Книга: Приспособиться и выжить!
Назад: Глава 6 Deja vu. Как и почему эволюция повторяется
Дальше: Одним путем к одной цели
Второе пришествие трихроматических обезьян
Наличие полноценного цветового зрения у ревунов, обнаруженное во время масштабного исследования обезьян Нового Света, оказалось для ученых неожиданностью. Эта способность у ревунов, как и у приматов Старого Света, могла возникнуть в результате реализации одного из двух альтернативных эволюционных сценариев. В первом случае ревуны могли унаследовать цветовое зрение от своего общего предшественника с обезьянами Старого Света. Во втором, который, как я уже говорил, и имел место, ревуны должны были обзавестись цветовым зрением в результате последовательности событий, не связанной с эволюцией обезьян Старого Света.
Откуда мы знаем, какой сценарий правильный?
Допустим, какие-либо два вида организмов имеют общие признаки. Чтобы установить, унаследованы ли они от общего предшественника или появились независимо, нужно посмотреть, как связаны эти виды на эволюционном древе. Отмечая наличие или отсутствие признака на эволюционном древе, можно проследить историю возникновения этого признака. Каждая точка ветвления на древе соответствует общему предку исходящих из этой точки ветвей. Если у всех видов, произошедших от данного предка, этот признак есть, то, скорее всего, предок также обладал этим признаком (рис. 6.1 а). Однако если две ветви, обладающие данным признаком, соединяются с другими ветвями, в которых этого признака нет, то, вероятнее всего, в двух ветвях этот признак появился независимым образом (рис. 6.1 б).
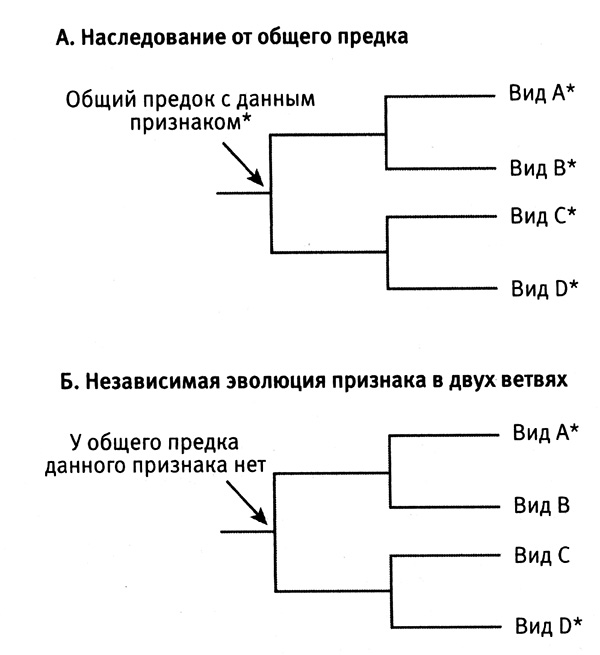
Рис. 6.1. Альтернативные варианты появления общих признаков у разных видов. Разные виды организмов могут обладать общим признаком (звездочка), если они унаследовали его от общего предка (A) или в результате независимой эволюции этого признака у данных видов после их отделения от общего предка (Б). Рисунок Лианн Олдс.
Теперь давайте взглянем на эволюционное древо приматов Старого и Нового Света (построенное на основании наследования SINE, LINE и других последовательностей ДНК), изображенное на рис. 6.2.
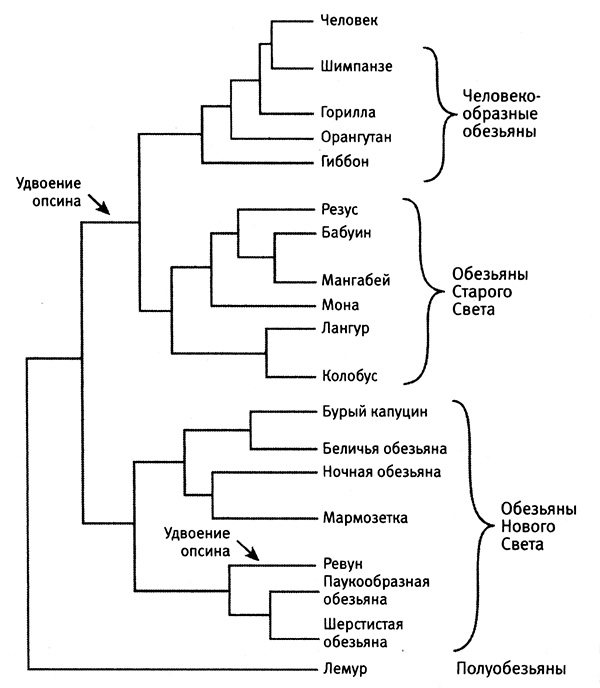
Рис. 6.2. Полноценное цветовое зрение возникало у приматов дважды. На основании распределения способности к цветовому восприятию и эволюционных связей между приматами, представленных на этой диаграмме, можно сделать вывод о том, что полноценное цветовое зрение возникало у приматов дважды (стрелки): один раз у общего предка человекообразных и других обезьян Старого Света, второй раз — у ревунов. Из статьи Giladet at., 2004, PloSBiology 2 (1): e5 (с изм.).
Из рисунка следует, что ближайшими родственниками ревунов являются другие обезьяны Нового Света, но ни один из этих видов не обладает полноценным цветовым зрением. Теоретически возможно, что у предшественника обезьян Нового Света цветовое зрение было, но все потомки, кроме ревуна, эту способность потеряли. Однако такой сценарий предполагает лишь одно приобретение и слишком много потерь. Более простое объяснение, подразумевающее гораздо меньшее количество эволюционных изменений, заключается в том, что ревун произошел от предшественника с дихроматическим зрением и самостоятельно приобрел способность различать цвета.
К счастью, чтобы докопаться до истины, мы не обязаны полагаться лишь на информацию, заключенную в эволюционном древе. Удвоение генов оставляет в ДНК свои следы. Анализ последовательности и организации современных генов опсинов в ДНК приматов Старого Света, а также ревунов и других обезьян Нового Света проливает свет на их историю. Из текста ДНК однозначно следует, что удвоение гена опсина у приматов Старого Света и у ревуна — несвязанные события. Мы можем это утверждать, поскольку в этих двух случаях размеры удвоенных фрагментов ДНК разные. У приматов Старого Света вне кодирующей последовательности каждого из двух генов имеются одинаковые последовательности длиной по 236 пар оснований. Это означает, что при создании гена средневолнового опсина была удвоена еще и прилежащая последовательность из 236 пар оснований. А у ревуна к удвоенным генам опсина примыкают гораздо более протяженные некодирующие участки. Такое могло произойти лишь в том случае, если удвоение гена опсина у ревуна произошло независимо от удвоения гена опсина у нашего с вами предка.
Кроме того, дополнительным доказательством независимости двух событий является степень сходства генов в каждой паре. Со временем после удвоения гена в каждой его копии накапливаются мутации. Чем раньше произошло удвоение, тем больше различие между двумя копиями. Так вот, у приматов Старого Света последовательности двух генов опсина различаются более чем на 5 %, тогда как гены ревуна различаются лишь на 2,7 %. Это означает, что удвоение гена опсина у ревуна произошло позднее, чем у приматов Старого Света. Это заключение вполне согласуется с палеонтологическими доказательствами более поздней эволюции обезьян Нового Света, начавшейся после разделения южноамериканского и африканского континентов.
Конвергентная эволюция опсинов ревуна и приматов Старого Света не сводится лишь к удвоению генов. Вспомним, что для полноценного цветового восприятия необходима тонкая настройка опсинов на разные длины волн и что на значение максимума поглощения опсинов MWS и LWS влияют аминокислоты, расположенные в трех ключевых позициях этих молекул. Средне- и длинноволновые опсины ревуна настроены на те же самые длины волн, а в трех ключевых позициях имеют те же самые аминокислотные остатки, что и аналогичные опсины человека и других приматов Старого Света. Это означает, что в ходе эволюции «нового» опсина MWS у ревуна и «нового» опсина MWS у приматов Старого Света произошло три одинаковые замены.
Анализ ДНК свидетельствует о том, что эволюция зрения и обоняния у ревунов шла тем же путем, что и у приматов Старого Света за миллионы лет до этого. Удвоение гена опсина, тонкая настройка ключевых позиций в молекулах опсинов и отмирание генов, ответственных за обоняние, повторились в той же последовательности и где-то с теми же деталями.
Изучение конвергентной эволюции цветового зрения у приматов и восприятия ультрафиолетового света у некоторых птиц показывает, что родственные виды могут приобретать похожие признаки, однако конвергенция ни в коем случае не ограничена степенью родства. Вспомните историю эволюции родопсина, настроенного на восприятие тусклого синего света, у глубоководного угря и у афалины (глава 4), а именно появление трех идентичных аминокислотных остатков в ключевых положениях их опсинов. Одна и та же история, но разные гены и крайне далекие друг от друга виды.
История эволюции опсинов вызывает вопрос более общего плана: насколько часто появление одинаковых признаков у разных видов организмов связано со сходными эволюционными изменениями ДНК? Давайте рассмотрим четыре примера того, как с помощью похожих механизмов с участием одного и того же гена или одних и тех же генов у разных видов и в разное время возникали одинаковые признаки.

