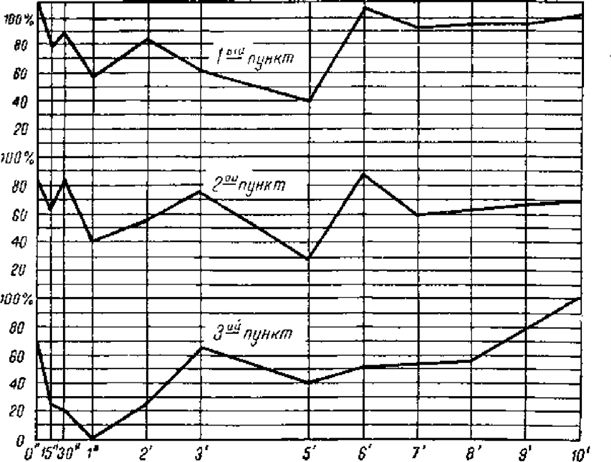
Как видите, влияние тормозного процесса, производимого раздражением на пункте 0, на всех положительных пунктах оказывалось гораздо более различным, чем сходным. Самое общее сходство состояло в том, что торможение рано или поздно достигало максимума, а затем исчезало. Затем оно сказалось и в том, что на всех этих пунктах в одно и то же время обнаружились два максимума торможения: первый к концу первой минуты по прекращении тормозного раздражения, второй к концу пятой минуты. На этом сходство кончилось, а потом выступили разницы, и значительные. На первом, ближайшем к тормозному пункте сейчас же по прекращении тормозного раздражения было небольшое повышение возбудимости – положительная индукция. На двух остальных пунктах ее не было, началось прямо с торможения. На первом и втором пунктах второй максимум торможения был больше первого, на третьем пункте первый максимум представлялся полным торможением, совершенным исчезанием положительного эффекта, второй обозначился слабо. Затем на первом и втором пунктах происходило много колебаний, и очень резких. На третьем пункте было только довольно значительное ослабление торможения на третьей минуте. Без него ход последовательного торможения был бы такой же регулярный, как мы познакомились с ним раньше в девятой и десятой лекциях. На этом третьем пункте опять так же, как это мы видели раньше, период достижения максимума был в несколько раз короче периода полного исчезания торможения, на первом же и втором пунктах период достижения наибольшего максимума был или равен, или даже меньше периода полного возвращения нормальной возбудимости. Наконец, если взять валовое последовательное торможение, т. е. если сложить все проценты торможения, обозначенные в таблице, то окажется, что торможение всего более дало себя знать на третьем пункте, менее на втором пункте и всего меньше на первом пункте.
«Перед нами – положительная индукция, сперва возникающая поодаль от исходного пункта торможения и обнаруживающаяся более поздно, а затем постепенно подвигающаяся по направлению к исходному пункту и все быстрее появляющаяся после начала торможения, т. е. как бы постепенно преодолевающая и вытесняющая и во времени и в пространстве тормозной процесс»
Эту сложность в распределении возбудимости на разных пунктах кожного анализатора под влиянием тормозного процесса всего естественнее было бы понять так, что индуцированное повышение возбудимости дало себя знать всего больше около пункта, где внешним раздражением было вызвано торможение, и потому здесь сильнее выступили волны борьбы, уравновешивания двух противоположных процессов. А что здесь действительно индуцированный раздражительный процесс был всего значительнее, доказывается фактом явной положительной индукции, которая только на этом пункте и наблюдалась сейчас же по прекращении тормозного раздражения. На дальнем пункте торможение обнаружилось наиболее беспрепятственно, проделав свое обыкновенное и постепенное усиление до максимума, а затем более растянутое во времени ослабление до возвращения нормального эффекта.
С методической стороны в этом случае надо, однако, указать, что отдельные цифры, из которых были получены приведенные средние цифры, относящиеся к ближнему пункту, расходились между собой гораздо больше, чем цифры, относящиеся к среднему и в особенности к дальнему пунктам. Может быть, отчасти на это следует отнести менее значительные разницы в кривых разных пунктов. Но, может быть, это самое непостоянство отдельных цифр обусловливалось именно большими колебаниями возбудимости на ближнем пункте, чем на дальнем.
Наконец, я приведу еще опыты (Подкопаева), опять же на новом животном, которые так же резко подчеркнут большую сложность разбираемых отношений.
На пути движения того или другого нервного процесса перемежаются районы то с повышенной, то с пониженной возбудимостью, с преобладанием то раздражительного процесса, то тормозного»
У собаки на одной половине тела по одной линии расположены приборчики для механического раздражения кожи на запястье (№ 0), на плече (№ 1), на груди (№ 2), на животе (№ 3) и на бедре (№ 4). На запястье выработан положительный условный пищевой рефлекс, на остальных – тормозные, между собой выравненные. После действия каждого из тормозящих приборчиков непосредственно, т. е. без всякого промежутка времени, испытывается положительно действующий приборчик. С начала опыта определяется его нормальный эффект. Опыты с действием разных тормозящих приборчиков на эффект положительно действующего ставились беспорядочно то с одним, то с другим, так что полученный результат отнести на счет фазовых изменений с течением времени нет оснований.
В прилагаемой таблице приведены как отдельные опыты, так и среднее из них. Эффект положительно действующего приборчика вычислен в процентах по отношению к его нормальной величине.
Из таблицы ясно, что состояние положительно действующего пункта очень различно в зависимости от того, действие какого из тормозящих приборчиков ему предшествовало. При раздражении ближайшего и отдаленнейшего пункта наступала резкая положительная индукция: значительно увеличивалось слюноотделение сравнительно с нормой, укорачивался латентный период и часто усиливалась пищевая двигательная реакция, причем индуцирующее действие дальнего пункта было несколько больше, чем ближнего. От третьего пункта не наблюдалось никакого действия, а от второго – чрезвычайно смешанное: тормозящее, положительно индуцирующее и никакого. Сходство отдельных цифр при повторении тождественных опытов в трех рядах так велико, что нет основания заподозрить их точность, и это же располагает отнестись с доверием и к цифрам, относящимся к второму пункту, считая, что он представляет какую-то переходную зону.
«Изменения возбудимости различных пунктов коры под влиянием внешних раздражений, специально под влиянием тормозного раздражения, происходят волнообразно как во времени на одном и том же пункте, так и в пространстве на разных пунктах в одно и то же время»
Как понимать этот результат? Если считать, что линейное расположение наших пунктов на коже соответствует такому же расположению проекционных пунктов в коре, то представленный результат надо считать ярким доказательством волнообразного состояния возбудимости в пространстве. Мы должны будем признать, что на пути движения того или другого нервного процесса перемежаются районы то с повышенной, то с пониженной возбудимостью, с преобладанием то раздражительного процесса, то тормозного. Ввиду важности этого вывода необходимо иметь специальные доказательства, что принимаемая нами соответственность расположения пунктов на коже и в коре есть точный факт.
Весь приведенный в текущей лекции материал более или менее согласно свидетельствует, что изменения возбудимости различных пунктов коры под влиянием внешних раздражений, специально под влиянием тормозного раздражения, происходят волнообразно как во времени на одном и том же пункте, так и в пространстве на разных пунктах в одно и то же время. И это не должно казаться неожиданностью, раз мы имеем несомненные факты, с одной стороны, движения нервных процессов, с другой – их взаимной индукции. Остается только большим вопросом общее точное правило, по которому это происходит: почему один раз выступает волнообразность, в другой раз ее нет, почему в одних случаях распространяющееся торможение предшествуется положительной индукцией, в других ее не видно, и т. д.? Сейчас мы стоим перед беспорядочным разнообразием фактов. Но имеющийся в наших руках материал позволяет заключить, что это разнообразие обусловливает соучастие трех факторов. Это: особые типы нервной системы наших экспериментальных животных, фазы установки новых отношений в коре под влиянием внешних раздражений и виды установки, так как, например, наши случаи внутреннего торможения очень различаются между собой по их напряженности и стойкости. Серьезная задача будущего полного анализа исследуемых отношений и заключается в точном определении участия в данный момент деятельности коры каждого из указанных факторов, сведя все на силовые отношения нервных процессов. Постепенное приближение к этой задаче, конечно, возможно и в настоящее время. Уже в описанных опытах можно указать на некоторые намечающиеся частичные подходы к решению нашей задачи. Но еще отчетливее выступает это в опытах, которыми мы занимались и занимаемся в последнее время. Мы видели постоянно в последних четырех лекциях, что торможение, производимое соответственным раздражением в определенном пункте, начинает прямо или после предварительной кратковременной положительной индукции распространяться по анализатору лишь по прекращении тормозного раздражения. Почему так? Как создается такое положение дела? Ради ответа на этот вопрос мы поставили, очевидно до сих пор недостававшие в нашем плане исследования предмета, опыты.
Положительная индукция, возникающая под влиянием тормозного раздражения определенного пункта, ограничивает распространение тормозного процесса»
Мы испытывали другие пункты анализатора, или другие анализаторы, не по прекращении тормозного раздражения, как всегда делали это раньше, а во время продолжающегося тормозного раздражения, на фоне его, как выражался Подкопаев, первым разрабатывавший этот вопрос. Испытание было произведено на четырех различных собаках. Прежде всего опишу опыты (Подкопаева) с угасательным торможением.
Это та же собака, которая служила для приведенных в текущей лекции опытов с иррадиацией торможения после однократного неподкрепления условного рефлекса и после угашения до нуля. В описываемых теперь опытах угашение условного механического раздражения на определенном месте кожи было полное. Раздражение других пунктов производилось или на ближайшем к угашенному пункте (1 см расстояния), или на дальнем (89 см расстояния). Это присоединение происходило на тридцатой секунде продолжающегося тормозного раздражения, причем совместное раздражение также продолжалось 30 секунд. На ближайшем пункте эффект такого раздражения был 84,5 % нормального, на дальнем 88 %, т. е. одинаковый, в то время как по прекращении тормозного раздражения через 0 секунд промежутка, как это мы видели раньше, раздражение ближайшего и дальнего пунктов соответственно давало 44,5 % и 66,5 %.
Для полной оценки этого результата надо еще прибавить, что латентный период рефлексов при одновременном раздражении угашенного пункта с другими пунктами был значительно короче, чем в норме, а также и двигательная пищевая реакция резче, чем обыкновенно. И то и другое – бесспорные указания на повышение возбудимости, положительное индуцирование остальных пунктов анализатора, кроме специально угашенного. Несколько меньший слюноотделительный эффект от остальных пунктов, очевидно, надо понимать как результат алгебраического суммирования их эффекта с эффектом из угашенного пункта.
Особенный интерес для нас в этих опытах представляет еще следующее. Угашение до нуля на определенном месте кожи производилось повторением неподкрепляемого условного раздражения через 2 минуты. В новой вариации опытов по получении первого нуля процедура угашения продолжалась, и к третьему нулевому раздражению присоединялось раздражение других пунктов кожи. Теперь эффект рефлекса от соединенного раздражения был большей частью совершенно равен нормальному эффекту изолированного положительного раздражения. Значит, углубление торможения, усиление тормозного процесса повело к более значительной положительной индукции. Рядом с этим в тех же опытах обращает на себя внимание и следующее сопоставление фактов. У этой собаки, как мы видели, к 2 минутам после угашения до нуля иррадиированное торможение на кожном анализаторе еще держится на уровне не ниже 50 % полного торможения, между тем как при совместном раздражении угашаемого пункта в третий раз до нуля (через 2 минуты после предшествующего угашения) и остальных пунктов не видно ни малейшего знака торможения. Следовательно, оно во время повторного тормозного раздражения опять сконцентрировалось в пункте этого раздражения.
В той же работе (Подкопаева) те же результаты были получены на другой собаке, у которой были, между прочим, выработаны положительные пищевые условные рефлексы на определенный тон и удары метронома (120 ударов в минуту) и тормозной (дифференцировка на 60 ударов метронома в минуту). На «фоне» дифференцировочного торможения от метронома тон из 9 проб только один раз дал мало заторможенный эффект, в 4 случаях – нормальную величину его и еще в 4– величину, даже значительно превосходившую норму. Для следующих опытов, относящихся к той же теме, были употреблены две собаки. У одной (опыты В. П. Головиной) среди многих пищевых условных рефлексов были: положительные на метроном, определенный тон, свисток, лампочку, вспыхивающую в определенном месте, и механическое раздражение кожи и три тормозных дифференцировки – на удары метронома другой частоты, на другое место кожи и на другое место вспыхивающей лампочки. Когда все эти рефлексы были хорошо выработаны, применялось то или другое положительное раздражение совместно с каким-нибудь из указанных тормозных раздражений. Все эти пробы дали совершенно однообразный результат. Положительные раздражители в этих комбинациях всегда имели или нормальный для них по величине эффект, или несколько увеличенный, и более часто с укороченным латентным периодом.
«Торможение, производимое соответственным раздражением в определенном пункте, начинает прямо или после предварительной кратковременной положительной индукции распространяться по анализатору лишь по прекращении тормозного раздражения»
У другой собаки (опыты Павловой) также среди многих условных рефлексов были положительные – на свисток, звук бульканья и механическое раздражение кожи и тормозной – дифференцировка на удары метронома. В опытах над этим животным каждое из указанных положительных раздражений по нескольку раз применялось с одним и тем же тормозным раздражением. Здесь результат получился несколько иной. Хотя латентный период также большей частью был укорочен, но величина эффекта комбинированного раздражения почти постоянно была меньше, и часто значительно, сравнительно с величиной эффекта одиночного положительного раздражения. Значит, у этой собаки концентрирование нервных процессов было менее совершенным.
Нужно прибавить, что эти две последние собаки явно принадлежали к двум разным типам нервной системы. В то время как у первой дифференцировки были вообще очень прочны и не страдали от комбинированных раздражений, у второй дифференцировка часто сама по себе была неполной и всегда еще более нарушалась после комбинации с положительными раздражениями.
«Взаимодействие между раздражительным и тормозным процессами определяется то фазой установки новых отношений в коре, то типом нервной системы разных животных»
Опыты последнего ряда ясно показывают, с одной стороны, как положительная индукция, возникающая под влиянием тормозного раздражения определенного пункта, ограничивает распространение тормозного процесса, с другой – свидетельствуют, что взаимодействие между раздражительным и тормозным процессами определяется то фазой установки новых отношений в коре (опыт Подкопаева), то типом нервной системы разных животных (опыты Головиной и Павловой).
Назад: Лекция одиннадцатая
Дальше: Лекция тринадцатая

