Книга: Онтогенез. От клетки до человека
Назад: Глава 6 Долгое деление
Дальше: Глава 8 Внутренние путешествия
Часть II
Постепенная детализация
Глава 7
Судьбоносные беседы
In principio erat verbum…Св. Иероним Стридонский
Процессы, описанные в главах 2–6, имеют внушительные последствия – бесформенное скопление клеток осталось далеко в прошлом. Теперь у эмбриона есть продольная ось тела с головным и хвостовым концами, дорсально расположенная нервная трубка и вентрально расположенный кишечник. По обе стороны нервной трубки залегают сомиты, а снаружи эмбрион покрыт эктодермой. Более того, благодаря избирательной экспрессии HOX-генов поведение клеток эмбриона специфично для уровня, на котором они находятся. Тем не менее на этой стадии развития эмбрион отличается от взрослого человека примерно так же, как эскиз отличается от законченного полотна: контуры видны, но их еще предстоит проработать.
На стадии формирования сомитов общее количество типов клеток в организме невелико. Есть клетки эктодермы, клетки нервной трубки, клетки кишечника, клетки сомитов и некоторые другие, но это лишь малая часть типов клеток, имеющихся у взрослого человека. Клетки первичных тканей эмбриона должны поделиться на группы, а группам предстоит специализироваться и дать начало разным структурам: костям, сухожилиям, мышцам и кровеносным сосудам. Более того, клетки должны сделать это по определенному алгоритму. Когда эмбрион впервые столкнулся с проблемой превращения одинаковых клеток в клетки разных типов (то есть когда у него обособилась внутренняя клеточная масса и трофэктодерма; глава 3), он использовал естественную асимметрию среды: клетки, часть поверхности которых была обращена в свободное пространство, дали начало трофэктодерме. Клетки нервной трубки и сомитов тоже полагаются в процессе дифференциации на сигналы, связанные с асимметрией окружающей среды. К этому моменту, однако, большая часть поступающей информации связана уже не с геометрическими характеристиками (такими, как наличие у клеток свободной поверхности), а с сигнальными молекулами, выделяемыми другими тканями. С помощью этих молекул соседние ткани ведут судьбоносные беседы и, таким образом, провоцируют друг у друга дифференцировку многих типов клеток, взаимное расположение которых исключительно точно задано в пространстве.
Хороший пример этих «клеточных бесед» – дифференцировка специализированных зон нервной трубки, которые в дальнейшем дают начало разным частям нервной системы. Нервная трубка, образованию которой была посвящена глава 5, проходит по средней линии эмбриона между эктодермой и нотохордом. Таким образом, нервная трубка оказывается естественным образом асимметричной среде: ее дорсальная поверхность ближе к эктодерме, а вентральная – к нотохорду. Эта асимметричная близость к нотохорду очень важна, потому что его клетки секретируют белок, получивший название Sonic Hedgehog («ежик Соник»). Концентрация этого белка меняется по градиенту: она максимальна вблизи нотохорда и снижается по мере удаления от него. Клетки нервной трубки чувствительны к белку Sonic Hedgehog, и те из них, которые расположены ближе к нотохорду, стимулируются очень сильно (рис. 30). В результате эти клетки начинают синтезировать новые белки и отличаться от других клеток нервной трубки. С этого момента они называются клетками вентральной пластинки.
Роль нотохорда в определении положения вентральной пластинки была показана в двух дополняющих друг друга экспериментах. В одном эксперименте в эмбрион цыпленка был хирургическим путем помещен дополнительный нотохорд, но не в обычное положение, а сбоку от нервной трубки. В нервной трубке этого эмбриона сформировались две вентральные пластинки: одна там, где положено (то есть над естественным нотохордом), а вторая на той стороне нервной трубки, которая была обращена к трансплантату. Это подтверждает гипотезу о том, что положение вентральной пластинки контролируется нотохордом. В другом эксперименте нотохорд был полностью удален, и вентральная пластинка вообще не сформировалась.
Среди белков, синтезируемых клетками вентральной пластинки, есть и сам Sonic Hedgehog, таким образом, она становится новым центром производства молекул этого белка, которые начинают распространяться в дорсальном направлении. Sonic Hedgehog – короткоживущий белок, поэтому он не может уйти далеко от места своего производства, и его концентрация резко падает с увеличением расстояния. Поведение клеток остальной части нервной трубки зависит от того, какое количество этого белка они получают, а это, конечно, зависит от их расстояния до вентральной пластинки.
Реакция клеток нервной трубки на градиент концентрации белка Sonic Hedgehog заключается в том, что они встают на путь специализации, который должен (в конечном итоге) привести к образованию одного из нескольких базовых типов нервных клеток. К числу таких базовых типов относятся двигательные нейроны, которые посылают сигналы непосредственно мышцам, и вставочные нейроны, которые принимают сигналы от других нейронов, обрабатывают их и передают дальше.
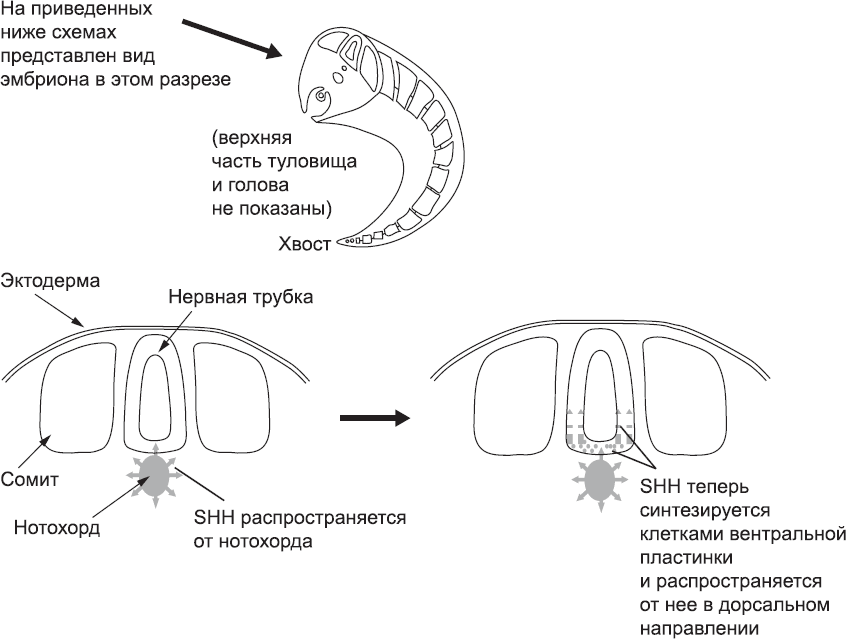
Рис. 30. Нотохорд производит белок Sonic Hedgehog (SHH), который распространяется к прилегающим тканям нервной трубки. Под его воздействием эти ткани образуют вентральную пластинку, которая тоже начинает синтезировать этот белок
Двигательные нейроны и вставочные нейроны находятся в разных зонах вентрально-дорсальной оси спинного мозга. Они организованы принципиально разным образом: чтобы нормально функционировать, клетка должна быть или двигательным нейроном, или вставочным нейроном, но ни в коем случае не половинчатым гибридом, сочетающим черты нейронов двух типов. Поэтому возникает проблема: по законам физической химии градиент концентрации белка Sonic Hedgehog может быть только плавным, а вот реакция клеток должна быть «ступенчатой» – сначала возникает один тип клеток, потом другой. Трансляция плавного градиента сигнала в ступенчатый отклик клеток осуществляется за счет целой серии взаимодействий различных генов и белков.
Основная роль белка Sonic Hedgehog заключается в активации специфичных генов. Различные гены обладают разной чувствительностью к этому белку. В клетках развивающейся нервной трубки есть как «отзывчивый» ген, который активируется даже при низких концентрациях Sonic Hedgehog, так и «равнодушный» ген, который активируется только при высокой концентрации этого белка. «Отзывчивый» ген настолько восприимчив, что активизируется сразу во всей вентральной половине нервной трубки, а «равнодушный» – только в клетках самой вентральной четверти нервной трубки, где концентрация белка достаточно высока. При этом присутствие в конкретной клетке белка, синтезированного «равнодушным» геном, отключает «отзывчивый» ген в этой клетке. Таким образом, непосредственно над вентральной пластинкой располагается зона клеток, экспрессирующих «равнодушный» ген, а над ней залегает слой клеток, экспрессирующих «отзывчивый» ген. Эти зоны не перекрываются. В результате формируются обособленные полосы (рис. 31).
Вентральная пластинка наряду с нотохордом принимает участие в разметке преимущественно нижней (вентральной) части нервной трубки. Однако верхняя (дорсальная) часть нервной трубки тоже должна быть размечена, и это достигается опять же за счет сигнала от прилегающих тканей. Ближе всего к дорсальной части нервной трубки залегает эктодерма, от которой совсем недавно отделилась сама нервная трубка (глава 5). Она выделяет еще один сигнальный белок, который легко распространяется до нервной трубки. Этот белок оказывает двоякое действие. Во-первых, он «отменяет» любые сигналы белка Sonic Hedgehog, если вдруг они смогли распространиться до этих районов нервной трубки. Во-вторых, под действием этого белка дорсальная часть нервной трубки сама становится центром распространения сигналов. Это очень похоже на то, что произошло в вентральной пластинке, когда под действием нотохорда центром распространения сигналов стала нервная трубка. На этот раз, однако, все не так просто. Клетки вентральной пластинки производят молекулы того же белка, который они получают от нотохорда (Sonic Hedgehog). С клетками дорсальной стороны нервной трубки ситуация иная: под действием сигнала от эктодермы они производят не тот белок, который получают, а новые сигнальные белки – WNT и BMP.
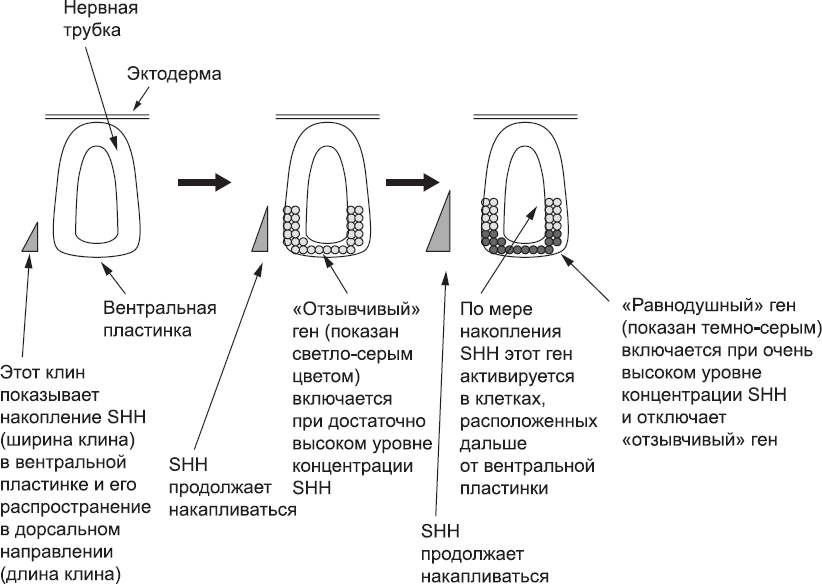
Рис. 31. Градиент концентрации белка Sonic Hedgehog (SHH) возрастает, пока он синтезируется в вентральной пластинке. Клетки, подпадающие под действие этого белка, начинают специализироваться, и образуются разные зоны
Белки WNT и BMP тоже распространяются из клеток, в которых синтезируются, с образованием градиента концентрации. Этот градиент используется для образования специализированных зон в дорсальной части нервной трубки примерно так же, как градиент белка Sonic Hedgehog используется для зонирования вентральной части нервной трубки (эти процессы отличаются целым рядом молекулярных подробностей, но общая идея одна и та же).
Затем нервная трубка использует сигналы, поступающие из асимметрично расположенных тканей сверху и снизу, а также, возможно, от сомитов, чтобы аккуратно разделить ткань, состоящую из схожих клеток, на зоны с определенной структурой, каждая из которых по мере дальнейшего развития спинного мозга даст начало особому типу нервной ткани. Однако клетки нервной трубки не только получают сигналы от других тканей, но и генерируют свои собственные сигналы. Распространяясь в обратном направлении, они вносят вклад в разметку прилегающих к нервной трубке тканей. Таким образом, соседние ткани не отдают односторонние приказы, а как бы ведут разговор, обмениваясь репликами на языке биохимии белков.
Сразу после образования сомиты представляют собой простые структуры, состоящие из одного типа клеток (глава 6). Однако они дадут начало многим структурам тела, в том числе костям, мышцам, сухожилиям и внутреннему слою кожи. Поэтому внутри сомитов, как и в нервной трубке, должны возникнуть отличающиеся друг от друга области, и это тоже происходит за счет сигналов от соседних тканей.
Участки сомитов, расположенные ближе всего к дорсальной части нервной трубки, находятся под действием высоких концентраций белков семейства WNT (рис. 32). Клетки сомитов уже экспрессируют несколько иной набор генов, чем клетки нервной трубки (именно поэтому они являются клетками сомитов). Как следствие, они реагируют на сигналы белков WNT иначе, чем клетки нервной трубки. Наверное, каждый из нас сталкивался с тем, что интерпретация сигнала может зависеть от внутреннего состояния того, кто его получает. При слове suspenders англичанин и американец представят себе совершенно разные вещи, и один мой американский знакомый, не знавший об этой разнице в словоупотреблении, однажды оказался в прекурьезной ситуации в фешенебельном эдинбургском универмаге. Это далеко не единственный пример. Фразу I propose we table this motion в Англии поймут как приглашение к немедленному обсуждению вопроса, а в Америке – как предложение отложить обсуждение на потом. Обилие повседневных слов и выражений, способных сбить с толку американца в Великобритании или англичанина в Америке, является хорошей иллюстрацией того, что даже в человеческих языках смысл сообщения во многом определяется не его содержанием, а внутренним состоянием адресата. С подобным явлением мы сталкиваемся и в биологии: один и тот же сигнал, например молекула белка WNT, может иметь разное значение (читай, оказывать разное воздействие) в зависимости от особенностей получающей его клетки.
Когда клетки сомита получают сигналы WNT из дорсальной части нервной трубки, они начинают производить белки, необходимые для формирования мышц. Белок WNT распространяется лишь на довольно небольшое расстояние, поэтому сигнал для активации развития мышц получает только та часть сомита, которая находится ближе к нервной трубке. WNT, поступающий из другого источника, запускает образование второго центра развития мышц на наружном, нижнем крае сомита (рис. 32). В дальнейшем из клеток этих зон образуются разные мышцы: зона около нервной трубки даст начало мышцам спины, а другая зона – мышцам стенки тела. Зона между ними, расположенная вдали от обоих источников WNT, станет соединительной тканью кожи (дермой).
Если бы на сомит действовали только те три типа сигналов, о которых мы только что говорили, он стал бы похож на сэндвич, в котором зона, дающая начало коже, залегает между двумя зонами, образующими мышцы.
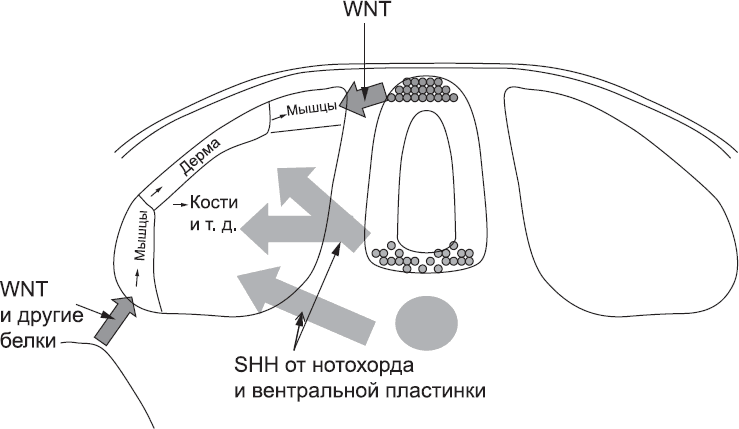
Рис. 32. Разметка областей сомита, которые будут дифференцироваться в разных направлениях, происходит за счет сигналов от соседних тканей
Однако сомит также чувствителен к сигналам, поступающим от нотохорда и вентральной пластинки нервной трубки. В участках сомита, расположенных ближе всего к этим источникам сигналов, концентрация соответствующих сигнальных белков достаточно высока. Они блокируют сигналы, стимулирующие образование мышц и кожи, и «сообщают» клеткам, что они должны образовать соединительную ткань и кости, (рис. 32).
Итак, обобщая сказанное в этой главе, мы видим, что окружающие ткани подают сигналы, за счет которых происходит дифференциация нервной трубки, а клетки нервной трубки реагируют на эти сигналы, в том числе и тем, что подают сигналы, под действием которых происходит разметка окружающих тканей и они приступают к дифференцировке. Таким образом, тонкая детальная разметка строения эмбриона, который еще недавно имел довольно однородное строение, является результатом многочисленных «обменов репликами» на клеточном уровне.
Ткани, о которых шла речь в этой главе, не уникальны. Те же самые общие принципы можно было бы продемонстрировать на примере любой другой группы соседних тканей. Во всем эмбрионе смежные участки тканей используют друг друга как источник сигналов для выделения внутри себя областей, клетки которых будут подвергаться дифференцировке, характер которой зависит от их расстояния до источника сигнала. Непосредственным результатом этого процесса является образование новых границ между только что дифференцировавшимися типами клеток в пределах исходно гомогенного участка ткани. Если эти типы клеток секретируют разные сигнальные белки, тот же прием можно использовать еще раз для создания еще большего количества отличающихся друг от друга областей. Это высокоэффективный механизм пространственной дифференцировки, и неудивительно, что примерно пятая часть генов человека нужна для производства белков, вовлеченных тем или иным образом в процесс клеточной сигнализации.
Клеточная коммуникация не только позволяет тканям дифференцировать внутри себя практически неограниченное число областей. Она также служит отличным механизмом борьбы с ошибками в процессе развития. Представьте себе эмбрион, в котором дифференциация участков тканей не зависела бы от положения окружающих тканей, а происходила бы каким-то иным путем, например, клетки действительно следовали бы пошаговым инструкциям к какому-то чертежу. Малейшие неточности в положении конкретных клеток приводили бы к постепенному накоплению ошибок; когда пришло бы время тканям собираться вместе, типы клетки, которые должны были бы соседствовать, оказались бы в разных местах, и развитие остановилось бы. В принципе, таким образом можно построить очень маленький и простой организм – при малом числе типов тканей и жизненно важных связей между ними ошибки просто не успеют накопиться. Однако создать «по инструкциям» организм, состоящий из сотен разных типов клеток, которые должны тонко взаимодействовать друг с другом, нельзя. Напротив, если ткани в системе полагаются на сигналы друг друга, дифференциация клеток в каждой из них автоматически происходит на правильном расстоянии от ткани, сигналы которой управляют этим процессом, даже если, скажем, эта «управляющая» ткань находится немного не там, где надо. Таким образом, организация системы подстраивается к меняющимся обстоятельствам, и ошибки не накапливаются, а корректируются на каждом этапе. Поэтому эмбрион способен справиться практически со всеми ошибками, за исключением серьезных аномалий, – постоянное «общение» клеток позволяет регулировать развитие в соответствии с реальной ситуацией, а не с абстрактным планом.
Использование общения на языке белков для регуляции дифференцировки тканей имеет одно интересное следствие для развития животных. Расстояние, на которое может распространяться белок в рабочих концентрациях, ограничено биофизическими и биохимическими законами и составляет для большинства белков около одной двадцатой доли миллиметра (50 мкм). Это означает, что длина группы клеток, использующих этот метод для нанесения разметки, предшествующей клеточной дифференцировке, как правило, составляет примерно 1/20 мм. Это относится как к расстоянию между дорсальной и вентральной частями нервной трубки, так и, например, к расстоянию между развивающимися корнями зубов или волосками. Из этого следуют две вещи. Во-первых, невозможно нанести разметку на весь эмбрион сразу. Сначала, пока эмбрион еще достаточно мал, нужно нанести грубую разметку, а потом, когда уже намеченные части увеличатся в ходе роста эмбриона, можно приступать и к их детализации. Это одна из причин того, почему человеческое развитие не может идти по пути создания крошечного, но полноценного ребенка, который затем только увеличивался бы в размерах. Последовательные фазы детализации должны быть распределены по этапам роста. Сначала обособляются голова и туловище, потом, когда голова вырастает, намечается, например, челюсть, затем, когда увеличивается челюсть, намечается положение зубов и так далее.
Второе следствие заключается в том, что нанесение любой разметки, например выделение зон нервной трубки, может произойти только на определенном этапе развития, когда размер эмбриона находится в довольно строгих границах. В частности, это означает, что на этом этапе развития зародыши землеройки, человека и синего кита должны быть практически одного размера. Эволюционное сходство животных – скажем, лошадей, китов и летучих мышей, которые, будучи очень разными, относятся к одному и тому же классу, – гораздо отчетливее видно на эмбрионах, чем на взрослых особях.
Назад: Глава 6 Долгое деление
Дальше: Глава 8 Внутренние путешествия

