Книга: Медицинская микробиология, иммунология и вирусология
Назад: Глава 33 Клеточные основы иммунитета. Органы иммунитета
Дальше: Глава 35 В-лимфоциты и их функции
Глава 34
Основные популяции иммунокомпетентных клеток. Т-лимфоциты и их функции
Основную роль в реализации иммунных функций играют следующие клетки: 1) макрофаги, а также другие антигенпредставляющие, или вспомогательные, или А-клетки (англ. accessory – вспомогательный); 2) популяции В-лимфоцитов; 3) популяции Т-лимфоцитов; 4) популяции лимфоцитов, обладающих естественными цитотоксическими свойствами – способностью разрушать опухолевые клетки, вирусинфицированные клетки, клетки трансплантата и т. п.: NK-клетки, Pit-клетки, нулевые клетки.
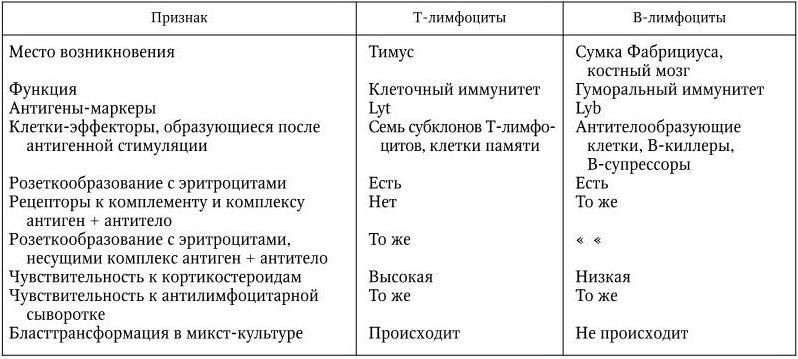
По морфологическим свойствам Т– и В-лимфоциты не отличаются друг от друга. Однако они существенным образом различаются по вкладу в реакции иммунитета, по многим другим свойствам, в том числе структуре и функции рецепторов и по антигенной специфичности (табл. 14).
Таблица 14
Характеристика Т– и В-лимфоцитов
Т-лимфоциты
Самый ответственный момент в процессе иммунного ответа – это распознавание химического маркера, свойственного «чужому» агенту и отличающегося от «своего». Эту роль выполняют макрофаги, антитела, Т– и В-лимфоциты. Антитела распознают антиген с помощью своих активных центров, а макрофаги, Т– и В-лимфоциты – благодаря имеющимся на их мембранах особым рецепторам.
Экспериментальное обоснование предположения о наличии у клеток лимфоидной системы специфических рецепторов было получено в 60-е гг. ХХ в. после обнаружения двух важных феноменов: цитопатического действия лимфоцитов и розеткообразования (иммуного прилипания).
Способность сенсибилизированных лимфоцитов распознавать трансплантационный антиген и активно прикрепляться к клеткам-мишеням послужила прямым указанием на наличие у лимфоцитов рецепторов, подобных по структурной специфичности антителам.
Феномен розеткообразования состоит в том, что если к культуре нормальных или иммунных лимфоцитов добавить какой-либо антиген, например чужеродные эритроциты, то последние, прикрепляясь к лимфоцитам, образуют структуру типа розетки. Это явление высоко специфично: удаление лимфоцитов, образовавших розетки с эритроцитами данного вида животных, лишает популяцию лимфоцитов способности образовывать розетки из этих клеток.
Этот феномен не только свидетельствует о наличии у лимфоцитов особых рецепторов, но и говорит о том, что популяция лимфоидных клеток представлена клонами лимфоцитов, различающимися по специфичности этих рецепторов.
Первоначально Т-лимфоциты по своим функциям были разделены на три субкласса:
1) Т-хелперы, или Т-помощники (англ. help – помогать);
2) Т-киллеры (англ. kill – убивать), или Т-цитотоксические лимфоциты;
3) Т-супрессоры (англ. suppress – подавлять).
Т-хелперы необходимы для превращения В-лимфоцитов в антителообразующие клетки и клетки памяти. Т-киллеры разрушают клетки трансплантата, опухолевые клетки и клетки, инфицированные вирусными, бактериальными и другими антигенами. Т-супрессоры подавляют функции определенных эффекторных Т– и В-клеток и обеспечивают иммунологическую толерантность.
Особенность Т-лимфоцитов состоит в том, что их рецепторы, в отличие от антител и рецепторов В-лимфоцитов, не распознают свободно циркулирующих антигенов. Они распознают только те чужеродные вещества, точнее их пептидные фрагменты, которые представляются им клетками организма через посредство антигенов МНС класса I или класса II. Разные Т-лимфоциты распознают разные собственные антигены (белки МНС). Т-цитотоксические лимфоциты распознают клетки, несущие антигены МНС класса I и чужеродные антигены, представляемые ими. Основное назначение Т-цитотоксических лимфоцитов – обеспечение противовирусного, противоопухолевого и трансплантационного иммунитета. Т-киллеры с помощью своих специфических рецепторов распознают чужеродные антигены, ассоциированные на мембранах клеток с их антигеном МНС класса I, они атакуют такие клетки и уничтожают их.
T-хелперы распознают клетки, имеющие антигены МНС класса II (макрофаги, B-лимфоциты и некоторые другие), и представляемые ими чужеродные антигены. Белки МНС класса II связывают чужеродный антиген в макрофагах в ходе процессинга, выносят его на поверхность мембраны макрофага, и в таком виде этот антиген представляется T-хелперам. Последние также распознают его с помощью своих рецепторов и при участии своих белков МНС класса II. В свою очередь, B-лимфоциты также представляют чужеродные антигены Т-хелперам, используя собственные белки МНС класса II.
Таким образом, T-хелперы осуществляют регуляцию иммунного ответа, стимулируя В-лимфоциты, а также другие T-клетки, специализированные к данному антигену.
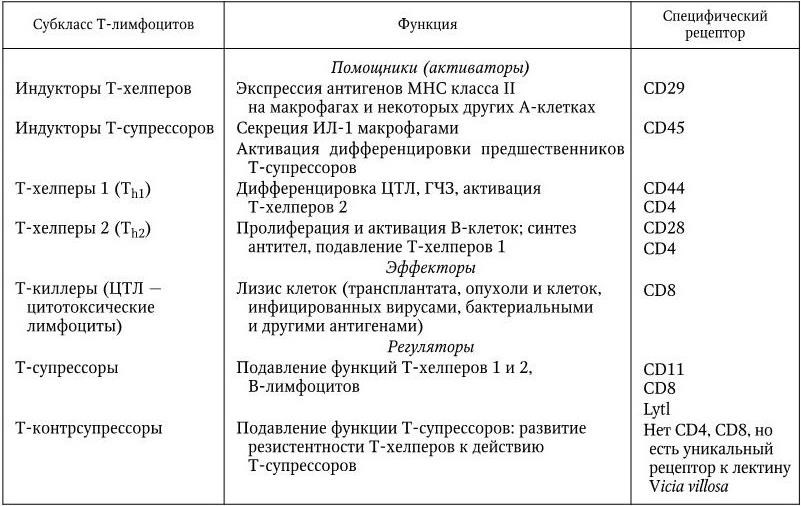
В настоящее время Т-лимфоциты подразделяют на 7 основных субклассов, объединенных в следующие три группы (табл. 15).
1. Т-помощники, или активаторы: индукторы Т-хелперов, индукторы Т-супрессоров, Т-хелперы 1, Т-хелперы 2.
2. Т-эффекторы – Т-цитотоксические лимфоциты. 3. Т-регуляторы: Т-супрессоры, Т-контрсупрессоры.
Кроме того, существуют различные варианты Т-клеток иммунной памяти.
Т-лимфоциты различаются по своим рецепторам. Каждый из субклассов выполняет специфические функции. Для активации Т-клеток требуется контакт их рецептора с комплексом чужеродного антигена с собственным белком МНС класса I или класса II. Через 16 – 20 ч после установления контакта возникают индукторы Т-хелперов. Последние, реагируя на представляемый А-клеткой процессированный антиген, быстро секретируют медиаторы, которые способствуют образованию молекул МНС класса II на поверхности макрофагов и продукции ими IL-1. Индукторы Т-хелперов отличаются от эффекторных Т-хелперов тем, что они имеют особый рецептор CD29, который может быть заблокирован моноклональными антителами, не влияющими на активность Т-хелперов. Только после исполнения функции индукторов данный антиген связывается с белком МНС класса II и возникает комплекс, который узнается рецепторами Т-хелпера. В результате этого на мембране Т-хелпера образуются рецепторы к IL-1. Присоединение к нему интерлейкина-1 приводит ко второму этапу активирования Т-хелпера: он начинает синтезировать и секретировать различные варианты лимфокинов. Существуют две категории Т-хелперов: Т-хелперы 1 (Th1) и Т-хелперы 2 (Th2), которые имеют одинаковые рецепторы CD4 и 4В4, но различаются по другим, а именно: Т-хелперы 1 имеют рецепторы CD45 и CD44, а Т-хелперы 2 – CD28. Их функции различны: Т-хелперы 1 секретируют IL-2, IL-3, фактор некроза опухолей, γ-интерферон и другие цитокины, которые способствуют созреванию Т-цитотоксических лимфоцитов после их взаимодействия с комплексом чужеродный антиген + молекула МНС класса I. Таким образом, Т-хелперы 1 обеспечивают дифференцировку и пролиферацию эффекторных Т-киллеров, способных распознавать и уничтожать как клетки аллотрансплантата, так и свои собственные, если на их поверхности содержатся антигены вирусов, других патогенов или опухолей. Кроме того, Т-хелперы 1 активируют Т-хелперы 2 и способствуют цитотоксической функции макрофагов.
Таблица 15
Субклассы Т-лимфоцитов, их функции и специфические рецепторы
Т-хелперы 2 секретируют IL-4, IL-5, IL-6 и вводят их в цитоплазму В-лимфоцитов путем прямого контакта своих рецепторов с определенными рецепторами на мембране В-лимфоцитов. Таким образом, Т-хелперы 2 вызывают пролиферацию и дифференциацию В-лимфоцитов в антителообразующие клетки и тормозят функцию Т-хелперов 1. Т-хелперы 1 и Т-хелперы 2 различаются по чувствительности к IL-2 и IL-4; кроме того, Т-хелперы 1 устойчивы к радиоактивному облучению, а Т-хелперы 2 – чувствительны.
Активированные Т-супрессоры играют важную роль в регуляции иммунитета. Они способны непосредственно контактировать с рецепторами Т-хелпера, а также синтезировать и секретировать свои медиаторы. Благодаря этому Т-супрессоры могут подавлять функции Т-хелперов. Последние обладают высокой чувствительностью к Т-супрессорам и их медиаторам. Взаимодействуя с мембраной Т-хелпера, Т-супрессоры угнетают секрецию ими интерлейкинов, синтез рецепторов к IL-1, IL-2 и другие иммунобиологические функции Т-хелперов, в результате чего подавляется пролиферация и дифференциация эффекторных Т-клеток, а также формирование антителообразующих клеток против данного антигена. Иначе говоря, Т-супрессоры подавляют временно или постоянно образование клонов иммунокомпетентных Т– и В-клеток против определенного антигена. Особо важную роль Т-супрессоры выполняют в эмбриональном периоде и в течение первой недели жизни новорожденного: они способствуют возникновению толерантности к собственным белкам и подавляют действие проникающих в организм новорожденного Т-цитотоксических лимфоцитов матери на аллоантигены самогmо развивающегося ребенка. Функция Т-супрессоров через неделю после рождения блокируется возникающей к этому времени особой популяцией Т-клеток – Т-контрсупрессорами, которые продуцируют собственный лимфокин. Этот лимфокин взаимодействует с Т-хелперами, они приобретают резистентность к Т-супрессорам и их лимфокинам. Т-контрсупрессоры отличаются от Т-супрессоров по своим специфическим рецепторам, в частности, они содержат отсутствующий у Т-супрессоров рецептор к лектину Vicia villosa. Этот рецептор способствует выполнению функций Т-контрсупрессорами.
Т-лимфоциты распознают чужеродные антигены, предсталяемые им молекулами МНС класса I или класса II, с помощью особого Т-клеточного рецептора (ТКР). В результате взаимодействия этих структур образуется тримолекулярный комплекс: молекула МНС + антиген + ТКР (см. рис. 63, в). Биосинтез ТКР, как и антител, кодируется комплексом генов: V, D, J, C, однако они отличаются от генов Ig, это самостоятельное семейство генов. Число возможных вариантов их объединения составляет 1016, что обеспечивает большое разнообразие антигенсвязывающих центров ТКР. Все субпопуляции Т-клеток используют ТКР непосредственно для распознавания антигена.
Структура ТКР во многом напоминает структуру молекулы антитела. ТКР состоит из двух цепей – α и β (редко встречается вариант γ/δ). Каждая цепь содержит V– и С-домены, обладающие высокой степенью гомологии с V– и С-доменами Ig. За С-доменами, около мембраны, располагается шарнирная область – место образования дисульфидной связи между цепями. Каждая цепь заякорена в мембране своим гидрофобным участком. Антигенсвязывающий центр ТКР формируется тремя гипервариабельными участками каждой цепи и имеет форму щели, структура которой соответствует пространственной структуре представляемого антигена.
Каждый Т-лимфоцит несет ТКР, распознающий свой антиген только в комплексе с молекулой МНС (рис. 68А). Последняя имеет два участка связывания: гистотоп для ТКР и дезотоп – для пептида. Связь с пептидом осуществляется с помощью вариабельных аминокислотных остатков, обращенных внутрь антигенсвязывающего центра молекулы МНС.
Передача сигнала к ядру Т-клетки происходит по пути, общему для различных сигналов (факторы роста, деления, дифференцировки, антигены и др.). Трансмембранные рецепторы, воспринимающие эти сигналы, обладают (или приобретают ее, например, в случае присоединения к ТКР антигена) протеинтирозинкиназной активностью и регулируют так называемые Ras-зависимые сигнальные пути, в которых ключевую роль играет продукт протоонкогена ras – белок Ras. От него расходятся многие пути. Те трансмембранные белки-рецепторы, которые лишены протеинкиназной активности, приобретают ее, образуя после активации лигандом прочный комплекс с цитоплазматическими протеинтирозинкиназами Src-семейства. Именно через эти протеинкиназы цитокины осуществляют регуляторные функции, стимулируя Ras-зависимые и Ras-независимые сигнальные пути. Конечное звено всех сигнальных путей, регулирующих рост, деление и дифференцировку клеток, – образование фактора транскрипции и связывание его со специфическим цис-регуляторным элементом гена-мишени. Факторами транскрипции служат фосфорилированные белки – продукты протоонкогенов c-myc, c-fos, c-jun, c-myb и генов-супрессоров клетки. Цис-регуляторные элементы – это сравнительно короткие олигонуклеотиды, богатые гуанином и цитозином. Они образуют вилки («палиндромы»). Взаимодействие их с промоторами генов активирует или подавляет транскрипцию. Сигнальные белки узнают партнеров с помощьью особых некаталитических доменов SH2 (100 – 150 аминокислотных остатков) или SH3 (50 – 80 аминокислотных остатков), названных так из-за расположения рядом с каталитическим доменом Src-киназы SH1. За короткое время SH2– и SH3-домены были обнаружены не только в цитоплазматических протеинтирозинкиназах, но и во многих других сигнальных белках (липидкиназах, фосфолипазах, факторах транскрипции и т. д.). Каждый SH2-домен узнает своего партнера по определенной последовательности 3 – 5 аминокислот, расположенных сразу же после фосфотирозина, а SH3-домен – по участкам, богатым пролином. Такая специфичность взаимодействия позволяет связать звенья одного сигнального пути в единую цепь и обеспечить безошибочное проведение сигнала к месту назначения. В некоторых сигнальных белках обнаружены домены с избирательным сродством к мембранным фосфолипидам. Они названы плекстриновыми доменами (PH). Мишенью для сигнала, воспринятого ТКР и переданного в ядро, служит ген, кодирующий синтез ИЛ-2. Активация этого гена приводит к синтезу и секреции ИЛ-2, который стимулирует пролиферацию и дифференцировку В-лимфоцитов.
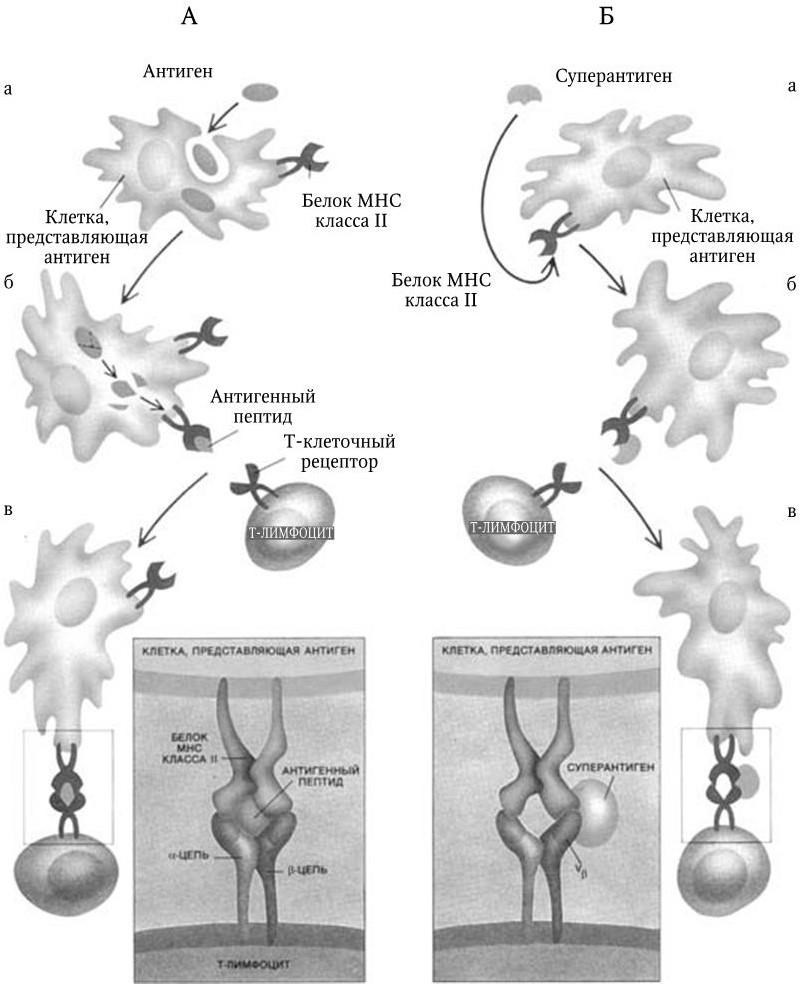
Рис. 68. Взаимодействие антигенов (А) и суперантигенов (В) с Т-лимфоцитами (по: Д. Бернс-Пайзеру, 1992):
Обычные антигены (слева) вначале поглощаются какой-либо из клеток, представляющих антигены (а), подвергаются в ней процессингу (б), и их антигенный пептид представляется в комплексе с молекулой МНС класса II Т-лимфоциту, который с помощью своих молекул МНС класса II распознает его и связывается с ним (в). Обе цепи рецептора (α и β) контактируют с антигеном (в рамке). Суперантигены (справа) прямо связываются с наружной стороной молекул МНС (а и б), а затем с Т-клеточным рецептором (в), присоединяясь к вариабельной (Vβ) части β-цепи (в рамке). Связывание с Vβ обусловливает мощное воздействие суперантигенов на Т-лимфоциты
Существует полная взаимосвязь между фенотипом Т-лимфоцитов (CD4/CD8) и молекулами МНС, которые определяют Т-клеточный ответ: все без исключения Т-киллеры (CD8+) несут молекулы МНС класса I, а все Т-хелперы (CD4+) – молекулы МНС класса II.
Предполагается, что гены Ir, открытые Бенасеррафом в 1967 г., каким-то образом определяют силу иммунного ответа на конкретный антиген через тримолекулярный комплекс.
Особенности взаимодействия суперантигенов с Т-лимфоцитами
Суперантигены отличаются от обычных антигенов очень большой силой иммунного ответа. Предполагается, что необычная сила ответа на суперантиген обусловлена по крайней мере двумя обстоятельствами: 1) суперантиген связывается с молекулой MHC без предварительного процессинга и 2) суперантиген присоединяется к наружному участку молекулы MHC. Поэтому он и распознается не внутренней поверхностью антигенсвязывающего центра ТКР (как обычный антиген), а той частью вариабельного участка b-цепи, которая находится с наружной стороны цепи (см. рис. 68Б). Обычный антиген распознается только Т-специфическим лимфоцитом, вызывает его пролиферацию и адекватный иммунный ответ (рис. 69). Суперантиген может распознаваться и связываться разными Т-лимфоцитами, у которых β-цепь ТКР содержит определенные типы вариабельных доменов (у человека таких типов около 30). Связываясь с ними, суперантиген оказывает сильное воздействие на Т-лимфоциты. Результат этого – суперпродукция ИЛ-2, отравляющего организм и вызывающего СТШ (рис. 70). Свойствами суперантигенов обладают некоторые бактериальные экзотоксины, а также различные компоненты самой бактериальной клетки. Наличие суперантигенов у бактерий влияет на клиническую картину вызываемых ими заболеваний.
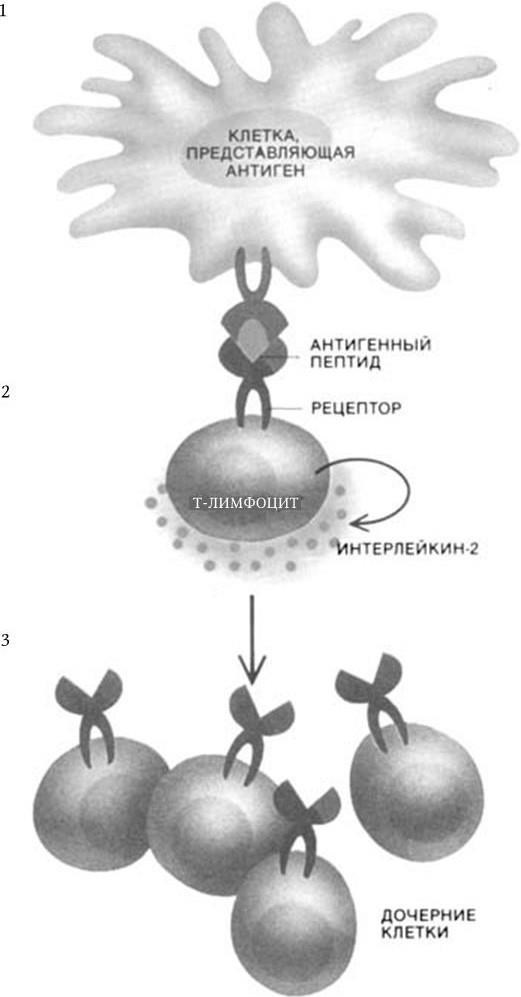
Рис. 69. Иммунный ответ на обычные антигены (по: Д. Бернс-Пайзер, 1992):
1. С данным антигеном связываются только те клетки, которые имеют специфичный к нему рецептор (≈ 0,01 % Т-хелперных лимфоцитов).
2. Связавшие антиген клетки секретируют интерлейкин-2.
3. Интерлейкин-2 вызывает пролиферацию антиген-специфических Т-лимфоцитов. Иммунная система распознает и разрушает зараженные любым патогеном клетки, не причиняя вреда нормальным клеткам
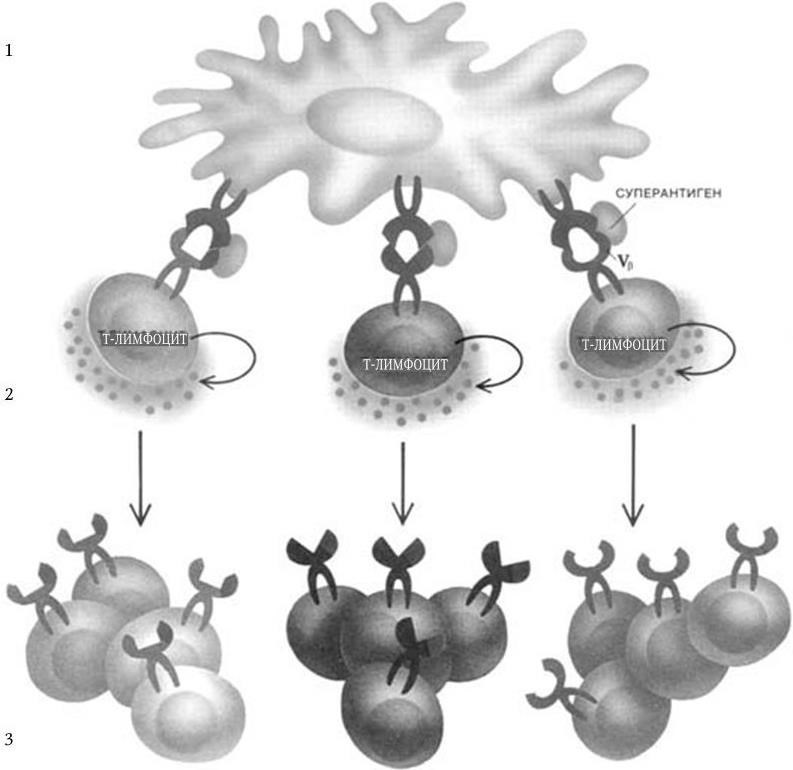
Рис. 70. Иммунный ответ на суперантигены (по: Д. Бернс-Пайзер, 1992):
1. С суперантигеном связывается около 20 % Т-лимфоцитов, несущих на своей поверхности в области Vβ рецепторы для различных антигенных пептидов.
2. Связавшие суперантигены Т-лимфоциты секретируют интерлейкин-2, что приводит к аномально высокой концентрации его.
3. Т-лимфоциты размножаются, образуется огромное множество таких клеток. Следствием суперпродукции интерлейкина-2 могут быть лихорадка и другие симптомы интоксикации, а образование избыточного количества антиген-специфических Т-лимфоцитов может стать причиной аутоиммунных реакций (активация и размножение немногочисленных Т-лимфоцитов, реагирующих на «свои» антигены, приводят к атаке нормальных тканей) или привести к подавлению иммунной системы, так как Т-лимфоциты, стимулированные суперантигеном, могут после активации погибнуть
Назад: Глава 33 Клеточные основы иммунитета. Органы иммунитета
Дальше: Глава 35 В-лимфоциты и их функции

