Книга: Онтогенез. От клетки до человека
Назад: Глава 10 Организация органов
Дальше: Глава 12 X или Y? Вот в чем вопрос
Глава 11
Не покладая рук (и ног)
Твоя рука свежа, легка,
Как дуновенье ветерка.
Уильям Вордсворт (пер. И. Меламеда)
Наличие конечностей – обязательное условие того образа жизни, который ведут млекопитающие. Конечности служат всем млекопитающим для движения и защиты, а людям и их ближайшим сородичам также для сложных манипуляций с предметами. В ходе эволюции позвоночные приобрели длинные конечности уже после того, как у них сложились основные анатомические черты туловища и головы. Поэтому и у эмбрионов конечности появляются только после формирования основного плана строения. Первые намеки на появление конечностей заметны у человеческого эмбриона примерно через двадцать четыре дня после зачатия, то есть тогда, когда уже основные структуры уже сформировались и функционирует примитивная система циркуляции.
Развитие конечностей начинается с появления двух небольших бугорков по бокам тела эмбриона немного выше уровня сердца. Это будущие руки. В скором времени появляются и бугорки в нижней части туловища – они дадут начало ногам. Появление бугорков связано с энергичным размножением клеток, залегающих непосредственно под эктодермой (напомню, что эктодерма покрывает зародыша снаружи). Дело не в том, что в этих областях клетки делятся более интенсивно, а в том, что скорость их размножения остается прежней, в то время как рост зародыша в целом замедляется. В результате пролиферация клеток в развивающихся конечностях происходит интенсивнее, чем в боковых частях тела. Клетки «принимают решение» о сохранении высоких темпов пролиферации не самостоятельно, а под действием сигналов мезодермы туловища. Клетки мезодермы на тех уровнях туловища, где в дальнейшем появятся руки и ноги, запускают экспрессию нового набора генов, (возможно, руководствуясь HOX-кодом; см. главу 6) и, в частности, начинают производить сигнальные белки семейства WNT. Эти белки активируют производство сигнальных молекул семейства FGF, а они, в свою очередь, запускают образование зачатков конечностей. Способность FGF вызывать формирование конечностей была продемонстрирована в весьма показательных экспериментах: кусочки геля, пропитанные этими белками, имплантировали куриному эмбриону между участками формирования крыла и ноги. Как вариант, в ткани куриного эмбриона вводили вирусы, подвергнутые генетической инженерии и кодирующие FGF. Клетки, расположенные над искусственными источниками FGF, формировали дополнительные выросты, из которых получились дополнительные конечности., Такой же эффект наблюдается при экспериментальной индукции производства сигнальных молекул WNT в этих местах. С другой стороны, если блокировать активность WNT или FGF, конечности не образуются даже там, где они должны быть.
После того как эти сигналы запускают развитие конечностей, тонкий слой эктодермы в области их формирования утолщается, и залегающие под ней клетки мезодермы начинают размножаться. Эта пролиферация «выталкивает» наружу почку конечности, постепенно принимающую лопатообразную форму с эктодермальным утолщением на конце (рис. 51).
Клетки, залегающие в толще почки конечности, могут давать начало разным тканям, например костям и сухожилиям. Понятно, что клетки не должны случайным образом решать свою судьбу, иначе получится не конечность, а сплошное безобразие. Они должны «принимать решение» в соответствии со своим положением. Клетки, которые в конечном итоге окажутся в районе кисти, должны сформировать мелкие косточки пальцев, а клетки в районе плеча – крупную плечевую кость. Клетки, расположенные между ними, образуют локтевой сустав, или кости предплечья, или, если они находятся вне участков формирования костей, сухожилия и мышцы. Таким образом, развивающейся конечности требуется надежная система, заставляющая клетки образовывать определенную ткань в зависимости от их расположения. Насколько мы знаем (знаем мы, правда, далеко не все), клетки конечности способны определять концентрацию разных сигнальных молекул, поступающих из разных участков конечности, примерно так, как это делают клетки сомитов (глава 7). Кроме того, их судьбу, как и судьбу клеток сомитов, возможно, в определенной мере контролируют «молекулярные часы».
Анатомию конечности можно описать с помощью системы координат трех осей: первая идет от плеча до кончиков пальцев, вторая – от большого пальца до мизинца, третья – через ладонь и тыльную сторону кисти. Хотя системы, задающие в ходе развития ориентацию этих осей, не полностью независимы, нам будет проще рассматривать их по одной, упоминая взаимодействия между ними по мере их появления.
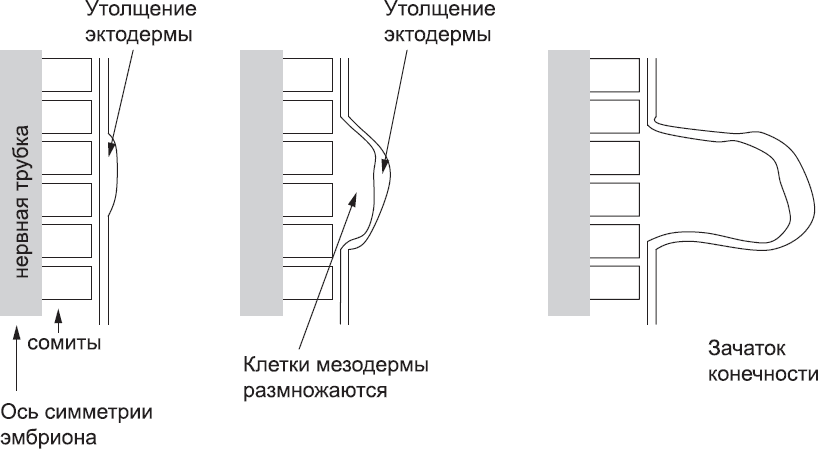
Рис. 51. Формирование почки конечности (в данном случае правой руки) на боку эмбриона. На участке, где должна сформироваться конечность, утолщается эктодерма, а в результате деления залегающих под ней клеток мезодермы появляется бугорок – почка конечности, которая затем приобретает лопатообразную форму
В настоящее время (точнее говоря, на момент написания этой книги) вопрос о том, как определяется положение структур конечности на оси, идущей от плеча к пальцам, является предметом бурных научных споров. Есть несколько гипотез, и у каждой есть свои приверженцы. Убедительно доказать или опровергнуть какую-либо из них пока не удалось. С одной стороны, это досадно, так как я не могу предложить вам однозначный ответ. С другой стороны, существующие разногласия предоставляют мне хорошую возможность продемонстрировать вам процесс познания в биологической науке.
Начну с ряда бесспорных фактов. Во-первых, конечности удлиняются постепенно, и основная пролиферация клеток происходит в так называемой зоне прогресса, расположенной в дистальной части конечности. По мере удлинения конечности зона прогресса оставляет позади себя клетки (так оставляет за собой след ползущая улитка), которые дифференцируются и формируют структуры конечностей. Во-вторых, клетки на обоих концах развивающейся конечности подвергаются влиянию разных наборов сигнальных молекул (или, во всяком случае, разного соотношения сигнальных молекул). Клетки эктодермы на самом конце зачатка конечности выделяют белки FGF., На плечевом конце в зачаток конечности поступает из туловища другая молекула – ретиноевая кислота (о ней уже шла речь, когда мы обсуждали образование сомитов). В-третьих, плечевые кости закладываются раньше, чем кости предплечья, а те, в свою очередь, – раньше, чем кости кисти.
Тот факт, что клетки, проводящие разное время в зоне прогресса, в конечном итоге создают разные части руки, как раз и лежит в основе одной из самых спорных современных моделей определения положения элементов конечностей. Согласно этой модели, время, проведенное в зоне прогресса, само по себе является сигналом, и чем больше времени клетки провели в ней, тем больше признаков, характерных для пальцев, приобретет сформированная ими структура (рис. 52). Клетки, которые рано покинули зону прогресса, образуют плечо, а клетки, покинувшие ее немного позже, – локоть. Клетки, запоздавшие еще сильнее, образуют предплечье, запястье и кисть, а клетки, которые, по сути, так и не покинули зону прогресса, – кончики пальцев. Эта изящная модель предсказывает, что если вынудить клетки провести в зоне прогресса аномально долгое время, то плечевой ткани в конечности будет мало, а ткани кисти – слишком много. Пытаясь создать именно такие условия, исследователи облучали зачаток конечности цыпленка рентгеновскими лучами. Под действием облучения многие клетки в зоне прогресса погибли, а те, которые выжили, провели в этой зоне больше времени, чем обычно, восстанавливая свою численность за счет пролиферации; лишь после этого некоторые из них смогли покинуть ее. В результате действительно образовалась конечность без плечевой кости, но со структурами кисти, в полном соответствии с предсказанием временной модели.
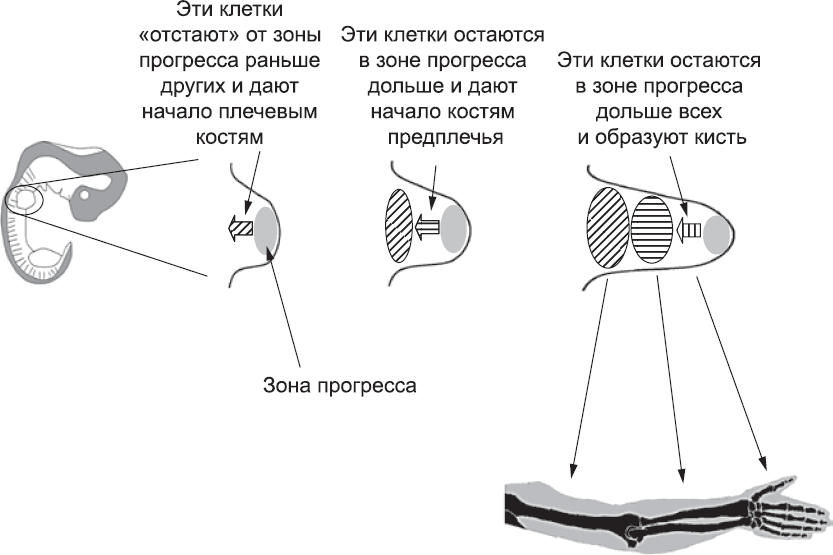
Рис. 52. Временная модель развития конечности: по мере удлинения зачатка конечности активно размножающиеся клетки «отстают» от зоны прогресса в разное время. Согласно этой модели, кончики пальцев руки, например, будут образованы клетками, которые провели больше всего времени в зоне прогресса. На схеме изображены только три стадии, соответствующие трем крупным отделам руки, но эта модель работает и для более детального уровня дифференцировки структур конечности
Проблема описанного выше эксперимента в том, что отложенный выход выживших клеток из зоны прогресса был лишь косвенным эффектом воздействия рентгеновских лучей. Их непосредственный эффект выражался в гибели многих других клеток. Резонно предположить, что отсутствие плечевых костей с тем же успехом могло объясняться разной чувствительностью разных типов клеток к рентгеновскому излучению. Недавно этот эксперимент был повторен с использованием современных методов, позволяющих отследить экспрессию генов, и выяснилось, что рентгеновские лучи никак не влияют на клетки, находящиеся на ранних этапах специализации в клетки плечевых структур. Изучение убивало только клетки, находившиеся в процессе создания видимых структур плеча. Таким образом, эксперимент, задумывавшийся для подтверждения временной модели, не оправдал надежд. Нельзя сказать, что эта модель была опровергнута, просто приходится признать, что она не столь хорошо подкреплена доказательствами, как считалось раньше.
Другая модель определения положения структур конечности никак не связана со временем. Она основана на существовании двух противоположных градиентов концентрации – белков FGF от кончиков пальцев к плечу и ретиноевой кислоты в обратном направлении. Суть в следующем: при таком распространении сигнальных молекул клетки в разных точках конечности подвергаются воздействию разного соотношения концентраций ретиноевой кислоты и FGF, и эта разница определяет различия в их дальнейшем поведении (рис. 53). Поскольку сигнальные молекулы могут распространяться лишь на небольшие расстояния, большинство версий этой модели постулируют, что клетки приобретают отличительные черты на ранних стадиях, когда зачаток конечности небольшой, даже если их дифференциация проявляется гораздо позже. Основная идея этой модели подтверждается экспериментами, в которых клетки извлекали из ранних зачатков конечностей и выращивали в чашках Петри. Если добавить в такую культуру чистые сигнальные белки, все клетки получат их в одинаковой концентрации. Под действием сигнальных белков FGF, которые в норме поступают из дистальной части зачатка конечности, в клетках подавлялась экспрессия генов, связанных с развитием плеча, и увеличивалась сначала экспрессия генов, связанных с развитием предплечья, а затем генов, связанных с развитием кисти. Ретиноевая кислота, источником которой в норме является плечевая часть руки, стимулировала формирование плечевых структур, даже если одновременно с ретиноевой кислотой в культуру добавляли белки FGF. Похожие результаты были получены и в серии экспериментов, в которых клетки зачатка конечности трансплантировали в другие участки эмбриона, богатые FGF или ретиноевой кислотой. В частности, эти эксперименты ясно показали, что для формирования верхней части руки нужна ретиноевая кислота или эквивалентная ей сигнальная молекула, поступающая со стороны туловища.
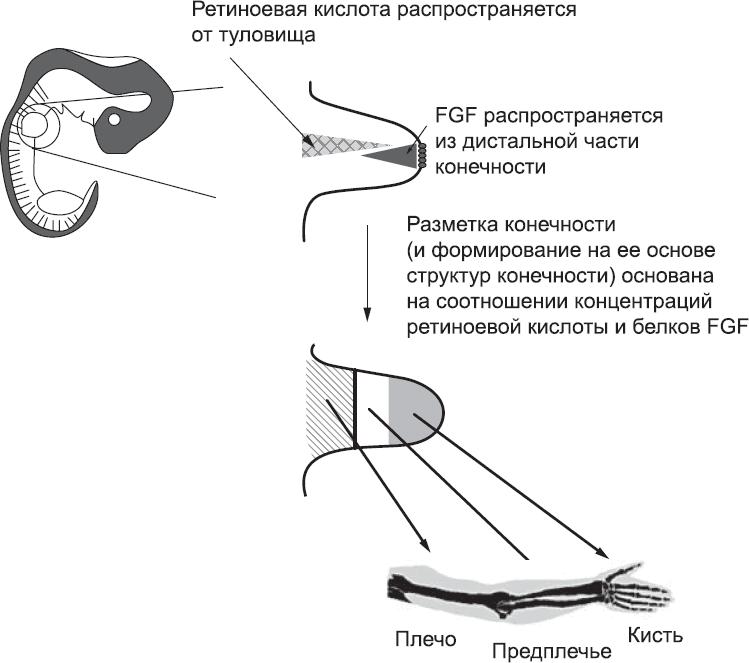
Рис. 53. Модель, основанная на соотношении концентраций сигнальных молекул. Противоположные градиенты концентрации – белка FGF из дистальной части конечности и ретиноевой кислоты, поступающей из туловища, – активируют разные паттерны экспрессии генов. Клетки, которые подвергаются воздействию высокой концентрации FGF и низкой концентрации ретиноевой кислоты, образуют структуры кисти, а клетки, на которые белок FGF действует в низкой концентрации, а ретиноевая кислота – в высокой, образуют структуры плеча. Точные уровни концентрации FGF и ретиноевой кислоты определяют, образует ли клетка структуры, характерные для кончиков пальцев или для основной части кисти, для локтя или для плеча
Если модель, основанная на соотношении концентраций сигнальных молекул, верна, то добавление дополнительной порции FGF в зачаток конечности должно вызывать образование более обширной зоны кисти или пальцев в ущерб остальной части руки. Однако этого не наблюдается – зона клеток, идущая на формирование кисти, имеет такой же размер, что и обычно. Это говорит о том, что модель, основанная на соотношении концентраций сигнальных молекул, не вполне верна, во всяком случае для развития области кисти.
Итак, на данный момент у нас есть по меньшей мере две основные модели развития конечности. Каждая из них подтверждается рядом фактов, но расходится с результатами как минимум одного эксперимента. Когда в науке возникает проблема такого рода, полезно подумать, нет ли каких-то неочевидных допущений, присущих обеим моделям. В нашем случае такое скрытое допущение есть, и оно заключается в том, что разметка всей конечности от плеча до кончиков пальцев осуществляется с помощью одного и того же механизма. Возможно, это неверно. В эволюции сложная конечность возникла не сразу. В грудных плавниках лопастеперых рыб, например, есть структуры, гомологичные плечу и предплечью, но нет ничего, что соответствовало бы кисти. Значит, развитие кисти «добавилось» к развитию конечности позже. Это подтверждается тем, что при развитии конечностей наземных животных экспрессия генов проходит те же основные стадии, что и у рыб, а затем появляется совершенно новый, отсутствующий у рыб этап. Более примитивные бесчелюстные рыбы имеют (или, в случае вымерших видов, имели) даже более примитивные плавники. Таким образом, возможно, что части конечности, расположенные на разных уровнях оси «плечо – кончики пальцев», размечаются за счет разных механизмов (рис. 54). Тогда возможно, что модель, основанная на соотношении сигнальных молекул, правильно объясняет, скажем, особенности формирования плечевой части – сигнальная молекула (ретиноевая кислота), источником которой является туловище, «защищает» прилегающую часть руки от превращения в какую-либо другую область конечности. А особенности образования предплечья и кисти, которые после удлинения зачатка конечности оказываются за пределами распространения сигналов от туловища, возможно, верно объясняются временной моделью. Тогда становится понятно, почему повышенная доза белка FGF не увеличивает размер кисти в ущерб плечу.
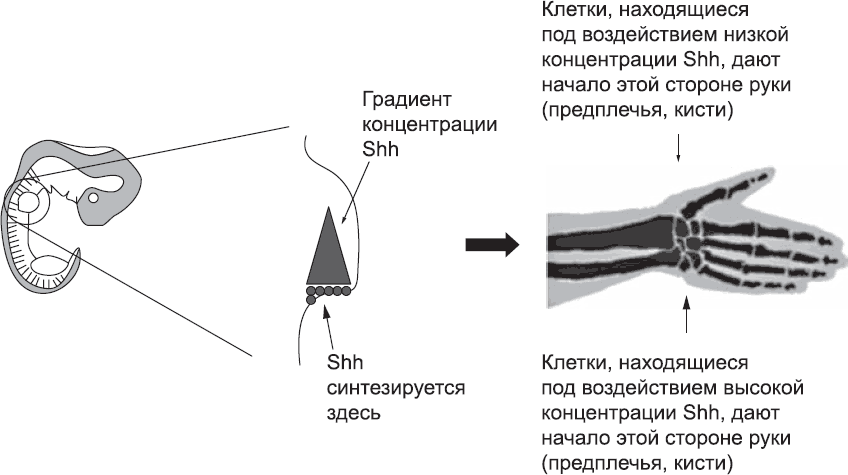
Рис. 54. «Принимая решение» о том, какой палец или какую кость предплечья (лучевую или локтевую) они должны сформировать, клетки руководствуются концентрацией белка Sonic Hedgehog (Shh), который синтезируется в конечности со стороны мизинца
Однако закладка и разметка проксимально-дистальной оси – лишь часть проблемы формирования структур конечности, существует еще две оси. Ориентация и разметка оси, идущей от мизинца до большого пальца, в основном контролируются группой клеток, залегающей со стороны мизинца. Они синтезируют и секретируют сигнальный белок, который распространяется по всей конечности, но его концентрация снижается по мере распространения. И на этот раз речь опять идет о сигнальном белке Sonic Hedgehog. Клетки кисти, которые его секретируют и получают в высокой концентрации на ранних этапах развития, формируют структуры конечности, располагающиеся со стороны мизинца. Клетки, которые получают на ранних этапах меньшее количество этого белка, формируют средние пальцы, а те клетки, которые получают очень мало этого белка или вообще его не получают, дают начало большому пальцу (рис. 54). Такой же принцип действует и в области предплечья, только здесь клетки, получающие высокую концентрацию белка Sonic Hedgehog, образуют локтевую кость, а те, которые получают этот белок в меньшей концентрации, – лучевую, и так далее.
Предположение о том, что тип пальца, который образуют клетки, зависит от полученного ими количества белка Sonic Hedgehog (Shh), было проверено в одном примечательном эксперименте, проведенном на почке крыла зародыша цыпленка (крыло птицы гомологично руке человека). Исследователи поместили еще один источник белка Shh на «неправильную» сторону ранней почки конечности цыпленка, то есть туда, где у людей образуется большой палец. Таким образом, по обеим сторонам почки конечности концентрация белка Shh была высокой, а самая низкая концентрация была в средней части. На крыле такого эмбриона образовался второй комплект пальцев, причем в обратном порядке. Со стороны мизинца последовательность пальцев была нормальной (1-2-3 по принятой на настоящий момент анатомической номенклатуре человека; пятого, или большого, пальца у кур нет). Правда, второй палец образовался ближе к середине конечности, где концентрация Shh была ниже всего. Вслед за ним образовался еще один второй палец, затем шел еще один третий, а затем еще один четвертый. В результате вместо нормальной последовательности пальцев (1-2-3) крыло эмбриона имело последовательность 1-2-3-3-2-1 (рис. 55). Образование таких аномальных конечностей, полностью согласующееся с теоретическими предсказаниями, говорит о том, что механизм, основанный на градиентах концентрации, в целом верен, хотя многие детали его работы, безусловно, еще предстоит изучить.
Относительно координат третьей оси конечности задаются различия между ладонью и ее тыльной стороной, а также определяется направление, в котором будет сгибаться локтевой сустав. И на этот раз почка конечности использует источник сигнальных молекул, расположенный на одной из ее сторон. Сигнальной молекулой на этот раз служит белок из семейства WNT (WNT7a), который синтезируется только с той стороны конечности, которая в дальнейшем станет тыльной стороной ладони. WNT7a подавляет формирование признаков, характерных для ладони: у мутантных животных, лишенных функционального гена WNT7a, структуры, характерные для ладони, развиваются с двух сторон конечности.
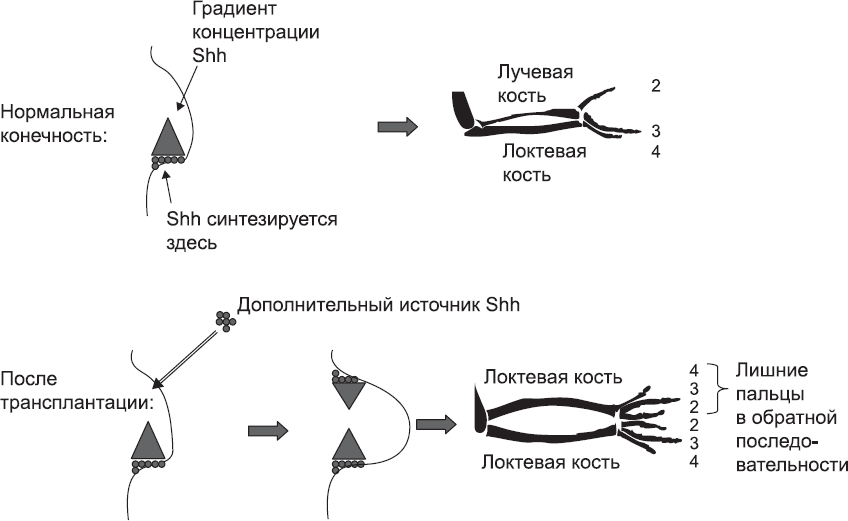
Рис. 55. В ходе нормального развития почки передней конечности цыпленка образуется крыло с тремя пальцами, гомологичными первому, второму и третьему пальцу человека. Если поместить на противоположной стороне почки крыла второй источник Shh, обычный комплект пальцев дублируется в зеркальном отражении, а обе кости предплечья развиваются в локтевые кости
Итак, складывается впечатление, что конечность контролирует свою разметку при помощи градиентов концентрации сигнальных молекул. Градиенты расположены под прямым углом друг к другу, охватывая все три пространственных измерения. Геометрически эти оси независимы, но с точки зрения биохимической активности они сплетены тесным и еще непонятным нам образом. Экспериментальное вмешательство в одну из осей нарушает, по крайней мере частично, и формирование структур относительно двух других осей. Сигнал WNT7a, определяющий положение структур относительно оси «ладонь – тыльная сторона руки», необходим для производства нормального количества белка Shh со стороны мизинца. Хотя бы небольшое количество белка Shh нужно для поддержания нормального уровня FGF. В свою очередь, Shh и FGF необходимы для поддержания нормального уровня WNT7a. Наших знаний о развитии конечностей недостаточно, чтобы понять, зачем нужна такая взаимозависимость. Возможно, она помогает поддерживать правильное соотношение сигналов, поступающих из разных источников.
По мере роста конечностей возникает потребность в эффективной кровеносной системе, обеспечивающей транспорт кислорода и питательных веществ к размножающимся клеткам. Новые сосуды образуются в ответ на сигналы от ткани точно так же, как и в других системах (см. главу 9). Проблема в том, что конечности удлиняются очень быстро, и кровеносная система должна формироваться так же быстро, чтобы от них не отставать. В противном случае клеткам конечности будет не хватать питательных веществ, и их размножение замедлится, что приведет к образованию маленьких, укороченных, аномальных конечностей. Печальной иллюстрацией этого служат последствия одной медицинской ошибки, пустившей под откос жизнь примерно десяти тысяч людей в 1958–1961 гг.
Причиной трагедии стало благое намерение смягчить или даже полностью устранить утреннюю тошноту. Это состояние, нередкое на ранних сроках беременности, может быть крайне изнурительным. Очень эффективным в борьбе с утренней тошнотой оказался один препарат, уже использовавшийся в качестве успокоительного и противовоспалительного средства, и его стали активно назначать беременным женщинам. Это был талидомид. В 1958 г. еще не знали (на самом деле выяснилось это только в начале этого века), что молекула, образующаяся в результате распада талидомида, ингибирует рост новых кровеносных сосудов. Это достаточно сильный эффект, и если будущая мать принимает талидомид в то время, когда быстро растущие конечности больше всего нуждаются в новых кровеносных сосудах, образование сосудов начнет отставать, и рост конечности замедлится. В крайнем случае это может привести к полному отсутствию конечностей или к образованию маленьких кистей или стоп непосредственно на туловище. Не совсем понятно, почему кисти рук и стопы продолжают развиваться, когда остальная часть конечности уже не растет, но если недостаток питательных веществ заставляет клетку задержаться в зоне прогресса, это явление хорошо согласуется с временной моделью формирования конечности.
На выявление связи между внезапным всплеском рождения младенцев с деформированными и укороченными конечностями и талидомидом ушло два года. К 1961 г. связь была доказана, и талидомид перестали прописывать от утренней тошноты. Следует заметить, что его продолжают использовать как лекарство от некоторых других болезней, включая проказу, и один-два врожденных дефекта в год (в среднем по миру) связаны с его применением в этих целях. Талидомид вернулся и на Запад, так как он эффективен при некоторых глазных и раковых заболеваниях, причем именно за счет способности подавлять рост кровеносных сосудов. Однако следует крайне внимательно следить за тем, чтобы препарат не назначался женщинам, которые потенциально могут забеременеть.
Назад: Глава 10 Организация органов
Дальше: Глава 12 X или Y? Вот в чем вопрос

