Книга: Далекое будущее Вселенной Эсхатология в космической перспективе
Назад: 6. Жизнь в мультивселенной
Дальше: 8. Время без конца Физика и биология в открытой вселенной[45]
Часть Третья. Биология
7. Экзотические генетические материалы и распространенность жизни во Вселенной
Э. Грэхем Кэрнс–Смит
7.1. Введение
Тот факт, что все живое на Земле обладает очень схожей биохимией, сообщает нам кое–какие сведения об истории жизни на Земле, но не о том, как в принципе должна быть устроена жизнь. Даже на Земле жизнь могла начаться с экзотических генетических материалов — я имею в виду материалы, структурно весьма отличающиеся от нашей ДНК, хотя и имеющие ту же суть и назначение, то есть способные хранить воспроизводимую информацию и влиять на окружающую среду таким образом, чтобы способствовать собственному выживанию и распространению и, следовательно, эволюционировать путем естественного отбора. По–видимому, наиболее приспособлены для поддержания такой эволюции вовсе не органические материалы, а неорганические кристаллы; и вполне возможно, что эволюция на Земле началась именно с эволюции информации, заключенной в подобных материалах (минералах) [3]. Исследование экзотических генетических материалов способно принести большую пользу при создании искусственных форм жизни [4]; кроме того, они могут представлять интерес для размышлений о распространенности жизни во вселенной — внеземной жизни, существующей сейчас или могущей существовать в далеком будущем.
Многие конкретные черты жизни на Земле кажутся универсальными, таково, например, повсеместное присутствие определенных аминокислот и нуклеотидов, универсальность белкового кода и многое другое. Однако нет основательных причин предполагать, что так называемые «молекулы жизни» являются какой-то биологической необходимостью. Существует более основательное и достаточное объяснение наблюдаемого нами единства биохимии: вся жизнь, существующая сейчас на Земле, произошла от общего предка, у которого эти черты присутствовали и были уже зафиксированы, запечатлены намертво, скорее всего, благодаря их взаимозависимости [3]. Но, если этот общий предок сам появился на свет в результате дарвиновской эволюции, значит, было время, когда эти его компоненты еще не были зафиксированы. Как такое может быть?
С помощью наивной аналогии мы можем вообразить себе ситуацию, при которой пошаговый дарвиновский процесс приводит к фиксированной взаимозависимости компонентов. Представим себе арку из камней, построенную без помощи цемента:

Рис. 7.1
Это грубая модель любой системы, состоящей из взаимозависимых компонентов. Мы можем спросить, как возможно построить такую арку шаг за шагом, то есть трогая лишь один камень в каждый момент. Вот один из возможных ответов: сначала нужно навалить груду камней —

Рис. 7.2
А затем извлекать нижние камни один за другим так, чтобы образовалась арка. На мой взгляд, жесткая взаимозависимость компонентов в современной биохимии сама по себе свидетельствует о том, что в более ранних биохимических системах компоненты были не столь жестко связаны друг с другом, что создавало своего рода «леса», позволявшие возводить шаг за шагом более сложные системы, после чего эти «леса» оказались излишними и исчезли [3].
Таким образом, мы имеем картину ранней, «текучей» стадии биохимической эволюции, за которой последовало «замораживание» жизни на земле в виде одной–единственной системы с высокой степенью взаимозависимости. Мы не знаем точно, какие материалы использовались в этой «текучей» эпохе, особенно на самых первых ее стадиях. Однако, к счастью, у нас есть возможность сделать кое–какие предположения об эволюционирующих системах вообще, а также о химических закономерностях, ограничивающих их возможности.
7.2. Потоки и гены: генетический взгляд на жизнь
Живые существа — это открытые системы, действующие внутри открытых систем, нуждающиеся в постоянном пополнении энергии и запасов материи, а также в методах избавления от продуктов разложения. Шеррингтон однажды уподобил живых существ водоворотам в потоке энергии [18]. Однако это хорошо контролируемые «водовороты», и наиболее характерный аспект такого контроля заключен в стабильных структурах, которые мы называем генами. Среда, окружающая организм, динамична, весьма динамичен может быть и он сам, но гены представляют собой некую статическую сердцевину, хранящую неизменность не только на малых и средних временных отрезках, но даже и в течение жизни многих поколений — почти невероятное постоянство, о котором столь живо писал Шредингер [15]. Чтобы подчеркнуть контраст между динамичной активностью и статичной генетической стабильностью живого существа, можно продолжить аналогию с водоворотом, уподобив живой организм системе турбулентности, возникающей вокруг камня, брошенного в поток. Форма этого камня — это унаследованная организмом генетическая информация. Форма потоков вокруг него — фенотип организма. Таким образом, фенотип можно рассматривать как часть окружающей среды, находящуюся под генетическим контролем; между фенотипом и прочей окружающей средой нет непреодолимой границы [2].

Рис. 7.3. Метафорическое изображение отношений между генетической информацией, фенотипом и окружающей средой.
Генетический взгляд на жизнь помещает в центр нашего существования генетическую память, противостоящую более динамическим, наблюдаемым аспектам нашего существования. Да, организмы — это открытые системы, способные к самоорганизации самыми различными, порой очень интересными путями. Это один из путей развития жизни. Однако открытость системы не может служить определением жизни — это слишком распространенная черта нашей вселенной. Открытые системы не просто широко распространены — в нашем мире они имеют универсальный характер. По большому счету все, с чем мы имеем дело, является открытой системой, действующей внутри открытой системы. Однако нельзя сказать, что все вокруг «живое». Отличительная характеристика живых существ — наличие у них набора средств, позволяющих успешно выживать и воспроизводить себя, долгосрочная память, переживающая процесс эволюции путем естественного отбора. В этом долгом процессе методы выживания, разумеется, меняются, но прежде всего они должны запоминаться. Вот почему им необходимо стабильное существование и возможность воспроизводить себя в копиях для следующих поколений.
Размышляя о возможной распространенности жизни во вселенной, мы должны принимать в расчет и «потоки», и «гены». Возможно, нам кажется, что единственное подходящее место для жизни во вселенной — планета типа Земли, находящаяся в непосредственной близости от своей звезды и обильно снабжаемая фотонами. Но даже для жизни на Земле важен отнюдь не только этот энергетический поток. Сама земля представляет собой ядерный реактор. Радиоактивное тепло движет земной геологией, постоянно обновляет минеральные ресурсы земной поверхности, необходимые для жизни, которую мы знаем, не говоря уж о создании активных открытых систем, например глубоководных теплых течений. Так что жизнь вполне может существовать и на планетах, отдаленных от звезд.
В дискуссии после заседания симпозиума Джон Бэрроу предположил существование еще одного источника энергии, не имеющего отношения к звездам: он напомнил о том, что, по–видимому, произошло в Окло (Республика Габон в Западной Африке), где геологические процессы в прошлом создали нечто вроде естественного ядерного реактора, расщепляющего уран-235 [13]. Это означает, что и небольшие небесные тела, гораздо меньше Земли, могут, по крайней мере иногда, предоставлять условия, приемлемые для существования жизни.
В вопросе «генов» мы также порой склонны мыслить слишком узко. Возможно, мы полагаем, что гены всегда должны представлять собой молекулы типа ДНК (или РНК). Пока никому не удалось создать иной органический полимер, способный воспроизводить более или менее значимый объем информации. Очевидно, это трудная задача. Однако органические молекулы — не единственные возможные хранилища воспроизводимой информации, которые могут существовать в природе.
7.3. Экзотические генетические материалы и генетический захват
Если мы попытаемся заглянуть в эпоху, предшествующую самому первому общему предку, и увидеть начало биоэволюционного процесса, происходившего на земле, едва ли носитель самой первой наследственной информации будет сильно напоминать ДНК. Маловероятно, чтобы молекулы ДНК или РНК возникали на земле, где еще не существовало жизни. Их нуклеотиды чересчур сложны, а сами нуклеиновые кислоты слишком хрупки и зависимы от вспомогательных систем: они могут быть только продуктами долгой эволюции путем естественного отбора. Но как это возможно, если генетический материал — необходимое предварительное условие эволюции?
Возможно, найти решение этой проблемы поможет нам аналогия с аркой. Говоря точнее, нам поможет понятие «генетического захвата». Это теоретическая схема ранней эволюции — эволюции самого центрального контролирующего механизма. В качестве простейшей его формы вообразим первый генетический материал, например минерал, способный существовать в неэволюционирующей окружающей среде. Далее он заменяется другим, более эффективным материалом (например, РНК). Этот следующий материал слишком сложен, чтобы начинать существование на примитивной земле «с чистого листа»: он становится возможен лишь в эволюционировавшей микро–среде (фенотипе), сформированной изначальными генами. Затем этот второй генетический материал постепенно вытесняет первый.
Генетический захват можно в определенной степени сравнить с тем, как в цивилизации одна технология вытесняет другую: например, для письма начинает использоваться не авторучка, а текст–процессор, лошадь заменяется двигателем внутреннего сгорания — таких примеров можно подобрать множество. Подобные замены могут быть постепенными: имеющаяся технология постепенно развивается, пока наконец не становится возможным внедрение какой-то новой, «высокотехнологичной» системы. Совершенство новой системы, делающее ненужной старую, тоже может достигаться постепенно. Однако это вовсе не значит, что новая технология всегда представляет собой развитие и усовершенствование старой. По меньшей мере столь же часто случается, что старую вещь заменяет собой совершенно новая, работающая на абсолютно других принципах. (Например, компьютерный текст–процессор возник вовсе не в результате постепенного усовершенствования авторучек.) Вполне возможно, что новая технология принесет с собой и новые материалы.
В своей простейшей форме идея генетического захвата может быть представлена в виде накладывающихся друг на друга прямых:

Первая линия — это организмы, обладающие первым (примитивным) генетическим материалом, g1, — линия, представляющая долговременную передачу от поколения к поколению информации, запечатленной в этом материале. Затем в окружающей среде, созданной эволюционировавшими фенотипами жизненной формы g1 возникает новый репродуцирующий материал — на свет появляются организмы с двумя типами генов, контролирующих различные аспекты деятельности, направленной на их выживание и распространение. Новый генетический материал не обязательно должен быть «совместим» со старым. Напротив, скорее наоборот — возможно, его преимущество состоит в том, что он несет информацию нового типа и воспроизводится новым путем. Взаимное дополнение хорошо объясняет, почему гены старого и нового типа существуют бок о бок.
Наконец, в соответствии с простейшей формой генетического захвата, «высокотехнологичные» подсистемы, основанные на материале g2, развиваются настолько, что старые, «низкотехнологичные» системы оказываются организму больше не нужны.
В дальнейшем я коротко сформулирую несколько конкретных предположений относительно g1. Но сначала мне хотелось бы развить общую идею и предположить, что на самых ранних стадиях эволюции жизни на Земле сменило друг друга множество, возможно десятки, генетических материалов, и нуклеиновые кислоты стали результатом продолжительного процесса «проб и ошибок» — эволюции генетического материала. Возможно, ближе к истине будет такая схема:
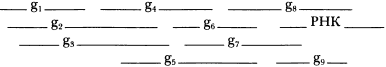
Первый шаг можно представить в виде g1 — единственной и простой воспроизводящейся характеристики, повышающей выживание и распространение вида, некоей чрезвычайно простой «функции», f1:
[g1—>f1]
Пожалуй, примером таких живых существ могут служить глины, образующие ил на дне морей. Эти глины представляют собой крошечные гибкие планки в несколько микронов шириной и несколько нанометров толщиной. Планки, в свою очередь, представляют собой пачки, состоящие, вероятно, из трех или четырех слоев, структура которых напоминает слюду. Как правило, эти планки крепятся к песчинкам (в том числе к зернам нефтеносных песчаников) на дне моря [14]. По–видимому, кристаллики ила растут благодаря расширению планок исключительно за счет атомов, прикрепляющихся к их краям. Таким образом, планки сохраняют постоянную толщину. Как правило, имеется три или четыре слюдоподобных слоя. Таким образом, толщина планки является воспроизводимой чертой, хотя она может и варьировать благодаря случайностям роста. Можно спросить: имеет ли толщина в три–четыре слоя какое-то селективное преимущество? Возможно. Ил растет благодаря «питательным» растворам, циркулирующим в порах песчаника. Слишком разросшиеся кристаллы могут загородить собой поры и снизить или остановить приток питательных веществ. Они не смогут расти и распространяться дальше. Слишком тонкие и хрупкие кристаллы легко сломать или оторвать от песчинок, к которым они прикрепляются: в этом случае они тоже закупорят поры, расположенные ниже по течению. (Именно это случается иногда, когда при добыче нефти из слоя песчаника прилагается слишком большое давление.) Возможно, толщина в три–четыре слоя идеальна для сохранения притока питательных веществ!
Разумеется, речь идет о крохотном объеме информации, да и мое объяснение в данном случае может быть неверным. Но заметим, что даже в этом случае можно говорить на биологическом языке: не только о термодинамической стабильности, доступности катионов, легкости формирования и так далее, но также о том, насколько та или иная структурная характеристика приспособлена для своего собственного выживания и распространения. Эволюция путем естественного отбора может начинаться и на самом низком уровне — были бы подходящие материалы. Их уже можно называть генетическими материалами, хотя и очень скромными.
Следующим шагом может стать сосуществование и сотрудничество различных материалов, воспроизводящих различные полезные для выживания черты («функции»):
[g1 —> f1; g2—> f2; g3 —> f3; и так далее]
Я не буду пытаться описать различные ранние функции (подробное рассмотрение этого вопроса можно найти в [3]), отмечу только, что для генов минеральных кристаллов эти функции могли включать в себя манипуляции с локально возникающими органическими молекулами. Сейчас существует значительная литература по активности глины и других подобных минералов в адсорбировании органических молекул определенным образом — расположении их между слоями минерала или по его краям, а также в катализации органических химических реакций. Есть причины считать, что эти действия функциональны в описанном выше смысле — способствуют выживанию и распространению: «гену глины» разумно окружить себя органическими молекулами, которые будут способствовать его росту или же тем или иным способом защитят его, если внешние условия (например, рН) изменятся к худшему. Известно, что органические кислоты, например лимонная кислота, активно способствуют кристаллизации глинистых минералов, перенося нерастворимые иными путями катионы, например алюминия [19].
Следующим шагом эволюции могут стать многофункциональные генетические материалы:
[…Gx—>fn, fn+1 и т. д.]
Со временем синтез органических молекул в хорошо организованном минерально–генетическом ансамбле мог принять постоянный характер, что сделало возможным воспроизведение органических полимеров. Так появился на свет новый многофункциональный генетический материал — РНК–подобный полимер, способный воспроизводить сложную и дифференцированную информацию.
Так мы переходим к истории «мира РНК» [10, 9]. Gx (РНК) начинает действовать косвенно, контролируя синтез других молекул, неспособных воспроизводиться самостоятельно, но создающих микромеханизмы (Y = белок):

Так возникает многофункциональная, косвенно действующая генетическая система (Gx= ДНК), после чего отбрасываются ненужные минеральные «леса».
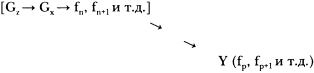
Это «жизнь, какой мы ее знаем». Структурные и каталитические функции fn, fп+1 и т. д. выполняются РНК напрямую: fp, fp+1 и бесчисленное множество других функций выполняется тысячами видов белков, каждый со своей особой структурой и уникальной последовательностью аминокислот, в свою очередь, контролируемой последовательностью ДНК, из которой он произошел. Это удивительно сложная система, но с точки зрения теории эволюции вполне объяснимая.
7.4. Кандидаты на роль кристаллических генов
В поисках возможных минеральных генетических материалов стоит вспомнить о микрокристаллических материалах с сильной внутренней связью и кристаллической структурой, обладающих некоей сложной, квазислучайной вариабельностью (отчасти сравнимой с вариабельностью последовательностей ковалентно связанных блоков в молекулах ДНК), так что каждый индивидуальный кристалл может быть уникален. Многие подобные «дефекты» воздействуют на физико–химические параметры кристаллических материалов: размер и форму кристаллов, способность поглощать мелкие молекулы, каталитические эффекты и т. п. Главный вопрос состоит в том, стабильна ли «информация» об этих дефектах и может ли она воспроизводиться в процессе роста кристаллов. Ответ на это следующий: хотя наиболее стабильные «дефекты» не воспроизводятся, существуют «дефекты», которые это делают, иногда с поразительным успехом.
Приведем пример. Структурные слои слюды и слюдоподобных глин (см. выше) образуют достаточно сложную «бутербродную» структуру с семью атомными плоскостями в каждом слое. Эти сложные слои имеют отрицательный заряд и располагаются в кристалле «пачками», один над другим, с прокладками из катионов между ними. Однако кое в чем их структура асимметрична. Так, структуры верхней и нижней плоскостей, образованных атомами кислорода, хотя и идентичны, но расположены не совсем точно друг над другом. Этот сдвиг придает слою в целом направление, которое можно изобразить стрелкой. Возникает вопрос: как расположены эти «стрелки» в слоях, лежащих друг на друге?
Часто все они направлены в одну сторону. Это можно изобразить так:
—> —> —> —> —> —> —>
Но часто случается, что их направление чередуется:
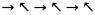
Таковы наиболее распространенные «правильные типы» слюды. Но чаще встречаются отклонения — «неправильные типы», в принципе способные нести информацию, так же как и единичная неправильность в напластовании (своего рода ДНК). Более того, известны случаи, когда какая-то «неправильность» повто–ряется через абсолютно регулярные промежутки. Например, в образцах биотитовой слюды [1] было обнаружено следующее:
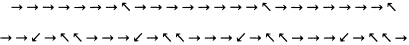
Похоже, что эти особенности возникают при росте кристаллов в результате процесса копирования.
Еще одна форма чередования встречается в глинах и других веществах, где материал представляет собой пачку химически различных слоев, чередующихся в более или менее случайной последовательности. Поразительный пример такого рода — феррит бария. В нем часто наблюдаются повторения сложных и неправильных структур, иногда простирающиеся на толщину в сто нанометров и более [12]. Изучив рост кристаллов этого материала в Глазго и Пейсли, мы предположили существование механизма копирования, обеспечивающего длинные повторы [20]. Согласно этому предположению, изначальная плата, состоящая из случайной последовательности слоев, растет путем добавления атомов по сторонам, так что изначальная последовательность сохраняется. Пачки слоев, как и «планки» морского ила, тверды и гибки. Благодаря микроморфологии начальных кристаллов, их рост происходит, по–видимому, неравномерно в разных направлениях, формируя гибкую ветвящуюся систему («водорослевый рост»). При этом разные части одной и той же пачки могут накладываться друг на друга и затем сливаться, образуя более крупные фрагменты кристалла, в пределах которых последовательность слоев, присущая начальной плате, повторяется, быть может, много раз. Сам феррит бария, возможно, растет только при высоких температурах (наши эксперименты проводились примерно при 1300 градусах по Цельсию); однако мы склонны полагать, что «водорослевый рост» представляет собой общий механизм формирования длинных повторов, присущий многим материалам, в том числе и обсуждаемым выше типам слюды.
Ил и слюда могут кристаллизоваться при обычной температуре, хотя и медленно. Быстрый синтез слюды требует, как минимум, гидротермальных условий и нескольких сот градусов по Цельсию. Это соответствует предположению, что самые первые стадии эволюции могли иметь место в океанических гидротермальных системах [6, 11].
7.5. Долгосрочное выживание и различные виды панспермии
Многие виды организмов способны выживать в сложных условиях, находясь в состоянии так называемой «отсроченной жизни»: примеры этого — семена растений или споры бактерий. Можно сказать, что перед нами потенциальные формы жизни, сохраняющие одно из двух важнейших качеств полноценной живой системы — способность сохранять информацию. Другое ключевое требование — существование открытых систем, которым эта информация передается, но они не обязаны существовать вечно — им достаточно лишь появляться время от времени.
Возможно, стоит подумать о том, как могли бы организмы приспособиться к межзвездному пространству, преодолев не только «голод» и «жажду», но и низкие температуры, космическую радиацию и огромные временные промежутки. Предположим, пара планет, таких, например, как Марс и Земля, в результате падения метеоритов обмениваются фрагментами материала. Этот обмен может создать давление отбора, благоприятное для микроорганизмов, способных попасть на метеорит и выжить в таком путешествии (ибо «Марс» периодически становится лучшим местом для жизни, чем «земля», и наоборот) — иначе говоря, отбор на способность к космическим путешествиям, включающую и возможность долгосрочной «отключки». Таким путем, возможно, разовьются споры, способные пережить миллионы лет, а затем — в редких случаях «высадки» в подходящем месте — снова «возвращаться к жизни».
Можно вообразить себе некую примитивную панспермию: споры организмов, чья генетическая память и основной механизм управления не органичны, то есть состоят не из органических молекул. Информация, передаваемая (скажем) через последовательность слоев материи, как в слюде, труднее уничтожается космической радиацией. В таком формате может существовать как примитивная, так и весьма высокоразвитая форма жизни. Она может хранить в себе информацию, позволяющую организму «вернуться к жизни», как только он встретится с благоприятными условиями, например «инструкции» по поглощению органических молекул и управлению ими, возможно, призванные сократить начальные стадии эволюции высших форм.
Затем можно задуматься о видах «развитой панспермии», основанной на генетических захватах. Первый ее вид может возникнуть благодаря естественному отбору, связанному с космическими путешествиями в форме спор, возможно, в нем появятся новые, более сложные органические полимеры.
Другую возможность «развитой панспермии» мы находим, не отрываясь от земли. Крик и Орджел, смущенные сложностью представления о нуклеиновой кислоте, возникшей на примитивной земле из «первичного бульона», предложили концепцию «направленной панспермии» [7]. Они объясняют возникновение жизни на земле очень просто: жизнь была занесена сюда пришельцами из космоса. Сами Крик и Орджел относились к этой идее не слишком серьезно; однако она не так уж безумна, если вспомнить, что совсем скоро такими «пришельцами» для других планет сможем стать мы, люди. Мы уже близки к созданию искусственных организмов — реальных организмов, а не просто компьютерных симуляций. Возможно, нам удастся создать жизнь, которая надолго переживет нас самих.
Когда мы подражаем природе, например, создаем машины, которые умеют летать или думать, естественно, что мы используем аналогичные общие принципы, но, как правило, иные материалы. Самолет, приближаясь к земле, выпускает закрылки. Также и лебедь распускает перья на крыльях, но у самолета перьев нет. Полагаю, когда мы перейдем к созданию для каких бы то ни было целей свободно живущих и развивающихся машин, то, скорее всего, материалом для них выберем отнюдь не «молекулы жизни»; в частности, чуждым для нас будет их генетический материал [4].
7.6. Заключение и итоги
Основное содержание этой главы посвящено прошлому. Я начал с рассуждения о том, что единство биохимии обусловлено неким сложным «замороженным» инцидентом в начале эволюции; все формы жизни, обитающие сейчас на Земле, обладают единой биохимией, поскольку в начале эволюции их центральные управляющие системы были «зафиксированы» в результате необратимой потери более ранних управляющих систем. Я предположил, что начальные, «низкотехнологичные» системы, не требующие совместной работы множества компонентов, сменились более эффективной «высокотехнологичной», состоящей из сложных взаимозависимых компонентов.
Такая точка зрения естественным путем приводит нас к мысли, что те же механизмы действуют и сейчас. Возможно, и сейчас в различных частях вселенной зарождается новая жизнь. Разные ее формы находятся на различных стадиях эволюции. Среди развитых форм должны встречаться «высокотехнологичные» системы, находящиеся в «замороженном» виде, что повышает их шансы на выживание в далеком будущем. Если же перейти к формам жизни, которые предстоит изобрести человечеству, очевидно, одним из желательных для них качеств является сверхдолголетие, способность существовать в условиях далекого будущего. Все эти идеи основаны на четком различении между «жизнью, какой мы ее знаем», и «жизнью вообще».
Литература
1. Baronnet, A., and Kang, Z. C., "About the Origin of Mica Poly-types", Phase Transitions, 16/17, 477–93 (1989).
2. Cairns-Smith, A. G., The Life Puzzle (Toronto University Press, Toronto, 1971), 63–64.
3. Cairns-Smith, A. G., Genetic Takeover and the Mineral Origins of Life (Cambridge University Press, Cambridge, 1982).
4. Cairns-Smith, A. G., "The Chemistry of Materials for Artificial Darwinian Systems", Int. Rev. Phys. Chem., 7, 209–50 (1988).
5. Cairns-Smith, A. G., "The Origin of Life: Clays", in Frontiers of Biology, vol. I, eds. D. Baltimore, R. Dulbecco, F. Jacob, and SR. Levi-Montalcini (Academic Press, New York, 2001), 169–92.
6. Corliss, J.В., "Hot Springs and the Origin of Life", Nature, 347, 624 (1990).
7. Crick, F., and Orgel, L. E., "Directed Panspermia", Icarus, 19, 341–46 (1973).
8. Frank, F. C., Phil Mag, 42, 1014 (1951).
9. Gesteland, R. F., Cech, T. R., and Atkins, J. F., The RNA World, 2nd ed. (Cold Spring Harbor Press, Cold Spring Harbor, N. Y., 1999).
10. Gilbert, W., "The RNA World", Nature, 319, 618 (1986).
11. Holm, N. G., ed., Marine Hydrothermal Systems and the Origin of Life (Kluwer Academic Publications, Dordrecht, 1992).
12. Kohn, J. A, Eckart, D. W., and Cook, C. F., "Crystallography of the Hexagonal Ferrites", Science, 172, 519–25 (1971).
13. Maurette, M., "The Oklo Reactor", Annual Reviews of Nuclear and Particle Science, 26, 319 (1976). Described in Barrow, J. D., Impossibility: The Limits of Science and the Science of Limits (Oxford University Press, Oxford, 1998), 187.
14. McHardy, W. J., Wilson, M. J., and Tait, J. M., "Electron Microscope and X-ray Diffraction Studies of Filamentous Illitic Clay from Sandstones of the Magnus Field", Clay Minerals, 17, 23–39 (1982).
15. Schrodinger, Е., What Is Life? (Cambridge University Press, Cambridge, 1944), chapter 4.
16. Shapiro, R., "Prebiotic Ribose Synthesis: A Critical Analysis", Orig. Life Evol Bio., 18, 71–85 (1988).
17. Shapiro, R., "The Pre-biotic Role of Adenine: A Critical Analysis", Orig. LifeEvol. Bio., 25, 83–98 (1995).
18. Sherrington, C, Man on His Nature (1937 Gifford Lecture; Cambridge University Press, Cambridge, 1940), chapter 3.
19. Siffert, В., "Clay Synthesis: The Role of Organic Complexing Agents", in Clay Minerals and the Origin of Life, eds. A. G. Cairns-Smith and H. Hartman (Cambridge University Press, Cambridge, 1986), 75–78.
20. Turner, G., Stewart, В., Baird, Т., Peacock, R. D., and Cairns-Smith, A. G. "Layer Morphology and Growth Mechanisms in Barium Ferrites" J. Cry. Or., 158, 276–83 (1996).
Назад: 6. Жизнь в мультивселенной
Дальше: 8. Время без конца Физика и биология в открытой вселенной[45]

