Книга: Медицинская микробиология, иммунология и вирусология
Назад: Глава 34 Основные популяции иммунокомпетентных клеток. Т-лимфоциты и их функции
Дальше: Глава 36 Кооперативное взаимодействие макрофагов, Т– и В-лимфоцитов в выдаче иммунного ответа
Глава 35
В-лимфоциты и их функции
Как уже отмечено выше, B-лимфоциты помимо антигенов Ly, общих с T-лимфоцитами, имеют свойственные только им антигены Lyb. B-лимфоциты подразделяют на Lyb5+, т. е. имеющие этот антиген, и Lyb5– , т. е. лишенные его. Разные антигены – ТЗ, ТН-1 и ТН-2 (см. с. 259) – активируют различные субпопуляции B-лимфоцитов. Так, ТЗ– и ТН-1-антигены активируют как Lyb5+, так и Lyb5–-B-лимфоциты. В свою очередь, TH-2-антигены активируют только Lyb5+-В-лимфоциты. B-лим+фоциты подразделяют на следующие -шесть основных популяций: 1) клетки Bla (CD5 ), или просто Bla; 2) клетки Blb (CD5 ), или Blb. Клетки Bla и Blb объединяют в одну группу B-1-клеток, потому что они первыми появляются в онтогенезе; 3) клетки B-2, которые затем дифференцируются в АОК; 4) MZ-B, или B-клетки маргинальной зоны (от лат. margo – край); 5) B-супрессоры и 6) B-киллеры. Клетки Bla, Blb и MZ-B продуцируют только IgM, т. е. неспецифические иммуноглобулины. Все они осуществляют функции 1-й линии защиты организма от инфекций, т. е. опосредуют видовой иммунитет. Лимфоциты B-2 играют решающую роль в формировании специфического приобретенного, или адаптивного иммунитета. B-2 лимфоциты рециркулируют между кровью и лимфой (MZ-B-лимфоциты – не циркулирующие клетки) и в лимфатических фолликулах селезенки и лимфоузлов встречаются с антигенами, связанными с дендритными фолликулярными клетками. Дендритные клетки, как и все макрофаги, обладают способностью осуществлять процессинг и представление антигена. Здесь образуются зародышевые центры, в которых и происходит превращение B-2-лимфоцитов в антителообразующие клетки и формирование клеток иммунной памяти. B-супрессоры выполняют такие же функции, как и T-супрессоры. B-киллеры взаимодействуют с Fc-фрагментами IgG, фиксированных на клетках трансплантата, и разрушают их.
Мембраны B-лимфоцитов несут большое количество рецепторов (более 40 типов) с разнообразными свойствами. В частности, они имеют рецепторы к Fc-компоненту иммуноглобулина, к C3-компоненту комплемента. Благодаря этим рецепторам B-клетки легко выявить с помощью метода розеткообразования или методом бляшек. Суть последнего заключается в том, что если эритроциты барана использовать в качестве антигена для иммунизации животных, то в их лимфоидной ткани будут накапливаться B-лимфоциты, вырабатывающие соответствующие антитела. Добавление таких клеток в среду, содержащую эритроциты, приведет к тому, что комплексы эритроцит + антитело + комплемент будут связываться с лимфоцитом и лизироваться вблизи него, образуя пятно (бляшку). Химическая природа B-лимфоцитов рецепторов была выяснена в 1969 г. Р. Кумбсом, который установил, что они являются иммуноглобулинами.
В-лимфоциты несут на своей поверхности большое количество таких рецепторов до 150 000 на один лимфоцит. При этом все рецепторы данного лимфоцита обладают одной и той же антительной специфичностью, т. е. один лимфоцит может реагировать только с одним антигенным детерминантом. Вместе с тем установлено, что В-лимфоциты раньше всех начинают синтезировать иммуноглобулины класса IgM. Зрелые лимфоциты могут нести рецепторы, относящиеся более чем к одному классу. Например, IgM + IgD, IgM + IgG и т. п. Однако во всех случаях такие иммуноглобулины обладают только одной антительной специфичностью.
В-лимфоциты представлены громадным количеством клонов (вероятно, более чем 108). Клон – совокупность генетически идентичных клеток. Клетки данного клона несут рецепторы одной и той же антительной специфичности, поэтому каждый клон отличается от других по антительной специфичности своих рецепторов. Такие клоны формируются из исходных клеток-предшественников В-лимфоцитов.
В ходе своего созревания (от эмбриональной клетки до рождения), а также после рождения В-лимфоциты подвергаются дифференцировке, биологический смысл которой состоит в создании клона клеток, синтезирующих и секретирующих антитела, специфически взаимодействующие с данным антигеном.
Процесс созревания В-лимфоцитов включает в себя две стадии:
1. Антигеннезависимую, которая протекает в эмбриональном периоде.
2. Антигензависимую, которая протекает после рождения и наступает только после встречи с соответствующим антигеном.
Антигеннезависимая стадия дифференцировки
Она начинается с раннего предшественника В-лимфоцита (ранний предшественник В-лимфоцита, в свою очередь, возникает из исходной полипотентной кроветворной клетки, он же является одновременно и предшественником Т-лимфоцитов) и заканчивается образованием зрелых В-лимфоцитов, которые несут на своей мембране иммуноглобулиновый рецептор одной антительной специфичности.
В ходе антигеннезависимой дифференцировки В-лимфоцита происходит последовательное формирование генов вначале для Н-цепи, затем для L-цепи, сборка полного гена иммуноглобулина М, образование вначале цитоплазматического IgM и, наконец, мембранных иммуноглобулиновых рецепторов IgM и IgD.
В формировании полного гена Н-цепи принимают участие следующие гены, расположенные в определенном локусе хромосомы на значительном расстоянии друг от друга: V (200 вариантов), D (20 вариантов), J (4 – 6 вариантов), С (9 вариантов). Вначале происходит выбор одного из генов J– и одного из D-сегмента, затем соединение одного из V-генов с D-J-сегментом. Образующийся при этом тандем V-D-J сближается с геном Сμ. В результате этого происходит выбор одного из множества возможных вариантов генов V, D, J, т. е. такого, который будет контролировать структуру активного центра данного антитела. Образовавшийся ген вариабельной части Н-цепи ассоциирует с геном Сμ, и возникает полный ген Н-цепи μ-типа. Его транскрипция приводит к образованию цитоплазматической Н-цепи типа Сμ. Наличие этой цепи в цитоплазме является характерным признаком пре-В-лимфоцита. Он является пролиферирующей клеткой, поэтому происходит накопление клона таких клеток, которые имеют более широкий спектр специфичности, чем зрелые В-лимфоциты. Широкая специализация зрелых В-лимфоцитов будет зависеть от ассоциации Н– и L-цепей. Следующая стадия – превращение пре-В-лимфоцита в незрелый В-лимфоцит – характеризуется сборкой гена для L-цепи, формированием полного гена иммуноглобулина IgM и синтезом мембранной (рецепторной) формы иммуноглобулина М. Для образования рецепторной формы иммуноглобулина надо, чтобы Н-цепь включила гидрофобный «якорь». Он представляет собой пептид из 41 аминокислотного остатка, контролируемый фрагментом μ-гена, отделенным интроном от его последнего экзона. Сборка гена для L-цепи происходит по такому же типу, как и генов Н-цепи. Регуляция образования полных генов Н-цепи и L-цепи осуществляется сплайсингом ядерных пре-мРНК этих генов.
Конечная стадия антигеннезависимой дифференцировки – образование зрелого В-лимфоцита. Он характеризуется наличием двух мембранных рецепторов, обладающих одной антительной специфичностью: IgM и IgD. Синтез IgD также контролируется на уровне сплайсинга, при этом происходит переключение тандема V × D × J × Cμ на V × D × J × Cδ – класспереключающая рекомбинация. Формированием рецепторного IgD завершается процесс антигеннезависимой дифференцировки предшественника В-лимфоцита в зрелый В-лимфоцит.
Зрелый В-лимфоцит имеет два рецептора (IgM и IgD) с одинаковыми активными центрами. Таким образом возникают клоны лимфоцитов, каждый из которых обладает своей антительной специфичностью и способен распознать только «свой» антиген. Зрелый В-лимфоцит – покоящаяся клетка, т. е. он находится в фазе цикла G0. Однако он способен к дальнейшей пролиферации, дифференцировке и превращению в антителообразующую клетку и в клетку памяти. Все стадии антигеннезависимой дифференцировки В-лимфоцита происходят в тесном контакте с клетками окружающих тканей. Созревание В-лимфоцитов в эмбриональном периоде происходит в печени. После рождения исходные стволовые клетки перемещаются в костный мозг, здесь и происходит дальнейшее образование зрелых В-лимфоцитов с высокой скоростью – 108 клеток в сутки.
Зрелые В-лимфоциты покидают костный мозг и заселяют селезенку, лимфатические узлы и другие скопления лимфатических клеток, где их дифференцировка «замораживается» до встречи с соответствующим антигеном, т. е. они уже полностью созрели для реализации своих функций, для этого им нужна только встреча с антигеном.
Антигензависимая дифференцировка В-клеток
Превращение зрелых В-лимфоцитов в антителообразующие клетки и клетки иммунной памяти происходит после встречи лимфоцита с антигеном. Антигензависимая дифференцировка складывается из трех основных событий: 1) активация В-клеток, 2) их пролиферация, 3) дифференцировка.
Биологический смысл активации В-лимфоцитов заключается в том, что они, вопервых, переходят из покоящегося состояния в начальную стадию клеточного цикла G1, и, во-вторых, в том, что в результате активации они приобретают способность вступать во взаимодействие с другими клетками (Т-лимфоцитами) и воспринимать те сигналы, которые они им передают.
Для само́й активации необходимы по крайней мере два сигнала: превый – для перевода клетки из фазы покоя (G0) в G1-фазу клеточного цикла, а второй – для вступления ее в S-фазу. Роль первого сигнала выполняют различные антигены или митогены. Он воспринимается Ig-рецептором. В качестве второго сигнала выступают различные лимфокины, в том числе интерлейкины-1, -2, -4, -6. В результате их воздействия происходит активация генома клетки, что и обусловливает пролиферацию и дифференциацию В-лимфоцитов в антителообразующие клетки (АОК). Механизм передачи сигнала от Ig-рецептора геному В-клетки во многом совпадает с таковым у Т-лимфоцитов.
Активация В-клеток может осуществляться разными путями, что зависит как от участия в ней других клеток (макрофагов, Т-хелперов), так и от свойств самих антигенов. Их природа может определять интенсивность и длительность сигнала, подаваемого в клетку рецептором В-лимфоцита. Если он недостаточен, требуется помощь Т-хелперов. Например, бактериальные полисахариды могут прямо активировать В-клетки и индуцировать выдачу сильного ответа. Такие антигены получили название Т-независимых. Напротив, вирусные и растворимые антигены вызывают слабый ответ В-лимфоцитов, и для их активации требуется помощь Т-хелперов. Такие антигены называют Т-зависимыми (ТЗ). Комплекс антигена с антителом является более слабым активатором В-лимфоцитов, чем сам антиген. Поэтому, когда антитела нейтрализуют большинство молекул антигена, происходит постепенное торможение образования антител. Т-независимые антигены, в свою очередь, делят на два типа: Т-независимые антигены 1 (ТН-1) и Т-независимые антигены 2 (ТН-2). Различие между ними состоит в том, что ТН1-антигены вызывают иммунный ответ как Lyb5+-В-лимфоцитов, так и Lyb5–, в то время как ТН2-антигены способны индуцировать иммунный ответ только Lyb5+-В-лимфоцитов при участии макрофагов.
Известны следующие пути активации В-лимфоцитов:
1) Т-зависимым антигеном с помощью белков МНС класса II;
2) Т-независимым (т. е. без участия Т-лимфоцита) антигеном, который в своем составе имеет митогенный компонент;
3) Т-независимым антигеном, не содержащим митогенного компонента;
4) поликлональным активатором (липополисахаридом);
5) антииммуноглобулинами μ.
Основным выступает механизм активации с помощью Т-лимфоцитов.
Процесс антигензависимой дифференцировки В-лимфоцитов находится под контролем интерлейкинов. Все они полифункциональны и их действие не ограничивается только регуляцией иммунного ответа. Сам процесс антигензависимой дифференцировки протекает так: В-лимфоцит с помощью иммуноглобулиновых рецепторов распознает и связывает антиген. Это стимулирует его переход из покоящегося состояния (G0) в фазу G1. В-клетка увеличивается, на ее мембране образуются новые рецепторы: один – к фактору роста, другой – к фактору дифференцировки, третий – к фактору, замещающему Т-клетки.
В-лимфоциты в ряде случаев под действием антигена могут дифференцироваться в АОК без пролиферации, а под воздействием особого фактора созревания В-клеток. Одновременно с В-лимфоцитом антиген по представлению макрофага распознается Т-лимфоцитом-хелпером, который также активизируется и начинает синтезировать и секретировать факторы роста и дифференцировки. Последние взаимодействуют с рецепторами В-клетки и переводят ее в следующую стадию клеточного цикла (S-фазу) и в фазу дифференцировки. Факторы пролиферации нужны в течение первого и всех последующих циклов деления В-лимфоцитов, а факторы дифференцировки, очевидно, только на последних этапах ее.
Активированная В-клетка претерпевает 8 – 10 делений и одновременно дифференцируется в антителообразующую клетку (плазмоцит). В процессе дифференцировки ядро В-лимфоцита уплотняется, а объем цитоплазмы увеличивается. В ней развивается сеть мембран эндоплазматического ретикулума, и образуется большое количество связанных с мембранами рибосом. Такая клетка синтезирует и секретирует антитела одной антительной специфичности. 90 – 96 % всего производимого ею белка представляет антитело с одинаковым активным центром против антигена, который вызвал антигензависимую дифференцировку данного В-лимфоцита. Так возникает клон антителообразующих клеток.
Такой путь активации и дифференцировки проходит популяция В-лимфоцитов, имеющих антиген Lyb5, т. е. примерно 50 % всей популяции В-клеток. Клетки, лишенные Lyb5-антигена (Lyb5–), нуждаются для активации в прямом контакте с активированными Т-хелперами, которые выделяют в этом случае антигенспецифический фактор дифференцировки. Такие В-лимфоциты тоже превращаются в антителообразующие клетки.
Антигены, вызывающие агрегацию иммуноглобулиновых рецепторов на мембране В-лимфоцитов, например липополисахарид, полианионы, антитела к μ-цепи; и антигены, несущие повторяющиеся детерминанты в своей молекуле, способны активировать В-клетки без участия Т-лимфоцитов, но эти механизмы играют незначительную роль в иммунном ответе.
Происхождение и дифференцировка клеток иммунной системы
Не все индуцированные антигеном В-лимфоциты подвергаются дифференцировке до конца. Часть из них после нескольких циклов деления перестает размножаться и образует субклон клеток памяти (из одной В-клетки образуется около 1000 клеток памяти, таким же образом образуются клетки памяти и из Т-лимфоцитов). Клетки памяти определяют продолжительность приобретенного иммунитета. При повторном контакте с данным антигеном они быстро превращаются в клетки-эффекторы. При этом В-клетки памяти обеспечивают синтез антител в более короткие сроки, в большем количестве и с более высоким сродством антител другого класса иммуноглобулинов – IgG вместо IgM. Эта перестройка происходит благодаря рекомбинации генов Н-цепи: тандем генов V × D × J переносится с Сμ-гена к одному из СН-генов – γ, α, ε. Ее вызывает сигнал, получаемый СD40-рецептором В-клеток памяти, когда они выходят из зародышевого центра в пул памяти, от gp39-рецептора Т-хелперов. Установлено, что мутации в гене, кодирующем gp39, являются причиной редкого тяжелого первичного иммунодефицита – гипер-IgM-синдрома. Отсутствие у Т-хелперов молекул gp39 в этих случаях приводит к утрате ими способности связываться с CD40, что ведет к неправильному ответу на Т-зависимый антиген – гиперпродукции IgM.
В ходе антигензависимой дифференцировки В-лимфоцитов используется и механизм соматических мутаций в V-генах. Они происходят с частотой в 10 000 раз большей частоты спонтанных мутаций и ограничиваются определенной стадией дифференцировки, а именно – периодом перехода от продукции IgM к продукции IgG. Благодаря этим мутациям обеспечивается максимальная подгонка структуры активного центра антитела к детерминанту антигена.
Таким образом, наиболее важными событиями дифференцировки В-лимфоцитов являются: 1) сборка гена иммуноглобулина из его фрагментов, содержащихся в ДНК эмбриональных клеток; 2) формирование пула клеток памяти; 3) возникновение новых вариантов генов Ig в результате дополнительных класс-переключающих рекомбинаций; 4) вспышка соматических мутаций на строго определенной стадии дифференцировки. В результате этих событий происходит образование множества генетически стабильных клонов антителообразующих клеток (вероятно, не менее чем 108). Общая схема происхождения и дифференцировки Т– и В-лимфоцитов и макрофагов из исходных стволовых клеток представлена на рис. 71.
В соответствии с этой схемой, исходная костно-мозговая клетка (HSC) генерирует два типа предшественников: лимфоидную стволовую клетку (LSC), от которой происходят клетки-предшественники Т-лимфоцитов (PTC), клетки-предшественники В-лимфоцитов (PBC); и клетку, являющуюся предшественником клеток красной крови, от которой, в свою очередь, происходит предшественник лейкоцитов (CFUc) и берет начало система мононуклеарных макрофагов. Предшественники Т-лимфоцитов под влиянием тимуса превращаются в Т-лимфоциты и их субклассы. Пути дифференцировки В-лимфоцитов описаны выше.
В целом система В-лимфоцитов обеспечивает синтез антител, отвечает за иммунитет против большинства бактериальных и вирусных инфекций, анафилаксию и другие реакции гиперчувствительности немедленного типа, некоторые аутоиммунные болезни, за формирование клеток иммунной памяти и иммунологическую толерантность.
Система T-лимфоцитов играет регуляторную роль по отношению к B-лимфоцитам, отвечает за все реакции гиперчувствительности замедленного типа, иммунитет против вирусных и некоторых бактериальных инфекций (туберкулез, бруцеллез, туляремия и др.), осуществляет иммунологический надзор, отвечает за противоопухолевый иммунитет, иммунологическую толерантность, некоторые виды иммунопатологии.
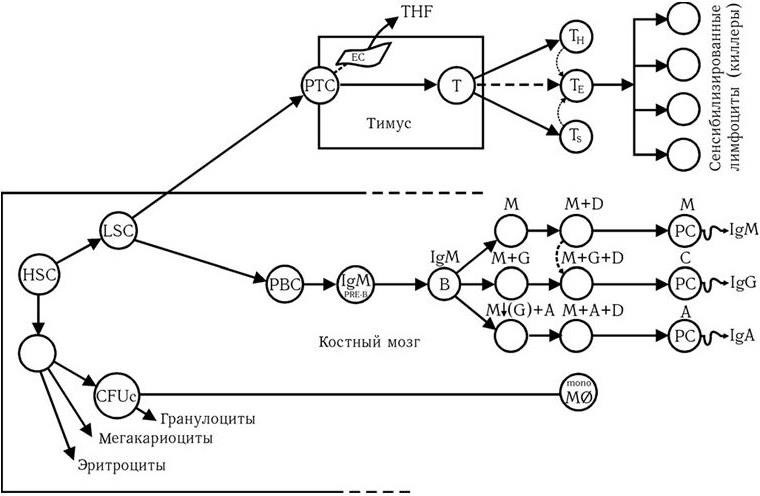
Рис. 71. Схема происхождения и дифференцировки клеток-эффекторов иммунной системы (ВОЗ, 1978).
HSC – костно-мозговая стволовая кроветворная клетка; LSC – лимфоидная стволовая клетка; PTC – предшественник Т-клеток; PBC – предшественник В-клеток; TE – Т-эффекторы; TH – Т-хелперы; TS – Т-супрессоры; CFUc – кроветворный предшественник макрофагов; PC – плазматическая клетка; EC – эпителиальная клетка; THF – тимусный гуморальный фактор
Вместе с тем T– и B-клетки являются двумя частями единой иммунной системы организма. Поэтому деление иммунитета на гуморальный и клеточный носит весьма условный характер, так как антитела синтезируются В-клетками, а Т-лимфоциты и другие клетки осуществляют свою иммунокомпетентность через синтезируемые ими гуморальные факторы (цитокины, лимфокины, интерлейкины и др.).
Координированное взаимодействие макрофагов, Т– и В-лимфоцитов при встрече с антигеном обеспечивает выдачу адекватного иммунного ответа.
Назад: Глава 34 Основные популяции иммунокомпетентных клеток. Т-лимфоциты и их функции
Дальше: Глава 36 Кооперативное взаимодействие макрофагов, Т– и В-лимфоцитов в выдаче иммунного ответа

