Книга: Онтогенез. От клетки до человека
Назад: Глава 11 Не покладая рук (и ног)
Дальше: Глава 13 В паутине проводов
Глава 12
X или Y? Вот в чем вопрос
Продолжение человеческого рода – великое чудо и таинство. Если бы Господь спросил моего мнения по этому вопросу, я посоветовал бы ему и дальше вылеплять новых людей из глины.Мартин Лютер
В предыдущих главах речь шла о том, что пережил каждый из нас без исключения, о том, что происходило с эмбрионом до возникновения памяти, индивидуальности и даже половых различий. В течение первых семи недель развития определить пол будущего ребенка по каким бы то ни было внешним признакам невозможно, и только хромосомный анализ может приоткрыть завесу этой тайны. Мужские и женские тела формируются из одного и того же материала; возникновение частей тела, характерных для одного из двух полов, не связано с усилением каких-то изначальных анатомических различий, поскольку таких различий нет. Это процесс, в ходе которого некоторые ткани эмбриона выбирают один из двух возможных путей развития – мужской или женский. У млекопитающих первыми выбирают этот путь половые железы (гонады), которые затем «сообщают» о своем решении другим частям организма.
Яичники женщин и семенники (яички) мужчин состоят из большого количества различных типов клеток, которые для простоты можно разделить на две группы. Во-первых, это клетки зародышевой линии – яйцеклетки или сперматозоиды и клетки, которые дают им начало. Только клетки зародышевой линии могут передать генетический материал новому человеку, поэтому в некотором смысле в них заключается суть репродуктивной системы. А если, как это делают некоторые биологи, считать, что жизнь – это передача генов из поколения в поколение, то в этих клетках заключается суть человеческой жизни. Как заметил Сэмюэль Батлер, «курица – это приспособление, с помощью которого одно яйцо производит другое». Вторая группа клеток половых желез известна под общим названием соматических клеток. Это название связано с тем, что эти клетки относятся к уже существующему телу («соме»), и ни они, ни их потомки не могут дать начало организму нового поколения. К соматическим клеткам относятся клетки, продуцирующие половые гормоны, клетки, обеспечивающие защиту и поддержку клеток зародышевой линии в процессе развития в сперматозоиды и яйцеклетки, а также многие другие клетки, которые поддерживают целостность гонад, обеспечивают их кровоснабжение и иннервацию, а также выполняют множество других рутинных функций.
Соматические клетки гонад и клетки зародышевой линии возникают в разных частях эмбриона. Соматические клетки гонад образуются по обе стороны от средней линии в верхней части туловища – примерно на уровне нижних долей легких, если мы представим в виде эмбриона самих себя. Такое положение гонад у эмбриона, резко контрастирующее с их локализацией у взрослых людей, особенно у мужчин, может показаться странным, но в свете эволюции оно вполне логично. Люди, как и другие млекопитающие, рептилии, птицы и земноводные, произошли от рыб. Даже у взрослых рыб, особенно у «примитивных» видов, гонады расположены в передней части тела. Там они могут взаимодействовать с формирующимися кровеносными сосудами и другими тканями, что жизненно важно для создания функциональной репродуктивной системы. Эти взаимодействия необходимы и в человеческом организме, а поскольку ткани, вовлеченные в процесс формирования гонад, участвуют и в формировании аорты (глава 9), они не могут сместиться без серьезнейших последствий для всей кровеносной системы. Поэтому и место формирования половых желез никуда передвинуться не может.
Клетки зародышевой линии (первичные половые клетки) впервые выявляются перед самым началом гаструляции, когда в эпибласте на заднем конце первичной полоски обосабливается группа из примерно пятидесяти клеток (глава 4). Вероятно, эти клетки вовлекаются в гаструляцию на очень ранних этапах, а затем они движутся вместе с мезодермой, которая выходит за пределы тела эмбриона., Пока в процессе гаструляции образуются туловище и голова (см. главу 4), клетки зародышевой линии находятся в верхней части желточного мешка (рис. 56). Там они и остаются, пока продолжается формирование тела эмбриона и его частей и структур: нервной трубки, сомитов, кровеносной системы и т. д. Когда основной план строения уже намечен, зародышевые клетки возвращаются обратно, в тело эмбриона, как за счет морфогенетических движений, связанных с формированием кишечника, так и за счет активной миграции. Внешняя поверхность развивающейся кишечной трубки служит им столбовой дорогой, по которой легко добраться от задней части тела до области развивающихся гонад, которые секретируют особый набор сигнальных молекул. Привлеченные этими молекулами, зародышевые клетки заползают в гонады. По ходу дела зародышевые клетки пролиферируют, и вместо пятидесяти пустившихся в путь клеток к гонадам прибывает пятитысячная армия (впоследствии клеток зародышевой линии станет еще больше).
Хотя именно клетки зародышевой линии дадут начало сперматозоидам и яйцеклеткам, пол эмбриона определяют не они, а соматические клетки половых желез. Процесс «принятия решения» о выборе пола основан на взаимодействии нескольких белков. Их названия неудобочитаемы, но необходимы для описания механизма определения пола, так что заранее прошу прощения.
На этом этапе развития конкретная группа соматических клеток половой железы начинает производить белок WT1. Он выполняет множество разных функций в клетках, но к самым важным относится связывание определенных последовательностей ДНК и включение вместе с другими ДНК-связывающими белками определенных генов. Большинство активируемых WT1 генов находятся на хромосомах, которые есть у каждого человеческого эмбриона, но один из них – ген SRY – находится на Y-хромосоме, которая есть только примерно у половины эмбрионов (почему это так, я объясню чуть позже). Если у данного эмбриона Y-хромосома есть, WT1 запустит образование белка SRY. Если же она отсутствует, то не будет и белка SRY. Это различие определяет весь дальнейший ход событий.
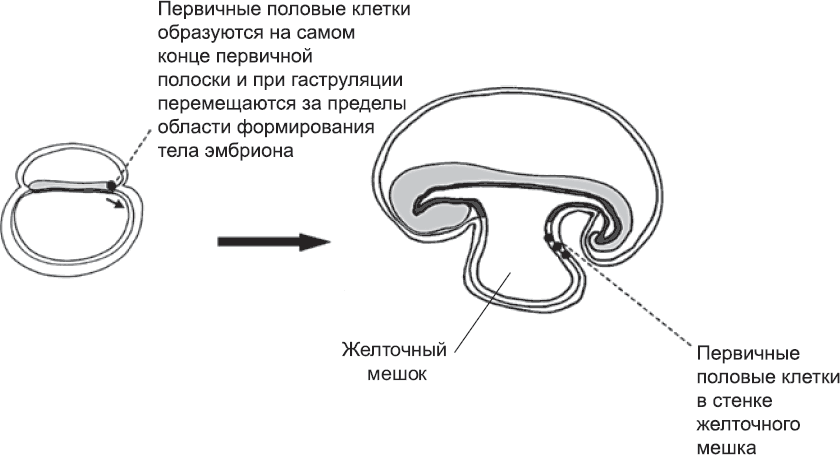
Рис. 56. Ранние клетки зародышевой линии (первичные половые клетки) образуются в задней части первичной полоски и перемещаются в желточный мешок, где ждут, пока идет процесс формирования тела эмбриона
Давайте сначала рассмотрим эмбрион, в котором может образоваться белок SRY. Он, как и WT1, связывается с определенными участками ДНК, но с другими, чем WT1. SRY активирует новые гены, которые, не будь этого белка, остались бы неактивными. Один из этих генов кодирует белок SOX9, а он в свою очередь активирует экспрессию очередной «порции» генов. Таким образом, различия в характере экспрессии генов между клеткой с Y-хромосомой и клеткой без нее растут как снежный ком. Отметим, что ген SOX9 находится не на Y-хромосоме, а на семнадцатой хромосоме, которая есть у каждого эмбриона.
Прежде всего белок SOX9 обеспечивает дальнейшее производство самого себя. Это важно, так как эмбрион должен принять четкое и бесповоротное решение по поводу своего пола, в противном случае может образоваться организм, сочетающий мужские и женские черты. Экспрессия SOX9 активирует сигнальный путь, основанный на белках FGF, а этот путь, в свою очередь, обеспечивает дальнейший синтез SOX9 даже в отсутствие SRY. Таким образом, петля SOX9 – FGF может поддерживать сама себя, и как только она активирована, Рубикон перейден: эмбрион встал на мужской путь развития (рис. 57).
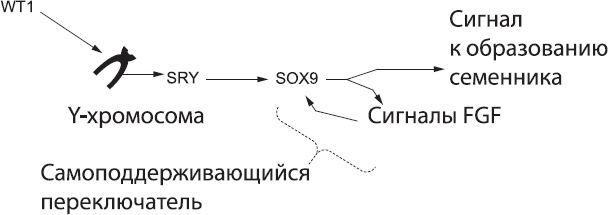
Рис. 57. Биохимический каскад «самцовости», который активирует переключатель SOX9 – FGF
Важность петли SOX9 – FGF была подтверждена в ряде экспериментов над генетически модифицированными мышами. Эмбрионы мышей, в клетках гонад которых Sox9 полностью отсутствовал, формировали признаки, характерные для самок, несмотря на присутствие Sry, так как Sry не может влиять на организм без работоспособного Sox9. Такой же эффект наблюдался после удаления сигнальной системы Fgf. Напротив, мыши, в гонадах которых экспериментально активировали экспрессию Sox9 независимо от присутствия Sry, развивались в самцов, даже если у них не было Y-хромосомы.
Под действием продуктов генов, активированных белком SOX9, клетки, продуцирующие этот белок, начинают активно размножаться, а также приобретают характерную форму и биохимические свойства клеток семенников, которые обеспечивают образование сперматозоидов. После того как эти клетки окончательно выбрали путь развития, они заставляют перейти к формированию семенника и все остальные клетки гонады (семенник содержит большое количество трубочек, в толстых стенках которых в свое время будут формироваться из клеток зародышевой линии сперматозоиды).
Мы только что обсудили мужской путь развития. Чтобы проследить путь женского развития, следует вернуться к тому моменту, когда в гонаде, еще не определившейся с выбором пола, образовывался белок WT1. Он действует далеко не только на ген SRY, а вызывает также последовательную активацию ряда других генов, находящихся на обычных хромосомах, имеющихся у всех эмбрионов, независимо от того, имеется у них ген SRY или нет. Мы пока не знаем все детали этой последовательной активации, но известно, что через несколько часов клетки начинают продуцировать еще один сигнальный белок из известного нам семейства WNT (см. главы 7, 10 и 11). Это белок WNT4. При условии отсутствия сильных ингибиторов его функции этот белок заставляет клетки гонад формировать яичник (рис. 58).
Понятно, что у мужчин путь WNT4 должен как-то блокироваться, и петля FGF – SOX9, активируемая белком SRY в мужских половых железах, как раз и является мощным ингибитором функций WNT4. Поэтому общий принцип заключается в следующем: «по умолчанию» эмбрион следует по женскому типу развития под руководством WNT4, а если гены, характерные для мужского пути, успели активироваться, женский путь развития подавляется.
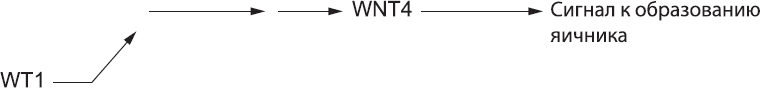
Рис. 58. Женский путь развития: экспрессия WT1 в конечном итоге приводит к экспрессии WNT4, а он подает клеткам гонады сигналы к образованию яичника
Как мы помним, мужской путь развития снабжен «переключателем» – сигнальной петлей SOX9 – FGF, который гарантирует, что клетки не сойдут с выбранного пути из-за случайных колебаний в системе. Женский путь развития также снабжен специальным переключателем, благодаря которому слабая или случайная активация «мужских» генов не приводит к полному сумбуру. После того как сигнальный путь WNT4 активирован, он ингибирует «мужской» путь, гарантируя, что никакой всплеск активности «мужских» генов ни в какой клетке не переведет «переключатель» в «мужское» состояние. Поэтому выбор пола – это вопрос борьбы за ручку переключателя. Если SRY присутствует и вовремя включает петлю FGF – SOX9, то развитие автоматически идет по мужскому пути, а женский путь WNT4 находится в положении «выключено». Если же SRY отсутствует или по какой-то причине не активирует петлю FGF – SOX9 вовремя (у мышей речь идет примерно о шести часах), то запускается женский путь развития, а мужской путь переходит в положение «выключен» (рис. 59).
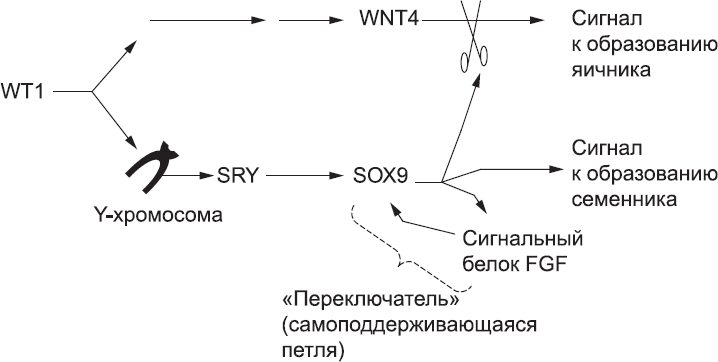
Рис. 59. Молекулярная логика определения пола. Появление в гонаде белка WT1 запускает последовательность событий, которая в конечном итоге приводит к синтезу WNT4, а он подает клеткам гонад сигнал к созданию яичника. Однако если у эмбриона есть Y-хромосома, WT1 также активирует находящийся на ней ген, который запускает производство белка SRY. Это стимулирует производство SOX9. Он имеет несколько функций: активирует сигнальный путь FGF, который поддерживает синтез SOX9, блокирует действие WNT4 («заглушая» сигнал к формированию яичников) и подает клеткам гонады сигнал формировать семенник
Важность сигнального пути WNT4 для выбора эмбрионом женского пути развития была показана во многих экспериментах над генетически модифицированными мышами. Мыши, в гонадах которых за счет какого бы то ни было генетического вмешательства был активирован сигнальный путь WNT4, развивались по женскому пути, даже если имели Y-хромосому.
Если основанный на WNT4 «переключатель» в соматических клетках гонады находится в положении «женский путь развития», то они будут развиваться в клетки, обеспечивающие формирование яйцеклеток. Они посылают сигналы, благодаря которым остальные клетки половой железы образуют ткани яичника, а не семенника (яичники не состоят из многочисленных трубочек, а заполнены рыхлой тканью и группами клеток, которые собираются вокруг развивающихся яйцеклеток). Один из сигналов этих клеток подталкивает зародышевые клетки к особому типу деления – мейозу, – который имеет решающее значение для формирования как яйцеклеток, так и сперматозоидов. В организме мальчиков мейоз начинается только после наступления половой зрелости, но у девочек все клетки зародышевой линии вступают в мейоз практически в самом начале развития яичников: на момент рождения девочки все ее яйцеклетки находятся в состоянии мейоза. Хотя мейоз начинается еще до рождения, он, как ни странно, приостанавливается в период примерно с двенадцати до пятидесяти лет, и только несколько яйцеклеток возобновляют свое развитие каждый менструальный цикл. Такой характер развития имеет одно неприятное медицинское следствие. Развивающиеся яйцеклетки, сделавшие паузу в мейозе, крайне уязвимы к лекарственным препаратам, которые иногда используется в химиотерапии, и для больных раком девочек и молодых женщин ценой спасения может оказаться бесплодие. К счастью, был разработан метод, позволяющий удалять участки ткани яичников перед началом химиотерапии, замораживать их, а затем трансплантировать обратно, если женщина, достигнув детородного возраста, решит, что хочет завести детей.
Мейоз, в отличие от обычного клеточного деления, не приводит к образованию двух дочерних клеток с теми же хромосомами, что были и у материнской. Обычная клетка человеческого тела перед делением имеет по две копии каждой хромосомы, одна из которых унаследована от матери, а другая – от отца. Есть две копии хромосомы 1, две копии хромосомы 2, и так далее. Если клетка женская, у нее есть две копии Х-хромосомы, а если клетка мужская, то у нее есть одна Х-хромосома и одна Y-хромосома (потому что, как я говорил выше, именно присутствие Y-хромосомы и белка SRY приводит к созданию мужского организма). При нормальном делении каждая хромосома копируется, и дочерние клетки получают тот же набор, что и материнская клетка, а вот при мейозе хромосомы распределяются между клетками таким образом, что одна из копий достается одной дочерней клетке, а вторая – другой. Поэтому в конечном итоге дочерние клетки обладают только одной копией каждой хромосомы. Это именно то, что нужно яйцеклетке или сперматозоиду – когда в процессе оплодотворения они сольются, у оплодотворенной яйцеклетки будет нормальное количество хромосом. Все яйцеклетки, образованные в процессе мейоза в организме женщины, несут исключительно Х-хромосомы. Однако в мужском организме половина сперматозоидов несет Х-хромосому, а вторая половина – Y-хромосому. Если яйцеклетку оплодотворит сперматозоид с X-хромосомой, у эмбриона будут две Х-хромосомы и ни одной Y-хромосомы, и родится девочка. Если же яйцеклетку оплодотворит сперматозоид с Y-хромосомой, то эмбрион получит Y-хромосому (а с ней и ген SRY), и родится мальчик. Примерное соотношение мужчин и женщин в человеческой популяции – 50/50 – непосредственно связано с характером распределения хромосом при мейозе.
Однако мейоз – это далеко не только распределение хромосом между дочерними клетками. В каждой паре хромосом копия, унаследованная от матери, и копия, унаследованная от отца, немного отличаются. Это никак не связано с полом родителей, а отражает их индивидуальные черты. ДНК хромосом со временем мутирует, поэтому в человеческой популяции существует множество слегка отличающихся версий каждой хромосомы. Это разнообразие хромосом – одна из причин различий между людьми (они также могут быть связаны с влиянием окружающей среды). На ранних стадиях мейоза две копии хромосомы сближаются, и происходит рекомбинация – сложный процесс, в ходе которого две хромосомы обмениваются участками, разрезая и снова склеивая свою ДНК. Это означает, что копии хромосом, которые передаются сперматозоиду или яйцеклетке, представляют собой гибридные версии копий, имеющихся в обычных клетках организма (рис. 60).
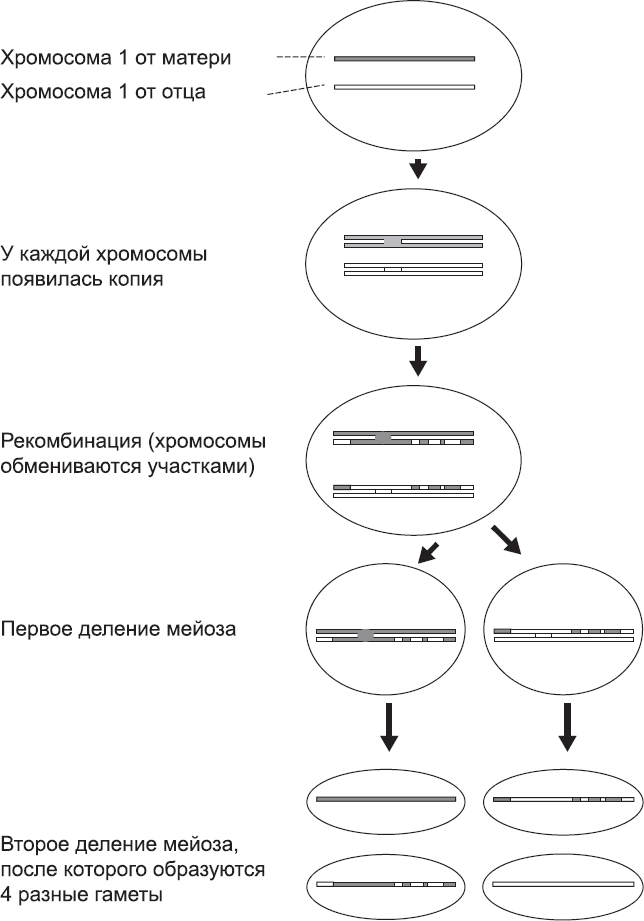
Рис. 60. Рекомбинация между хромосомами при мейозе (в данном случае в семеннике). Для лучшего восприятия на схеме изображена только одна пара из двадцати трех пар хромосом человека. Как и в случае любого деления, сначала все хромосомы копируются. Затем происходит событие, характерное только для мейоза, – унаследованная от матери копия хромосомы обменивается участками с копией, унаследованной от отца (происходит рекомбинация). После окончания этого этапа происходят два клеточных деления и образуются четыре гаметы, каждая из которых несет свою собственную (и отличающуюся от исходной) версию хромосомы 1. То же самое произойдет и с хромосомами 2–21, а вот хромосомы последней пары (X и Y) не рекомбинируют. Похожий процесс происходит и в яичниках женщины, но после каждого этапа деления одна дочерняя клетка погибает, и в результате образуется не четыре гаметы, а одна
Важно, что обмен участками хромосом при мейозе практически случаен, поэтому разные сперматозоиды (или яйцеклетки) одного и того же человека генетически отличаются друг от друга. Именно поэтому дети одних родителей могут быть очень разными.
По сути, рекомбинация, происходящая при мейозе в ваших половых железах, является первым моментом генетического объединения ваших родителей. Да, вы получили хромосомы и от матери и от отца, но они стоят особняком в каждой клетке вашего организма, нередко давая противоречивые указания по поводу вашего развития. Хромосомы ваших родителей постоянно «спорят» на языке функционирования генов о том, каким вам быть. «Глаза будут голубые!» – «Нет, карие!» – «Высокий!» – «Нет, коренастый!» – «Спокойный!» – «Нет, вспыльчивый!» И так далее. Одни черты, например цвет глаз, определяются относительно небольшим числом генов, а другие, например подверженность стрессам, – очень многими генами. Только после рекомбинации в процессе мейоза гены ваших родителей наконец встречаются и объединяются. После этого они уже готовы выступить единым фронтом и, когда придет время, «поспорить» с генами родителей вашего мужа или жены о том, каким будет ваш ребенок.
Однако определенный тип половых желез – не единственное, что делает человека мужчиной или женщиной. Как минимум существенно различается строение репродуктивной системы в целом, а к концу развития также накапливается множество менее значительных различий в строении других частей тела. У взрослых к таким отличиям относятся размер молочных желез (они развиты у женщин и недоразвиты у мужчин), форма костей (форма передней части таза немного различается у мужчин и женщин, и в том числе благодаря этому археологи могут определить пол скелета), размер костей (в пределах этнической группы мужчины в среднем крупнее женщин), размер мышц (опять же, у мужчин они крупнее), особенности роста волос (мужчины более волосаты), а также строение и функции некоторых частей мозга. Клетки во всех этих частях тела содержат ту же комбинацию хромосом (XX или XY), что и в гонадах, но нет никаких указаний на то, что клетки млекопитающих хоть как-то используют этот факт. Создается впечатление, что Y-хромосома, если она есть, неактивна везде, кроме гонад на раннем этапе развития. Вместо того чтобы «обратиться за советом» к собственным хромосомам, клетки остальных частей организма в основном полагаются на сигналы половых желез. Эти сигналы представляют собой мелкие молекулы, которые легко переносятся на большие расстояния, – гормоны.
Если на раннем этапе удалить эмбрионам кролика гонады, лишив их таким образом специфических гормонов, эмбрионы будут развиваться по женскому пути независимо от того, какую комбинацию хромосом – XX или XY – они несут. Этот эксперимент наводит на мысль, что не относящиеся к половым железам части эмбриона «по умолчанию» формируют женский организм и что для преодоления этих изначальных «настроек» нужны мужские гормоны, источником которых служат семенники.
После того как половая железа встала на путь формирования семенника (благодаря «переключателю» SOX9 – FGF9 – FGFR2), она начинает продуцировать два важных гормона – антимюллеров гормон и дигидротестостерон. Именно они в основном опосредуют влияние семенников на организм эмбриона. Антимюллеров гормон получил свое название из-за его действия на пару протоков, которые проходят в теле эмбриона параллельно вольфову каналу (рис. 61). Мюллеровы протоки есть у всех эмбрионов на ранней стадии, и если они сохраняются, то впоследствии дают начало внутренним протокам женской репродуктивной системы – маточным (фаллопиевым) трубам, матке и верхней части влагалища. Мужскому организму эти структуры не нужны, напротив, они серьезно мешали бы ряду аспектов развития. Сам по себе антимюллеров гормон безвреден, но клетки мюллерова канала синтезируют ряд белков, которые, если вокруг обнаруживается этот гормон, изнутри «подталкивают» эти клетки к «самоубийству». Избирательная гибель клеток, или «клеточное самоубийство», – распространенный и очень важный аспект развития животных, и я подробнее остановлюсь на этом явлении чуть позже (см. главу 14).
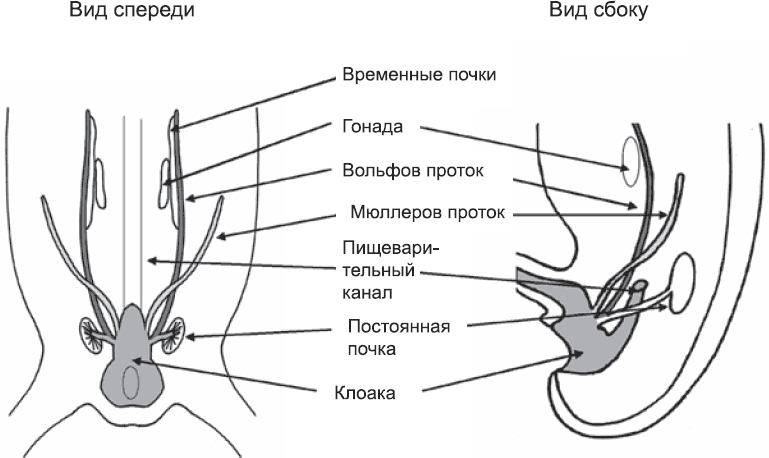
Рис. 61. Положение вольфова и мюллеровых протоков, а также связанных с ними структур в задней части развивающегося эмбриона
Итак, благодаря антимюллерову гормону мужской организм избавился от зачатков женской репродуктивной системы. Теперь дело за малым – построить соответствующую мужскую систему. Здесь в игру вступают вольфовы протоки. Они залегают по бокам тела эмбриона, проходя из первой временной почки мимо второй временной почки и развивающейся постоянной почки и открываясь в клоаку между зачатками ног (рис. 61). Таким образом, они находятся достаточно близко к развивающейся гонаде. В отсутствие тестостерона вольфовы протоки и почти все структуры временных почек отмирают за счет избирательной гибели клеток. Если тестостерон есть, вольфовы протоки не отмирают. Они сохраняют связь с временными почками, и некоторые ответвления почек подходят к семенникам, проникают в них и, окончательно утрачивая «почечные» функции, превращаются в каналы для транспорта спермы. Когда остальные структуры временных почек отмирают, эти ответвления сохраняются, а вольфов проток становится семявыводящим протоком, который у взрослых мужчин служит для прохода спермы от яичек к уретре, откуда она и извергается наружу.
Если же гонада вступает на женский путь развития, она не продуцирует ни антимюллеров гормон, ни тестостерон. В отсутствие антимюллерова гормона мюллеровы протоки сохраняются и с течением времени развиваются в женскую внутреннюю репродуктивную систему. Без тестостерона вольфовы протоки отмирают и почти полностью исчезают, что исключает возможность развития мужской половой системы.
Кроме внутренней половой системы, у эмбрионов должны возникнуть мужские или женские внешние половые органы. В отличие от механизма формирования внутренних структур, который основан на выборе одного из двух наборов тканей-предшественников, механизм образования внешних структур использует одни те же исходные ткани, но придает им разную форму – так из обычного листа бумаги можно сложить очень непохожие друг на друга фигурки оригами.
Внешние половые органы начинают формироваться из тканей, расположенных между зачатками ног эмбриона, примерно на пятой неделе развития. Ровно посередине между зачатками ног находится полость, которая называется клоака. На данном этапе она представляет собой общее отверстие пищеварительного канала, мочевыводящих и половых протоков (см. рис. 61). Клоака эмбриона напоминает нам о внутреннем строении наших далеких предков – рыб и рептилий. Пищеварительная, мочевыделительная и репродуктивная системы открывались у них наружу общим отверстием. Со временем благодаря росту тканей клоака подразделяется на два или три независимых отверстия. Ее передняя часть становится частью мочевого пузыря и мочеиспускательного канала, задняя часть – прямой кишкой, а центральная часть, куда открываются мюллеровы протоки, образует нижний отдел влагалища (это третье отверстие, разумеется, образуется только у женщин) (рис. 62).
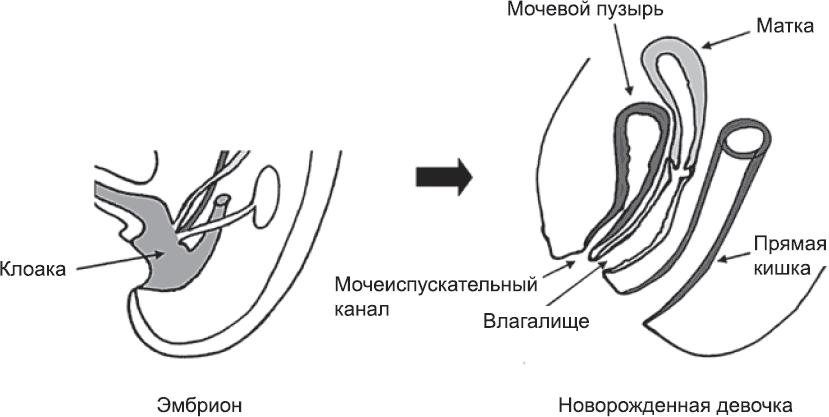
Рис. 62. В ходе разделения клоаки эмбриона образуются отдельные отверстия мочевыделительной, женской репродуктивной и пищеварительной систем
На пятой неделе развития около клоаки формируются три утолщения. Два из них – продолговатые валики, по одному с каждой стороны – проходят спереди назад и сходятся прямо перед третьим уплотнением – половым бугорком (см. рис. 62). По мере роста этот бугорок вытягивается в длину. У женщин он остается относительно небольшим и становится клитором, а у мужчин он существенно увеличивается и становится пенисом (рис. 63).
Валики по обеим сторонам клоаки растут. У женщин они не меняют своего положения и дают начало парным половым губам, образующим внешнюю границу женских половых органов. В мужском организме влагалища нет, и ничто не мешает этим валикам встретиться на срединной линии и сформировать непрерывную поверхность – мошонку. Семенники в конечном итоге опускаются в мошонку благодаря длинной связке, проходящей от половых желез к развивающимся гениталиям. Сначала эта связка подтягивает их в нужное положение просто за счет того, что не растет (поскольку остальная часть организма продолжает расти, она «обгоняет» семенники, оставляя их в нижней части), а на более поздних этапах подтягивает их прямо в мошонку за счет активного сокращения. Яичники женских эмбрионов тоже крепятся к связке, но она укорачивается гораздо менее сильно, подтягивая яичники только книзу таза.
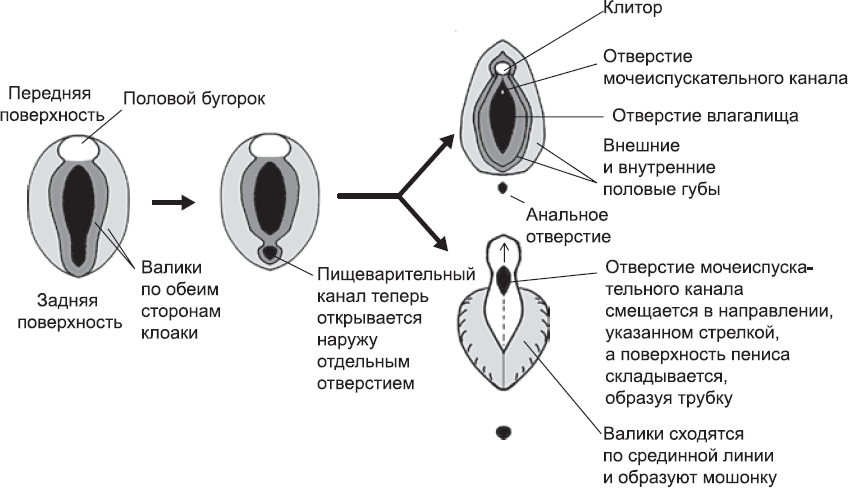
Рис. 63. Формирование мужских и женских наружных половых органов из одной и той же исходной ткани. На схеме показан участок между ногами развивающегося эмбриона (вид снизу). Сначала имеется отверстие клоаки, окруженное тремя утолщениями (педантичные анатомы делят их на подтипы, но, чтобы понять общую схему процесса, вполне достаточно знать, что утолщений три). После обособления анального отверстия и отверстия мочеполовой системы утолщения развиваются: половой бугорок увеличивается, превращаясь в клитор или пенис, а половые валики образуют половые губы или, смыкаясь по срединной линии, формируют мошонку
Создание мужского или женского организма – многоступенчатый процесс, задействующий сложнейшие механизмы развития. Неудивительно, что он может дать сбой. При этом, в отличие от серьезных ошибок на других этапах развития, например при формировании сердца, сбой при выборе пола не является несовместимым с жизнью. Поэтому нередки случаи, когда люди рождаются «не в том теле» относительно их хромосомного пола или представляют собой нечто среднее между мужчиной и женщиной, а бывает и так, что одни части тела – несомненно мужские, а другие – женские.
Носители мутаций, нарушающих нормальное функционирование SRY или SOX9, имеют мужской хромосомный набор и женское тело. Кроме того, для людей с мутацией, инактивирующей SOX9, характерен особый тип карликовости – кампомелическая дисплазия. Это связано с тем, что, в отличие от SRY, SOX9 также вносит вклад в развитие не связанных с полом частей тела. Мутации, приводящие к образованию дополнительных копий гена SOX9, оказывают обратный эффект. В 1999 г. был описан клинический случай, когда мальчик имел мужское тело и женский хромосомный набор (XX). Однако семнадцатая хромосома, на которой находится SOX9, присутствовала у него в двух экземплярах (это явление называется дупликацией хромосом). Очевидно, SOX9 синтезировался у него в таком большом количестве, что даже в отсутствие SRY мог перевести «переключатель» SOX9 – FGF в «мужское» положение.
Тот факт, что половые железы «оценивают» свой хромосомный набор и сообщают эту информацию остальному организму с помощью гормонов, создает предпосылки для интересных ошибок развития, когда человек имеет половые железы, характерные для одного пола, а тело – для другого. Гормон тестостерон, сообщающий телу мужские черты, можно обнаружить только с помощью особого рецептора. Если этот рецептор инактивируется в результате мутации, организм не сможет обнаружить присутствие тестостерона и пойдет по женскому пути развития, несмотря на наличие семенников. Многие люди с такой особенностью внешне никак не отличаются от женщин (однако у них нет менструальных циклов, потому что нет яичников, и нет структур, производных от мюллерова канала, потому что есть антимюллеров гормон).
Одна из невероятнейших половых аномалий у людей обусловлена мутацией, которая редко встречается в популяции в целом, но относительно часто отмечается у населения небольших островов. Она влияет на преобразование тестостерона в тканях. Хотя в массовой культуре тестостерон рассматривается как неотъемлемый атрибут мужественности, сам по себе он оказывает лишь слабый маскулинизирующий эффект. Однако в тканях под действием определенного фермента тестостерон превращается в значительно более сильный гормон – дигидротестостерон. Если мутация затрагивает ген, кодирующий этот фермент, тестостерон не вступает в нужную реакцию, а сам по себе он не может направить тело по мужскому пути развития. Родившийся ребенок будет иметь женское тело и в детстве ничем не отличаться от обычных девочек. Однако с наступлением половой зрелости у такого человека произойдет, как это обычно бывает у мужчин, всплеск активности тестостерона. Его будет столько, что при всей его «слабости» он сможет пустить организм по мужскому пути развития. Этого может оказаться достаточно, чтобы структура, которая раньше выглядела как клитор, превратилась в пенис, яички опустились в мошонку, а рост волос пошел по мужскому типу. Так, человек, который провел все детство девочкой, всю оставшуюся жизнь живет мужчиной (как Орландо, персонаж из романа Вирджинии Вулф, только наоборот).
Менее глубокие аномалии сигнальных путей и молекул-рецепторов могут вызывать менее резкую половую двойственность. Один человек с подобной двойственностью невольно внес значительный вклад в наше понимание полового развития. Инес де Торремолинос, жившая в XVI в., была вдовой и матерью троих детей, то есть, несомненно, женщиной, но из-за переизбытка тестостерона, вероятно вызванного эндокринным заболеванием, у нее развилась патология, ныне известная под названием клиторомегалии. При этой патологии клитор сильно увеличен, представляя собой нечто среднее между нормальным клитором и пенисом. Ее врач, естествоиспытатель Матео Коломбо, обратил внимание на этот необычный орган и по его промежуточной форме понял связь между развитием мужских и женских половых органов. Возможно, он был первым ученым, который понял, что мужские и женские гениталии – не разные структуры разного происхождения, а две версии одного и того же органа. Он провел своей пациентке, а затем и нормальным женщинам «терапию» (за которую в наше время его бы живо исключили из Списка практикующих врачей Генерального медицинского совета) и осознал в процессе своих «исследований» крайнюю сексуальную чувствительность клитора. Матео Коломбо опубликовал свои наблюдения в 1558 г., и его часто называют «первооткрывателем» сексуальной функции клитора. Это утверждение, конечно, звучит довольно странно: сексуальная чувствительность этого органа явно была известна мужчинам очень давно (судя, например, по древнеримской классической поэзии или непристойным изображениям в Помпеях), не говоря уже о том, что женщины, надо полагать, знали о ней всегда.
В ряде случаев частичные сбои определения пола могут быть вызваны не генетическими факторами, а влиянием окружающей среды. На протяжении жизни нескольких последних поколений репродуктивная функция мужчин в Западной Европе в среднем снизилась. Растет число доказательств того, что это отчасти связано с загрязнением окружающей среды – некоторые виды загрязнителей подавляют гормональные сигналы.,, Особое беспокойство вызывают фталаты, которые широко используются для повышения гибкости пластмасс. У лабораторных животных, по крайней мере, они сильно подавляют развитие половой системы. Также существует зависимость между воздействием фталатов на женщин и неполным развитием гениталий у их сыновей. Недавно в Европейском Союзе запретили использовать фталаты при производстве игрушек, но многие другие потенциально небезопасные соединения по-прежнему в ходу. Эта тема, неоднозначная сама по себе, осложняется политическими коннотациями. Мы пока не знаем точно, оказывают ли текущие уровни загрязнения существенное влияние на репродуктивную функцию человека, однако данные, полученные на лабораторных животных, как минимум должны послужить нам предостережением. Эмбриональное развитие – очень устойчивый процесс, но это не значит, что он устоит под натиском всех последствий людской беспечности.
Назад: Глава 11 Не покладая рук (и ног)
Дальше: Глава 13 В паутине проводов

