Книга: Онтогенез. От клетки до человека
Назад: Глава 15 Как набраться ума-разума
Дальше: Глава 17 Друзья и враги
Глава 16
Мера всех вещей
Будь твой дед хоть под потолок ростом, тебе расти самому.Ирландская пословица
В Венеции, в Галерее Академии, что на южном берегу Большого канала, хранится один из ярчайших символов Возрождения – рисунок «витрувианского человека» работы Леонардо да Винчи. На нем изображена обнаженная мужская фигура. И ноги и руки фигуры показаны в двух положениях: ноги вместе или немного расставлены, руки вытянуты в стороны параллельно земле или вытянуты и приподняты до уровня макушки. Тело вписано в окружность с центром в пупке, а также в квадрат, сторона которого соответствует росту человека (рис. 74).
Пояснения к рисунку написаны зеркальным письмом, которым Леонардо обычно делал тайные записи, и содержат сведения об относительных размерах частей тела. К ним относятся, в частности, следующие правила: длина разведенных в стороны рук равна росту человека (это показывает квадрат), расстояние от линии роста волос до подбородка равно одной десятой доле человеческого роста, расстояние от локтя до кончиков пальцев – четверти, длина уха составляет треть длины лица и так далее. Всего правил тринадцать. Придумал их не Леонардо, а римский архитектор Витрувий еще в первом веке до нашей эры, и поэтому рисунок Леонардо обычно называют «витрувианским человеком». Как показывает стилизованный рисунок «витрувианской женщины», выполненный современной художницей Сьюзан Доротеей Уайт, те же самые правила справедливы и для женского тела.
Хотя теперь мы понимаем, что эти правила отражают не абсолютные, а усредненные пропорции, и что у многих людей лица более вытянуты, а уши длиннее, чем следует из правил Витрувия, все же удивительно, с какой точностью большинство тел, за исключением аномальных, укладываются в эту схему. Каким же образом мельчайшие клетки могут так точно сложиться в определенные формы и пропорции тела, во много раз превышающего их собственные размеры?
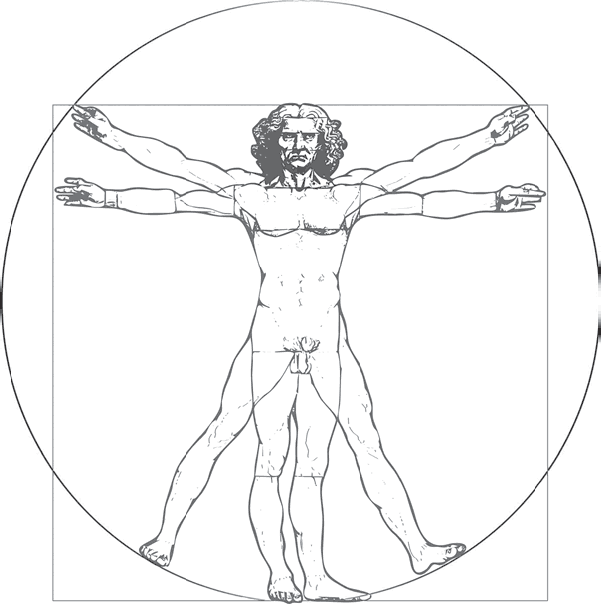
Рис. 74. Витрувианский человек (рисунок Леонардо да Винчи) (Wikimedia Commons) CC gaggio1980-Fotolia.com
Пропорции взрослого человека иные, чем у эмбриона: относительные размеры частей тела закономерно отличаются у плода, младенца, ребенка и подростка. Например, голова новорожденного младенца намного больше относительно общих размеров тела, а конечности намного короче, чем у взрослого человека, не говоря уже о том, что все тело гораздо меньше. На протяжении дальнейшего роста размер различных частей тела находится под контролем, так что они всегда пропорциональны по отношению друг к другу. Таким образом поддерживается замечательная, почти точная симметрия человеческого тела. При этом части тела не обязательно контактируют друг с другом (например, левая и правая нога), а длина таких крупных частей тела, как конечности, даже у новорожденного превышает длину отдельной клетки в десять тысяч раз. Как же части тела измеряют сами себя? Какие механизмы сообщают растущему организму чувство меры? Если в двух словах, то это пока неизвестно, но, собрав воедино данные экспериментов на разных животных, от плодовых мушек до млекопитающих, мы можем высказать ряд обоснованных предположений.
Прежде чем говорить о пропорциях, давайте посмотрим, как контролируется абсолютный размер человеческого тела. Существенная часть наших знаний на эту тему получена при изучении людей, у которых этот контроль так или иначе нарушен, а именно карликов и гигантов.
Уже давно было замечено, что гигантизм, при котором рост человека намного больше среднего, часто связан с опухолями гипофиза. Если такие опухоли активны в период нормального роста ребенка, он вырастает очень высоким (2,10–3,60 м). При этом такие люди, несмотря на исключительно высокий рост, имеют нормальные пропорции тела.
Гипофиз – сложный орган, выделяющий множество гормонов, но с точки зрения контроля размера тела важнее всего так называемый гормон роста. Здоровый гипофиз выделяет небольшие количества этого гормона в пульсирующем ритме, один выброс в несколько часов. Как правило, наибольшая активность гормона достигается во время сна. Концентрация гормона роста довольно высока в раннем детстве, когда организм растет быстро, и резко снижается до скромного «взрослого» уровня в возрасте примерно от 18 до 20 лет. Люди, в организме которых производится мало гормона роста, очень невысокие (около 1,2 м), но имеют более или менее нормальные, «витрувианские» пропорции тела. Сама по себе связь между количеством гормона роста и размером тела еще не говорит о том, что здесь причина, а что – следствие. Однако тот факт, что дети с низким содержанием гормона роста растут вполне нормально, если делать им инъекции этого гормона,,, явно указывает на то, что количество гормона роста определяет размер тела.
Гормон роста не оказывает непосредственного влияния на рост и размножение клеток. Он заставляет некоторые клетки, в частности клетки печени, образовать вторую сигнальную молекулу дальнего действия – инсулиноподобный фактор роста I (ИФР-I; родственная молекула ИФР-II контролирует рост плода). Именно ИФР-I передает большинству клеток организма инструкции, касающиеся размера состоящих из них структур. Гормон роста регулирует синтез ИФР-I за счет сложного процесса, включающего его связывание со специфическим рецептором и запуск внутреннего сигнального пути, приводящего к экспрессии гена, кодирующего ИФР-I. Мутация гена, кодирующего этот рецептор, приводит к тому, что клетки носителей мутации приобретают аномальную чувствительность к гормону роста. Это является причиной особого типа карликовости – синдрома Ларона. Для людей с этим синдромом характерен низкий рост и непропорционально малый размер таких внутренних органов, как сердце, в сочетании с нормальными пропорциями скелета. (Любопытно, что среди людей с синдромом Ларона много долгожителей. Самые разные животные, от круглых червей до грызунов, в организме которых уровень ИФР-I опускается ниже нормы, как правило, переживают нормальных животных своего вида, и, возможно, у людей имеет место тот же самый эффект.)
Далеко не при всех нарушениях контроля роста пропорции тела остаются нормальными. Яркий пример этого – Анри де Тулуз-Лотрек, художник, живший через четыре столетия после того, как Леонардо да Винчи нарисовал «витрувианского человека». Тулуз-Лотрек – пост импрессионист, запечатлевший на своих картинах, рисунках и плакатах богемный декаданс Парижа на рубеже веков, – обладал необычным телосложением. Его лицо и туловище были нормальными по размеру и пропорциональными относительно друг друга; голова, правда, была деформирована из-за позднего и неполного закрытия зазоров между костями черепа. В возрасте 13 лет у художника было нормальное телосложение, но затем ноги перестали расти, а туловище – нет. Поэтому в зрелом возрасте ноги выглядели короткими, и рост его составлял всего 1,5 м. Кости были хрупкими, и ноги часто болели. Большинство современных медицинских генетиков, рассматривавших случай Тулуз-Лотрека, сходятся на том, что он страдал от генетического нарушения, которое теперь называется пикнодизостоз.,,
Пикнодизостоз – заболевание крайне редкое. За всю историю медицины описано только около двухсот случаев. Он обусловлен мутацией в гене, кодирующем фермент, который должен в том числе высвобождать ИФР-I из костей.,,, Если такого фермента нет, ИФР-I остается в «ловушке» и не может стимулировать рост. Пациентам с пикнодизостозом хорошо помогают дополнительные дозы гормона роста, поддерживающие уровень ИФР-I в нужных пределах. Гораздо чаще, чем пикнодизостоз, встречается другой тип карликовости – ахондроплазия, затрагивающая примерно одного из 25 тысяч людей. Ахондроплазия вызвана мутацией в сигнальном рецепторе, которая нарушает нормальный рост костей конечностей.,, Люди с ахондроплазией имеют непропорционально короткие конечности, а также определенные неправильности их формы и формы других частей тела.
Телосложение Тулуз-Лотрека и людей с ахондроплазией свидетельствует о том, что неспособность некоторых частей тела поспевать за общим ростом не обязательно останавливает рост всего организма. Таким образом, пропорциональность не является следствием того, что каждая часть тела постоянно «проверяет», не переросла ли она другие части. Разные части тела должны по-своему реагировать на гормон роста, ИФР-I и другие влияющие на рост гормоны. Это справедливо даже для двух однотипных частей тела. В одном особенно наглядном эксперименте, проведенном двадцать лет назад, рост одной задней лапы кролика ингибировался с помощью местного введения химического препарата. Вторая задняя лапа кролика продолжала расти нормально, и кролик вырос кривобоким. Очевидно, что одинаковая длина ног не обусловлена тем, что растущие конечности обмениваются сигналами и подстраиваются друг к другу в процессе удлинения.,
Из строения конечностей Тулуз-Лотрека и людей с ахондроплазией следует еще один важный вывод. В обоих случаях определенное биохимическое нарушение, являющееся первопричиной отклонения в развитии, непосредственно влияет на рост длинных костей конечностей. Прямого влияния на рост кожи, мышц, нервов, кровеносных сосудов и т. д. нет, но тем не менее эти ткани не образуют вокруг короткой кости бесформенную мягкую массу, предназначенную для нормальной конечности. Они растут в соответствии с длиной аномальной конечности. Это хорошо иллюстрирует глубинную дихотомию контроля над размерами тела: некоторые ткани организма, например кости кролика в эксперименте, самостоятельно регулируют свой абсолютный размер и могут считаться главными детерминантами размеров тела, а другие ткани ориентируются не на свои абсолютные размеры, а на размеры относительно тканей первого типа. В плане контроля над размерами ткани второго типа оказываются подчиненными, их задача заключается в том, чтобы не отставать от роста тканей-начальников, но никогда не обгонять его. Поэтому проблема контроля над размерами распадается на два вопроса: во-первых, как ткани-начальники измеряют сами себя, и, во-вторых, как подчиненные ткани подстраиваются под начальников?
Давайте сначала посмотрим, как ткани-начальники, скажем кости, контролируют свой рост. Возьмем в качестве примера развитие конечностей. Кости конечностей растут не по всей длине, а в специализированной зоне роста – эпифизарной пластинке. Она располагается близко к концу, но не на самом конце кости. Эпифизарная пластинка подразделяется на несколько зон (рис. 75). На внешнем ее конце клетки активно размножаются. Эта пролиферация является одной из непосредственных причин роста кости, хотя и не самой главной. На внутренней границе зоны пролиферации клетки меняют свое поведение и начинают формировать хрящ – мягкий предшественник кости. За счет этого внутренняя граница зоны пролиферации смещается на один ряд клеток, так что в приграничном положении оказываются новые клетки. Им нужно некоторое время на то, чтобы отреагировать на изменение ситуации, после чего эти клетки тоже переключатся на образование хряща, и так далее. Таким образом, на дистальном конце зоны пролиферации постоянно появляются новые клетки, а на проксимальном конце клетки постоянно покидают эту зону и создают хрящ. В результате зона пролиферации продвигается вперед.
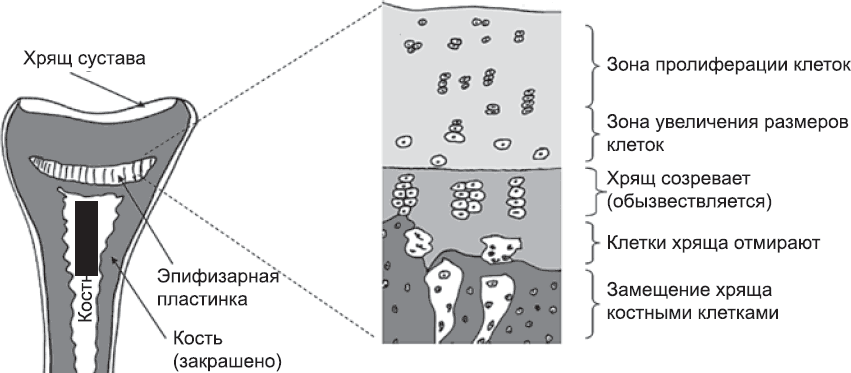
Рис. 75. Эпифизарная пластинка кости растущей конечности
Формирование хряща сопровождается увеличением объема: отчасти потому, что клетки увеличиваются в размерах, а отчасти потому, что они выделяют студенистое вещество, заполняющее межклеточное пространство. Это увеличение объема ткани и является основной причиной удлинения кости. Со временем хрящ созревает, хрящевые клетки отмирают и замещаются клетками, источником которых является прилегающая сформировавшаяся кость. Эти клетки заселяют хрящ, постепенно превращая его в костную ткань. Поскольку клетки в зоне пролиферации продолжают активно размножаться, процесс идет непрерывно: кость удлиняется, а зоны эпифизарной пластинки смещаются все дальше в дистальном направлении. Таким образом, скорость удлинения кости зависит прежде всего от скорости пролиферации клеток в наружной части эпифизарной пластинки и скорости, с которой клетки покидают зону пролиферации и начинают формировать хрящ.
Насколько мы знаем, скорость пролиферации и скорость перехода к созданию хряща зависят от сигналов двух типов: внутренних сигналов, которые организуют эпифизарную пластинку, и внешних сигналов, которые задают темп ее работы. Внутренние организующие сигналы необходимы для поддержания достаточного уровня пролиферации, то есть такого, при котором эпифизарная пластинка сохраняется в более или менее неизменном виде, несмотря на постоянную «потерю» клеток в каждой зоне («потеря» связана с тем, что клетки по мере созревания переходят в следующую зону). Зрелые хрящевые клетки, обреченные на гибель и замену костными клетками, выделяют сигнальный белок, под действием которого клетки на внутренней границе зоны пролиферации меняют поведение и начинают формировать хрящ. Тот факт, что сигнал подают сами зрелые клетки, автоматически приводит к равновесию между количеством полностью созревших, готовых погибнуть хрящевых клеток и количеством клеток в зоне пролиферации, которые получают сигнал к превращению в новые хрящевые клетки. Такое равновесие позволяет поддерживать размер зоны формирования хряща по мере ее продвижения в дистальном направлении.
Только что описанная система сигналов чревата одной опасностью: если под действием этих сигналов клетки начнут переходить к формированию хряща слишком быстро, пролиферирующие клетки не будут поспевать за ними, и популяция клеток зоны пролиферации может истощиться. Предотвратить такую ситуацию помогает другой сигнальный белок. Его выделяют клетки, которые только что перестали делиться и перешли к созданию хряща. Этот сигнал распространяется через развивающуюся кость. На него реагируют особые клетки на внешней стороне кости, за эпифизарной пластинкой. Они начинают вырабатывать еще один сигнальный белок. Он распространяется обратно к зоне пролиферации на эпифизарной пластинке и заставляет расположенные там клетки размножаться быстрее (рис. 76).
В результате совместного действия этих двух сигнальных систем одна из которых говорит «созревайте!», а другая «размножайтесь!», к созреванию приступает именно столько клеток, сколько нужно для замены погибающих, а число клеток, вступивших на путь созревания, точно компенсируется за счет пролиферации. Таким образом, система в целом остается стабильной.
Основным внешним регулятором роста костей является уже рассмотренная нами система «гормон роста – ИФР-I». Далее в этой главе я буду для простоты называть ее просто «гормоном роста». По-видимому, гормон роста меняет уровень пролиферации и, таким образом, действует сообща с внутренними сигналами эпифизарной пластинки, регулирует количество клеток, которое можно потратить на формирование хряща. Влияния гормона роста недостаточно, чтобы отменить способность эпифизарной пластинки к самоорганизации, поэтому ее структура остается постоянной, независимо от того, медленно или быстро растет человек. Предположительно, эпифизарные пластинки костей различных типов (бедренной кости, костей пальцев и т. д.) обладают разной чувствительностью к гормону роста. Поэтому они растут с разной скоростью, и образуются кости с характерными относительными пропорциями. Заметим, что какая бы система не отвечала за этот процесс, она должна легко перестраиваться, учитывая большое разнообразие пропорций костей у разных обезьян и человека.
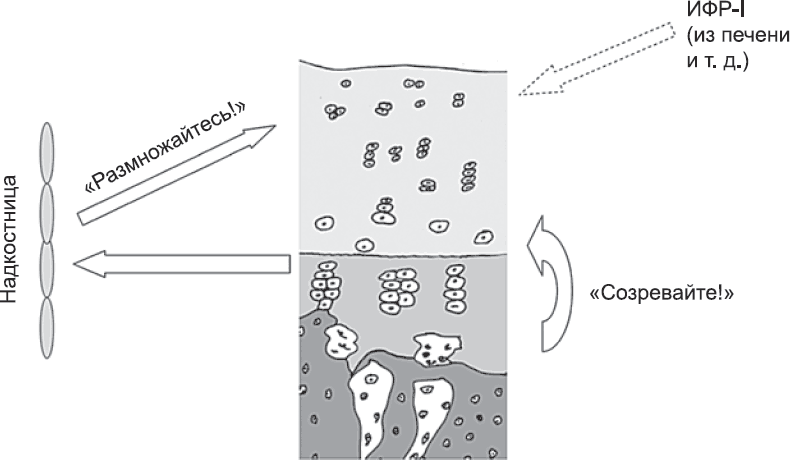
Рис. 76. По мере роста эпифизарной пластинки ее структура остается постоянной за счет различных сигналов. Некоторые из этих сигналов внутренние, а некоторые «ретранслируются» через промежуточную станцию в надкостнице
Как же все-таки получается, что кости одного типа, например бедренные кости левой и правой ноги, в конечном итоге примерно равны по размеру? Ответ подсказывает нам вариант описанного выше эксперимента с лапой кролика. В этом варианте, опубликованном в той же статье, что и основной эксперимент, исследователи ограничивали рост одной конечности так, чтобы кролик стал асимметричным, а затем снимали это ограничение. Результат был поразительным – короткая лапа тут же начинала расти быстрее, чем росла нормальная, и в конечном итоге достигала нормальной длины. Понятно, что в организме было достаточно гормона роста, чтобы поддерживать быстрый рост, но нормальная конечность не реагировала на него столь активно, как та, с которой сняли временное ограничение на рост. Почему же две конечности с разной интенсивностью реагировали на одну и ту же концентрацию циркулирующего гормона роста? Может быть, процесс роста сам по себе снижал чувствительность эпифизарной пластинки к подобным гормонам? Такой гипотетический механизм был бы изящным способом обеспечить равную длину парных конечностей: даже если бы по какой-то причине одна из них отстала в росте, она бы сохранила повышенную чувствительность к гормону роста и имела бы шанс «догнать» нормальную конечность.
Особенности одного из внутренних сигнальных путей наводят на мысль о возможном механизме того, как скорость роста кости может зависеть от уже достигнутого размера. Как я уже говорил, созревающие клетки влияют на увеличение числа клеток в зоне пролиферации не напрямую, а через сигнальную петлю, ведущую к внешнему слою растущей кости и обратно к зоне пролиферации. Если кость небольшая, внешний слой находится не слишком далеко даже от середины эпифизарной пластинки, и сигнальная петля получается короткой и эффективной. Когда кость увеличивается, края эпифизарной пластинки находятся еще довольно близко к наружному слою кости и получают эффективный сигнал, а вот середина лежит дальше и получает менее интенсивный сигнал. В среднем по эпифизарной пластинке темпы пролиферации падают, и спад продолжается по мере роста кости и, следовательно, удлинения сигнальной петли. Таким образом, чем длиннее кость, тем менее охотно она откликается на одно и то же количество гормона роста. Такой механизм полностью объяснял бы и обычную симметрию левой и правой конечности, и результаты эксперимента с кроликом. Тем не менее хочу подчеркнуть, что этот механизм является гипотетическим, и его существование пока не доказано. Содержание этого абзаца – чисто умозрительная попытка непротиворечиво объединить данные разных исследований.
Скорость нашего роста непостоянна. С наступлением полового созревания происходит скачок роста, который характерен только для человека и, по-видимому, появился в эволюции у Homo erectus. После этого скачка наш скелет перестает расти, хотя тело может увеличиваться в обхвате за счет мышц или жира. Как скачок роста, так и его остановка, судя по всему, запускаются половыми гормонами, которые вызывают и такие явные признаки полового созревания, как рост волос на теле у мужчин и формирование молочных желез у женщин. Из всех половых гормонов для роста важнее всего эстроген. Его часто называют женским половым гормоном из-за его роли в менструальном цикле, но он вырабатывается и в организме мужчин (под действием фермента, превращающего тестостерон в эстроген). Эстроген стимулирует производство гормона роста и ускоряет рост, а также непосредственно влияет на поведение костных клеток. В этом возрасте рост может даже немного опережать скорость отложения минералов, необходимых для окончательного формирования кости. Поэтому кости подростков могут быть очень хрупкими. Примерно половина детей ломают кости в подростковом возрасте, и половина этих переломов приходится на быстро растущие длинные плечевые кости. Следует заметить, что хрупкость костей – не единственная причина повышенного риска переломов в этом возрасте. Растущая физическая сила нередко сочетается у подростков с полным отсутствием головы на плечах.
Расплачиваться за обусловленный эстрогеном быстрый рост приходится эпифизарной пластинке. Под действием высоких концентраций эстрогена клетки перестают размножаться и переходят к образованию хряща. Этот эффект настолько силен, что, по-видимому, нарушает тонкое равновесие самоорганизующихся петель обратной связи, которые поддерживают организацию эпифизарной пластинки. Это означает, что к самому концу полового созревания, когда уровень эстрогена очень высок у обоих полов, пролиферация отстает от созревания. Постепенно вся эпифизарная пластинка превращается в зрелый хрящ. После этого она «закрывается» и утрачивает способность расти. Есть мутации, при которых эстроген не производится и, следовательно, эпифизарные пластинки не закрываются. Носители таких мутаций, повзрослев, продолжают расти. Такой рост можно остановить при помощи инъекций эстрогена, который закрывает эпифизарные пластинки. Были случаи, когда эстроген вводили вполне нормальным девушкам, если было понятно, что они рискуют вырасти слишком высокими. Аналогичным образом, невысоким мальчикам-подросткам вводили препараты, блокирующие действие эстрогена, чтобы эпифизарные пластинки не закрывались еще некоторое время и мальчики успели подрасти. Эта книга – не место для обсуждения вопросов об этичности такого вмешательства и о том, кто имеет право диктовать «правильный» рост конкретного человека. Я привел эти примеры для того, чтобы показать два важных момента: во-первых, эстроген закрывает эпифизарные пластинки, во-вторых, изменив время их закрытия, можно повлиять на окончательный рост. Таким образом, окончательный рост человека определяется, помимо скорости роста, еще и временны́м фактором.
Подведем промежуточный итог. Насколько нам известно, контроль над размером скелета осуществляется следующим образом.
I. Кости растут благодаря самоорганизации пролиферирующих и созревающих клеток, которые обмениваются сигналами.
II. Гормон роста стимулирует рост, но растущие кости постепенно теряют чувствительность к нему.
III. Половые гормоны вызывают быстрый рост в период полового созревания, но при сильном повышении их концентрации нарушается самоорганизация эпифизарных пластинок; они закрываются, и рост прекращается. Гормон роста производится непосредственно в гипофизе, в то время как половые гормоны производятся в половых железах в ответ в основном на команды из гипофиза. В этом смысле, если скелет является тканью-«начальником» процесса роста, то гипофиз осуществляет общее руководство процессом.
А что же происходит с тканями-«подчиненными» – с теми, которые подгоняют свой размер под общий размер тела, определяемый скелетом? Поразительно, но и у нормальных животных, и у мутантов со всевозможными отклонениями от нормального размера скелета и нормальных пропорций «подчиненные» ткани растут в соответствии с размерами тела, которое должны обслуживать. Это касается даже тех случаев, когда из-за аномальной формы тела эти ткани должны иметь необычные пропорции, например кожа на слишком короткой конечности нормальной ширины или кожа на животе тучного человека. Учитывая такую устойчивость к отклонениям, маловероятно, что «подчиненные» ткани слепо следуют биохимическим сигналам, сообщающим о количестве ткани-«начальника», ведь эти сигналы не могут сообщить тканям о необходимой форме, особенно в случае аномального строения. Тем не менее один «сигнал», способный эффективно донести информацию и об относительном размере, и о форме, все-таки есть. Этим «сигналом» является механическая сила.
Если ткань, например кожа ноги, не успевает за ростом нижележащих тканей, она будет растягиваться и испытывать натяжение. Натяжение – это сила, которая действует на весь объект, и 10 %-ное натяжение ткани можно будет обнаружить в любой точке этой ткани независимо от ее размера. Механизм, использующий чрезмерное натяжение как индикатор недостаточного роста, имеет то достоинство, что он не зависит от размера: он будет работать одинаково хорошо как на большом, так и на маленьком участке ткани. Другим достоинством этого механизма является то, что он никак не связан с конкретной формой. Если при избыточном натяжении клетки будут делится и образовывать дочерние клетки, формирующие дополнительную ткань в направлении натяжения, ткань всегда будет расти в нужном направлении. Знать заранее, куда расти, ей совершенно не нужно. Поэтому система будет надежно работать для разных форм тела. Это упрощает не только обслуживание самых необычных вариантов роста, но и эволюционные изменения формы тела.
Существуют убедительные доказательства того, что механическое натяжение может запускать пролиферацию клеток. Если в течение нескольких дней мягко оттягивать ухо живой крысы, темп пролиферации клеток уха увеличится, и ткань вырастет. Судя по оттянутым мочкам ушей у людей, которые постоянно носят тяжелые серьги, это явление характерно и для человека. Активизация пролиферации при растяжении может быть показана в простых экспериментах на культурах клеток, где одни клетки подвергаются натяжению в большей степени, чем другие., Клетки, используемые в этих экспериментах, в организме формируют клеточные пласты и трубки и соединяются друг с другом благодаря межклеточным контактам. Эти контакты механически связаны друг с другом внутри каждой клетки при помощи белковых микрофиламентов, которые проходят через клетку насквозь и всегда создают легкое натяжение. Клетки культивируются на поверхности, где «островки», к которым клетки могут легко прикрепиться, перемежаются участками, к которым они прикрепляться не могут. Островки в форме квадратов или звезд имеют прямые края и острые углы. Клетки, расположенные вдоль прямых краев, не испытывают сильного механического воздействия с их стороны; на них действует только легкое натяжение, которое испытывают и создают все клетки. А вот клеткам, расположенным в острых углах, приходится изгибаться самым невероятным образом и по-прежнему противостоять легкому натяжению со стороны соседей. Это означает, что они испытывают гораздо более сильное напряжение. Именно такие угловые клетки размножаются наиболее активно (рис. 77). Если растянуть поверхность, на которой растут такие клетки, имитировав таким образом растяжение ткани, не поспевающей за ростом тела, пролиферация будет происходить по всей площади поверхности.
Если напряжение и натяжение – это способ сообщить ткани, что она растет недостаточно быстро, то сжатие и скученность можно использовать, чтобы подать клеткам противоположный сигнал – их теперь достаточно, нужно перестать делиться. Уже давно было показано, что, если поместить нормальные клетки в чашку Петри, они размножаются, пока не покроют дно, после чего деление прекращается. Если удалить некоторые клетки, например проведя по дну чашку Петри стерильным ластиком, то клетки на границе образовавшегося просвета будут делиться снова, пока не заполнят его, а потом опять остановятся. Это явление, получившее название «контактное торможение», послужило одним из первых указаний на то, что пролиферация клеток может контролироваться их собственным ощущением скученности. Расшифровать механизмы этого явления не удавалось несколько десятков лет, но последние данные, полученные на плодовых мушках, проливают свет на эту проблему. Ключом к механизму, насколько мы теперь понимаем, являются две крупные молекулы белков клеточной поверхности, похожие по строению на адгезионные молекулы, которые сцепляют клетки между собой. Есть очень убедительные данные о том, что эти молекулы тоже «прилипают» друг к другу. Чем больше клеток, тем больше площадь соприкосновения их поверхностей и тем сильнее взаимодействие между поверхностными белками. Это взаимодействие запускает сложный сигнальный путь внутри клетки, который приводит к ингибированию пролиферации.,
Ткани-«подчиненные» могут почувствовать свою соразмерность еще одним способом – отследив свое собственное биохимическое воздействие на организм. Они могут сделать это напрямую или опосредованно за счет «переговоров» с другими типами тканей. О такой регуляции уже шла речь в связи с кровоснабжением (глава 9): рост новых кровеносных сосудов отчасти контролируется тем, насколько успешно имеющиеся сосуды обеспечивают ткани кислородом. Указания на то, что подобный метод могут использовать целые органы, были получены в экспериментах по трансплантации, в которых животному сначала удаляли какой-то орган (не из числа жизненно важных), а затем пересаживали один или несколько эмбриональных зачатков того же органа. Эти эксперименты обычно ставятся для изучения трансплантации, так что полученные сведения о контроле над размером оказались своего рода бонусом. Если пересадить животному с удаленной селезенкой один зачаток селезенки плода, он вырастет до размеров нормальной селезенки. Если пересадить животному несколько зачатков селезенки, то они перестанут расти, когда совокупность их индивидуальных объемов достигнет объема нормальной селезенки. Это говорит о том, что либо селезенки сами «поняли», что в организме достаточно селезеночной ткани, либо это обнаружил организм и подал сигнал маленьким селезенкам. Тем не менее это явление не носит универсального характера. Когда подобный эксперимент проводился над тимусом, каждый зачаток вырос до размеров нормального тимуса, и у животного возник избыток этих желез. Таким образом, разные органы регулируют свои размеры, руководствуясь разными правилами. Это значительно усложняет проблему.

Рис. 77. Когда клетки выращиваются на маленьких квадратных «островках», клетки по краям и особенно по углам испытывают сильное механическое напряжение. Уровень пролиферации у таких клеток выше, чем у клеток посередине «островка», которые не испытывают столь сильного напряжения (рисунок основан на оригинальной микрофотографии, сделанной Селестой Нельсон и ее коллегами)
Контроль над размером и пропорциями остается малоизученной областью. В этой главе я представил выжимку того, что нам уже известно. Размеры тела в основном диктует скелет, а большинство других тканей подстраивается под него. Рост скелета регулируется гормонами роста, которые производит гипофиз, и половыми гормонами. Они действуют на самоорганизующиеся эпифизарные пластинки в развивающихся костях. Другие ткани реагируют на механическое напряжение, обусловленное ростом скелета и других тканей, и растут в соответствии с ним. Некоторые внутренние органы могут «ощущать» собственную соразмерность, отслеживая свое биохимическое воздействие на организм, но это относится не ко всем органам. Ни один из этих механизмов не предполагает, что клетки должны иметь представление об организации своей ткани. Ни чертежи, ни подробные планы действий клеткам не нужны. Все, что от них требуется, – это следовать простым командам, таким как «размножайтесь быстрее, если получаете вот такой сигнал». Размер, пропорции и симметрия тела – результат следования этим простым, слепым, локальным правилам. С этой точки зрения, организм подростка, в котором больше клеток, чем звезд в нашей галактике, использует абсолютно те же самые основные принципы контроля над развитием, что и организм крошечного эмбриона, каким он когда-то был.
Назад: Глава 15 Как набраться ума-разума
Дальше: Глава 17 Друзья и враги

